
针阔混交林生态系统特征及生态效益分析
2021-01-13 20:21潘婷雷云申玲芝朱晓春徐明张健
山地农业生物学报 2021年5期
潘婷 雷云 申玲芝 朱晓春 徐明 张健
摘 要:针阔混交林是一种常见、可提供多重生态服务功能的森林类型,相对于人工纯林在生态效益、防护效益以及经济效益等方面具有明显优势。本文介绍了针阔混交林生态系统的生物多样性、生产力、空间结构、土壤养分、凋落物等方面特征,对其生态效益进行分析,以期为针阔混交林生态系统的科学管理与利用提供科学依据。
关键词:针阔混交林;生态系统;生态效益
中图分类号:S718.5 文献标识码:A
文章编号:1008-0457(2021)05-0040-08 国际DOI编码:10.15958/j.cnki.sdnyswxb.2021.05.007
Analysis of Ecosystem Characteristics and Ecological Benefit of Coniferous and Broad-leaved Mixed Forest
PAN Ting,LEI Yun,SHEN Lingzhi,ZHU Xiaochun,XU Ming,ZHANG Jian*
(College of Life Sciences,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:Coniferous and broad-leaved mixed forest is a common forest type that can provide multiple ecological services.Compared with pure artificial forest,it has obvious advantages in ecological,protection and economic benefits.This paper introduced the characteristics of biodiversity,productivity,spatial structure,soil nutrients and litter of coniferous and broad-leaved mixed forest ecosystem,and analyzed its ecological benefits,in order to provide scientific basis for scientific management and utilization of coniferous and broad-leaved mixed forest ecosystem.
Keywords:coniferous and broad-leaved mixed forest;ecosystem;ecological benefits
基金項目:国家自然科学基金(31960234);贵州省科技支撑计划项目(黔科合支撑[2018]2773);大学生创新创业训练计划项目(S202010657083)
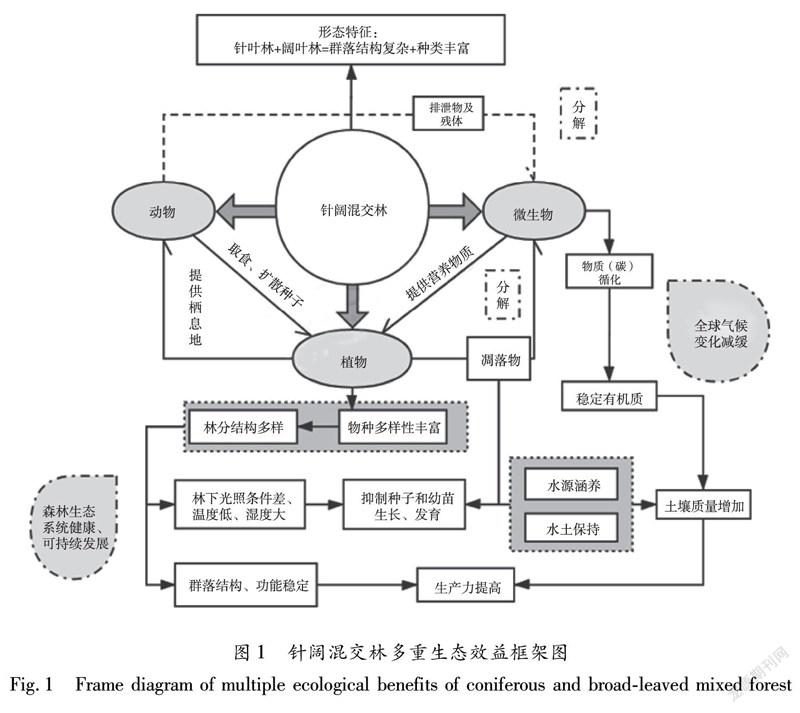
森林作为地球上最重要的陆地生态系统类型,有“地球之肺”的美誉,拥有着丰富的生物多样性和复杂的群落结构与功能,承载着陆地上70%的物种,对全球气候的调节与稳定有着重大意义[1]。森林生态系统(Forest Ecosystem)是森林中生物(以乔木为主)与其环境、生物与生物之间相互作用、相互依存的统一整体,按树种组成可分为纯林和混交林。混交林一般由两个或多个优势乔木树种或不同生活型的乔木所组成的森林。针阔混交林(Coniferous and broad-leaved mixed forests)是由针叶树种与阔叶树种为优势种共存的天然或人工混交林,是介于针叶林和阔叶林之间的过渡性植被类型。受区域气候影响,针阔混交林的种类组成在不同地区存在明显的差别。全球的温带针阔混交林主要分布于北半球北纬40°~60°之间的欧洲西缘、北美洲东缘和亚洲东缘,其物种多样性丰富,由栎树、椴树等夏绿阔叶树种和云杉、冷杉、松树等针叶树种混合组成,在群落演替过程中具有较高的研究价值。我国针阔混交林一般分为温带针叶、落叶阔叶混交林和亚热带山地针叶、常绿阔叶、落叶阔叶混交林两种类型,主要分布于东北地区(如大、小兴安岭和长白山山地)[2-3]和南方山地地区(如西南山地亚高山和中山的林区)。
我国是世界上人工林建设面积最大的国家[4],但仍以纯林为主,如马尾松纯林、樟子松纯林等,人工纯林存在病虫害多发、地力衰退严重、土壤环境恶化、易受外界环境波动的影响、森林更新困难等生态问题,而且还容易导致火灾的发生和蔓延[4-6]。天然针阔混交林在涵养水源,水土保持能力,提高生长率,提供多样化林产品,减少病虫害发生几率,防止火灾蔓延等方面发挥了重要的生态作用,在构建新型高生态、经济、社会效益的多功能森林生态体系方面也占有重要战略地位[2]。因此,有必要加强针阔混交林生态系统的基础理论与生态环境效益评价等方面的深入研究,以期为针阔混交林的科学管理与利用提供参考。
1 针阔混交林生态系统主要特征
针阔混交林植物区系的地理成分较为复杂,植物生活型谱以高位芽植物为主,其次为地面芽植物,含有一定比例的地下芽和地上芽植物,一年生植物种类极少[7]。实践证明,混交林能够有效地增加林地生物物种的多样性,提高林地养分水平和生产力,从根本上改善林分的生态环境[8]。
1.1 林分结构、多样性与生产力相互关系
物种多样性和生产力是森林生态系统的重要特征,而林分结构可以强烈地影响森林的生长和其他生态过程,如水的平衡、碳分配、营养物质的循环和光的动力学[9]。不同于多样性的间接作用,林分结构能直接影响森林生产力。植物种间互作将有利于增加在自然干扰、胁迫或捕食等条件下难于生存的植物种类,从而促进森林群落物种多样性增加[10]。“生态位互补假说”认为不同物种由于生态位差异会对资源进行更充分的利用,从而可提高森林生产力[11]。针阔混交林中不同林层林分结构对生产力的作用方向不同,林下层的物种多样性较林冠层的物种多样性对生产力影响弱,这主要是由于树木生长发育和环境异质性的综合作用[12]。
植物个体的生长、死亡和更新等动态将直接影响群落空间结构,良好的林分空间结构是判断群落能否维持稳定发展的重要前提。混交度是描述混交林中不同树种之间隔离程度的重要空间结构指数[13]。树种混交度是群落发育过程中种群与环境、种群与种群之间长期相互作用的结果。混交林中树种之间的隔离程度由树种类型及其不同树种数量和空间分布共同决定。一般群落的树种类型越多,其混交度也越大。群落混交程度越高反映出其群落结构越稳定。周红敏等[14]在凤阳山自然保护区针阔混交林空间结构研究中指出林木混交度较强,物种多样性高,林分空间结构相对稳定。朱军等[15]发现紫金山针阔混交林的混交度对林下植物多样性具有重要影响。混交林相对纯林的空间结构多样性较复杂;混交林的物种多样性、群落更新潜力和群落稳定性均高于纯林。混交林中的树种混交比例會改变森林内物种丰富度和多样性、种群更新能力、群落生物量等,从而影响森林群落的稳定性[16]。
森林空间结构是森林生长过程的重要驱动因子,对森林的功能表达和发展趋势具有关键性作用。物种的种类与空间分布格局决定群落空间结构,物种间不同空间关系塑造了群落结构的特异性,良好的空间结构是群落能够维持稳定发展的前提条件。在一个群落中,优势种作为群落演替的内在驱动力,最能体现一个群落内部的空间结构和格局的形成过程。林分空间结构决定林木间的竞争优势及其空间生态位,在很大程度上决定林分的稳定性、发展的可能性。有学者提出从林木空间分布格局、树种混交度和树木竞争等方面反映林分内部的结构变化[17]。林木空间分布格局包括聚集分布、随机分布和均匀分布等形式。李建等[18]研究发现长白山次生针阔混交林群落的水平格局为随机分布,大小分化程度为中庸分布且树种混交度较强时,群落具有良好且较稳定的空间结构。
群落结构受生境异质性、植物繁殖体传播范围和种间相互作用等因素影响且随其研究尺度的变化而变化。不同的研究尺度可能会得出不同的分布类型。在较小的尺度范围内可能受到种内或种间竞争、种子扩散限制等因素的影响,而在大尺度范围内可能受自身的生物学特性、种群分布区的异质性和外界无机环境等因素的影响[19]。近年来,许多学者对我国长白山地区针阔混交林空间格局进行了大量研究,为我国针阔混交林空间格局的进一步研究奠定了基础[20-24]。
1.2 土壤性质
林地土壤肥力主要靠林木养分的归还量和养分循环速度来维持和提高,并可直接影响植物生长,在植物体构成和生理代谢方面发挥着重要作用。混交林中植被的种类多数量大,对土壤涵养水源和累积有机质有巨大作用,明显改善了土壤的通透性,降低了土壤的紧实程度和容重,特别是表层土壤最为明显[25]。混交林中土壤最上层有大量的凋落物,它们分解后成为表层土,大大改善了表层土的物理性状,增加了土壤有机质和团粒结构,提高了土壤孔隙度,减小了土壤容重。在混交林中,由于增加了阔叶树种,加速了枯枝落叶的分解速率,各营养元素的释放量得到提高[26],故与纯林相比,所有混交林中同一土层中的有机质、全氮、水解氮、全磷、速效磷、全钾、速效钾的含量都有不同层度的增加。有更厚的枯枝落叶层和腐殖质层,较纯林地其土壤质地松散、结构优异、吸持水的能力强、透水性大[27]。土壤的物理性状尤其是土壤类型、孔隙度和结构等与持水能力、保水能力及渗水能力等方面直接相关,影响着林地植物对土壤养分的吸收和保持。针阔混交林的土壤理化性质随演替过程发生变化,毛管孔隙度、速效钾和全磷含量是影响不同演替阶段土壤质量的主要因子;经过不同演替阶段,表层土壤理化性质质量的综合得分表现为中龄林 < 成熟林 < 近熟林 < 老龄林;随着演替进行,针阔混交林的表层土壤理化性质基本呈现质量提高趋势,到老龄林时期达到最佳[28]。
1.3 凋落物
森林的凋落物在森林资源保护、可持续利用方面发挥着重要作用,且在维持土壤肥力,保证植物再生长养分的可循环利用性,促进森林生态系统正常的物质循环和养分平衡方面起着重要的作用[29]。凋落物是林木养分归还的主要途径,凋落物积累量和分解速率决定着林木养分归还量的大小[30];一般而言,凋落物分解速率与N、P含量成正比。凋落物量是一个动态值,它受气候、地形、海拔、土壤和植被类型等多因素影响。从热带到亚热带和温带,凋落物现存量随着纬度的增高而增加的结论基本一致。凋落物层具有调节和阻滞地表径流的生态水文功能,如缓冲雨水的动能,避免土壤结构受雨水溅击而破坏[31]。针对我国东北地区地带性顶级植物群落——针阔混交林的研究表明,其凋落物量平均为4.49 t/hm2,其中针、阔叶的凋落物量分别占全年的23%和60%左右[32]。凋落物的水源涵养功能由凋落物量和持水能力共同决定;一般情况下,凋落物的现存量越多,持水能力越强,其水源涵养功能越好[33]。针阔混交林的凋落量较大,且在不同森林类型中营养元素的储量一般表现为:未分解层 <半分解层 < 完全分解层[34]。总之,凋落物的分解对森林生态系统的结构和功能有着不可忽视的影响。
1.4 针阔混交林相关的生态机制
1.4.1 物种更新与维持机制
幼苗和幼树是植物群落更新的关键,是群落维持稳定的决定因素,且直接影响种群繁衍[35]。幼苗作为林木更新过程中最敏感脆弱的环节,其存活状况直接影响森林群落的物种组成和生态系统的稳定性[36]。针阔混交林由于林下光照条件差、灌木、草本层丰富,对幼苗和幼树形成了竞争抑制作用。再加上混交林下腐殖质及枯枝落叶层较厚,大大降低了种子接触土壤萌发的几率。已有研究表明,林下幼苗种类及物种丰富度随着采伐强度的增加而增加。首先,适度采伐可改善林下微环境(如光照和土壤温度)、优化林分结构、调整林分密度,使林下光照增多,利于耐阴树种的生长,影响着林下植被多样性,进而影响林下植被的更新和生长。其次,采伐也可能使森林树种的产子量增加,充足的光照和适宜的土壤温度促进种子萌发,提高物种多样性[37-38]。
物种多样性维持及其空间尺度效应是生态学研究的重点问题[39],不同空间尺度下生态学过程可能存在差异[40]。在不同尺度上植物群落物种多样性的发生和维持主要通过物种生物学和生态学特性的差异和生境异质性得以实现[41]。基于单物种—面积模型,可将物种对邻体多样性格局的影响区分为促进种、中性种和抑制种。生物多样性促进种在群落多样性格局组建过程中起主导作用,中性种所代表的随机过程在群落组建过程中所发挥的作用也不容忽视。同时,在中度和高度胁迫的环境中,植物间正向互作通过扩展现实生态位,可以提高群落生物多样性[42]。
1.4.2 竞争效应
竞争指2个或多个植物体在同一环境资源和能量的争夺中所发生的相互作用;研究树种的种内种间竞争特性对改善森林结构、提高森林景观品质具有重要作用;森林物种竞争关系研究是森林生态学的重要研究领域之一。辛营营等[43]通过对杭州青山湖地区针阔混交林优势种的数量研究发现,优势树种的种内竞争大于种间竞争,说明优势树种呈现出一定的聚集分布状态,竞争强度与胸径大小之间存在着幂函数关系,但当胸径达到一定程度即树种相对成熟稳定时两者之间的关系不再明显,认为这为针阔混交林的可持续经营实践中“间密留稀,去弱存强”减低林木间竞争的抚育措施提供了理论支持。杜秀芳等[44]对杭州市临安区多种森林类型的竞争关系研究表明,该区域森林的竞争主要来源于种内竞争,种内平均竞争顺序为:人工针叶林>天然阔叶林>天然针叶林>天然针阔混交林>人工针阔混交林>人工阔叶林;天然针阔混交林种间平均竞争最大,人工针阔混交林次之,针阔混交林的树种丰富度明显高于其它。
1.4.3 边缘效应
边缘效应指在两个或多个不同性质的生态系统(或其他系统)交互作用处,由于某些生态因子(可能是物质、能量、信息、时机或地域)或系统属性的差异和协合作用而引起系统某些组分及行为(如种群密度、生产力和多样性等)的较大变化。边缘效应的存在使林内到林外的生態环境产生梯度变化,进而影响动植物分布、群落结构和功能、群落能量流和物质流等[45]。奚为民等[46]通过对缙云山森林林窗及其边缘效应的研究表明,林窗边缘区由于边缘效应的作用而有增大物种多样性的趋势,其中针阔混交林中前期林窗的边缘效应更为显著;而针阔混交林林窗具有一般意义上的比常绿阔叶林林窗相对较强的边缘效应强度。林窗边缘效应规律可以为混交林的种群结构重建以及选择最佳的林木择伐面积和强度等提供科学依据。因此,维持一定的林窗边缘效应强度和边缘区面积,有利于森林物种多样性的保持和林木更新。
2 针阔混交林生态效益分析
森林是陆地生态系统的主体,是人类赖以生存的自然环境,具有生态效益、经济效益和社会效益三大效益。针阔混交林生态系统的多种特征决定了其独特的生态效益优势,其物种多样性优于针叶纯林,进而影响着森林生态系统的演替和动态,改变着森林群落的区系组成、结构和遗传多样性[47]。
2.1 针阔混交林的生态环境
针阔混交林物种多样性丰富、林分结构复杂、综合生产力高、生态系统相对稳定,生态环境良好。通过人工经营的方法,根据适地适树原则,有目的地将针、阔树种按一定比例混交种植,在其生长过程中,通过环境选择和竞争作用能自然形成合理协调的林分组合;同时,与针叶纯林相比,在针阔混交林内草本、灌木层植物更为丰富,森林的水平垂直层次结构十分合理[48];即使在混交林被砍伐后,其生态环境破坏较小,易于森林群落的更新。
2.2 针阔混交林的水循环效益
2.2.1 针阔混交林的树冠截留降雨率
不同林分因树冠结构特征的不同,其降雨截留作用会存在明显差异。有研究表明,人工针阔混交林对树冠截留的功能要强于松叶纯林[49]。针阔混交林的多层林冠一般会呈现出垂直互补协调的特点,能够阻拦、延长和增加雨水流经树冠的时间。因此,如果是在降雨强度相同且年龄条件相同的基础上,针阔混交林的林冠截留降雨率要比针叶纯林多[48]。林冠对降雨的截留可减少雨水对地面的冲蚀、也影响着土壤中水分的分布及蒸发、蒸腾等过程。林冠对降雨的截留作用揭示了森林与水的关系问题,具有重要的生态水文意义[50]。
2.2.2 针阔混交林的涵养水源和水土保持能力
针阔混交林的枯枝落叶量多,能相互促进分解,吸持雨水能力强,在一定程度上能缓冲雨水对地表的冲击;再加上植物细根能固定土壤,水土保持能力强。不同森林类型单位面积水源涵养能力一般表现出:常绿阔叶林>针阔混交林>常绿针叶林>落叶阔叶林>灌木林[51]。与针叶纯林相比,针阔混交林的枯枝落叶层和腐殖质层相对较厚,具体为:针阔混交林的枯落物总厚度比针叶纯林多58%,腐烂物的厚度多121%,持水率多37%[48]。此外,混交林具有数量大、密度高等特点,相对减少了混交林内的光照面积和时间。再加上林内通风效果不佳,水分的蒸发就随之得到抑制,提高了林内的湿度,改善了混交林的气候环境[52]。
2.3 针阔混交林的土壤理化性质改善作用
混交林较纯林相比,具有多层的树冠以及茂密的树叶,其林下物种多样性较高,凋落物丰富,易于形成松软易分解的腐殖质,其养分分解能力及转化速率加快,土壤有机质质量分数高。混交林植物可以有效改善土壤的通气状况,使得林地土壤变得疏松多孔,土壤孔隙度增大[53],提高了混交林地的土壤肥力,改善了土壤结构,增强林地的容蓄能力;混交林植物的根系生长迅速,根的吸收功能增强,有效解决纯林地力衰退问题,更有利于混交林植物的生长。有学者通过对针阔混交林和人工纯松林的土壤性质分析比较发现,人工针阔混交林不仅能提高土壤肥力,较之纯林还能吸收更多的二氧化碳,释放更多的氧气,从而有效地增加森林的碳汇能力[54]。
3 针阔混交林研究与应用的展望
针阔混交林是世界主要的森林类型之一,具有独特的森林生态系统结构与功能,发挥着重要的生态效益、防护效益和经济效益。随着全球气候变化和生态环境退化的挑战,针阔混交林已成为林学、生态学等领域研究热点之一。国内对针阔混交林的研究主要集中在其分布较为密集的大小兴安岭、长白山等山地,对西南地区也有探究[18,22-24,35,47]。针对针阔混交林的研究与应用是多方面的,包括生物多样性、生产力、动态变化、空间结构、土壤养分、凋落物、林下种植、近自然经营[55-56]等方面。下面将试图从造林学、生态学和学科交叉等角度出发,对针阔混交林研究与应用进行展望。
3.1 生态学角度:针阔混交林的生态学机制研究及其生态功能评价 针阔混交林中物种丰富,有复杂的结构,且林分的郁闭度大,因此造成叶面积指数大,景观层次及冠层结构相对较好。针阔混交林的枯落物层和土壤层的综合水源能力优于针叶混交林和杉木林,能改善森林的生态环境,对林分的增产有促进作用,应该加强对针阔混交林的研究保护,提高森林生态系统在涵养水源、保持水土方面的生态功能,充分展现混交林功能效益和潜在的性能[57]。
3.2 造林学角度:构建多重生态功能的针阔混交林 基于地理环境特征、种间关系、功能性状、空间格局等多角度的综合分析针阔混交林群落生态学特征与规律,探究混交林营建的最佳的针、阔混交比例,造林不同阶段的生态调控技术,激发混交林的独特优势,构建多重生态功能的针阔混交林,创建优良的生态环境,更好满足人类日益增长森林生态产品的多样化需求。
3.3 交叉学科角度:针阔混交林的森林康养与林下经济研究与应用 森林康养涉及了医疗、旅游、生态、社会和经济等领域,是林业发展催生的新模式[58],它有助于捕获碳排放,促进生物多样性,并在农村和城市周边景观中提供娱乐和美学价值[59]。较高的树种多样性增加了生态位的数量,从而进一步增加了伴生物种的数量,例如,能为林下植物和动物提供更好的栖息地[60],营造生态、景观、经济多效益多功能混交林群落[61]。林下经济是最近兴起的农林复合型产业,可实现农林牧业资源的合理共享和循环协调发展,具有良好的经济效益和社会效益[62]。目前,林下经济发展迅速,林下植物和动物都有具有较大发展潜力。同时,还可发展林下文化产业,发展旅游胜地、养生保健、有机食物等。例如:北京市建设林下经济示范园区发展亲子采摘、森林观光、幼苗种植等。东北地区發展的林间养鹿、种蛙、种参等。云南发展石斛、菌菇和蔬菜等林下产品,取得了显著的经济发展和环境保护效果[63]。由于针阔混交林具有物种多样性丰富、林分结构多样、形态多样、腐殖质层厚和生态环境良好等特点,为林下创造了更多的异质光环境,促进了林下动植物生长,对于森林康养和林下经济的发展提供了优质条件。
针阔混交林对快速恢复森林的生态环境性能,涵养水源,水土保持能力,生长率的提高,产品的多样化,防护效益的发挥,减少病虫害发生几率,防止火灾蔓延等方面发挥了巨大作用;同时还能为市场提供多元化、多性能的木制产品,拉动经济发展。因此,对于针阔混交林进行多方面多层次的研究,对保障森林生态系统持续性和稳定性具有实践意义,为林分结构化经营、混交林的抚育、林分质量评价和树种空间多样性评价等提供了科学依据,同时也为植被恢复与保护、资源利用及生物多样保护和森林管理提供参考。针阔混交林对水土保持、涵养水源、调节气候、农业生产和经济的可持续发展有重要作用,对土壤生产力匮乏地区尤其是喀斯特地区具有良好的生态影响,有较广阔的发展前景。
参 考 文 献:
[1] 苏巧灵,李安民,袁士云,等.甘肃小陇山暖温带针阔混交林物种组成和群落结构[J].应用生态学报,2020,31(10):3305-3312.
[2] 周秋静,韩文斌,赵常明,等.神农架天然针阔混交林的物种组成和群落结构[J].生态学杂志,2019,38(1):11-8.
[3] 张炜琪,陈辉,林文俊,等.武夷山针阔混交林的群落多样性[J].森林与环境学报,2016,369(2):136-140.
[4] DEL RIO M,PRETZSCH H,ALBERDI I,et al.Characterization of the structure,dynamics,and productivity of mixed-species stands:review and perspectives[J].European Journal of Forest Research,2016,135:23-49.
[5] NAKAJIMA T,TATSUHARA S.Silviculture guidelines for developing mixed forests in old coniferous plantations under long-rotation stand density control[J].Forest Science and Technology,2012,8:127-134.
[6] 王瑞辉.我国人工造林种存在的生态问题及对策[J].湖南林业科技,1999,26(4):30.
[7] 陈霞,袁在翔,金雪梅,等.紫金山针阔混交林群落特征[J].中国城市林业,2019,17(5):12-17.
[8] 田大伦,项文化,闫文德.马尾松与湿地松人工林生物量动态及养分循环特征[J].生态学报,2004,24(10):2207-2210.
[9] FORRESTER D I.Linking forest growth with stand structure:Tree size inequality,tree growth or resource partitioning and the asymmetry of competition[J].Forest Ecology and Management,2019,447:139-157.
[10] HACKER S D,GAINES S D.Some implications of direct positive interactions for community species diversity[J].Ecology,1997,78(7):1990-2003.
[11] 譚凌照,范春雨,范秀华.吉林蛟河阔叶红松林木本植物物种多样性及群落结构与生产力的关系[J].植物生态学报,2017,41(11):1149-1156.
[12] 鲁君悦,吴兆飞,张春雨,等.吉林蛟河针阔混交林林层结构对生产力影响研究[J].生态学报,2021,41(5):2024-2032.
[13] 张结存,徐丽华,张茂霞,等.基于物种空间结构和多样性的改进型混角度研究[J].浙江农林大学学报,2014,31(3):336-342.
[14] 周红敏,彭辉,蒋敏,等.凤阳山自然保护区针阔混交林空间结构研究[J].福建林业科技,2019,46(3):51-54,67.
[15] 朱军,湛斌,陈霞,等.紫金山针阔混交林空间结构特征与植物多样性分析[J].中南林业科技大学学报,2018,38(1):75-80,93.
[16] 康希睿,李晓刚,张涵丹,等.不同混交措施下杉木人工林群落稳定性特征[J].生态学杂志,2020,39(9):2912-2920.
[17] 邓红兵,陈喆菲,许东,等.森林多目标经营单元的时空尺度扩展[J].生态学报,2015,35(1):65-70.
[18] 李建,李晓宇,曹静,等.长白山次生针阔混交林群落结构特征及群落动态[J].生态学报,2020,40(4):1195-1206.
[19] 王燕,毕润成,许强.山西太岳山南部针阔混交林群落特征及空间分布格局[J].广西植物,2017,37(7):901-911.
[20] 高杰,张君,程艳霞,等.长白山不同林型空间格局及树种多样性多尺度分析[J].北京林业大学学报,2014,36(6):103-104.
[21] 杜志,亢新刚,包昱君,等.长白山云冷杉林不同演替阶段的树种空间分布格局及其关联性[J].北京林业大学学报,2012,34(2):14-19.
[22] 石蒙蒙,杨华,王全军,等.长白山云冷杉针阔混交林幼苗幼树空间分布与关联性[J].北京林业大学学报,2020,42(4):1-11.
[23] 李艳丽,杨华,亢新刚,等.长白山云冷杉针阔混交林天然更新空间分布格局及其异质性[J].应用生态学报,2014,25(2):311-317.
[24] 常新华,赵秀海,曾凡勇,等.长白山针阔混交林主要树种空间分布及其环境解释[J].北京林业大学学报,2009,31(1):7-12.
[25] 邓恢.马尾松阔叶树混交林土壤理化性质比较研究[J].福建林业科技,2012,39(1):41-44,52.
[26] PANG Y,TIAN J,ZHAO X,et al.The linkages of plant,litter and soil C:N:P stoichiometry and nutrient stock in different secondary mixed forest types in the Qinling Mountains,China[J].Peerj,2020,8:e9274.
[27] 于飞.临朐县石灰岩山区针阔混交林土壤理化性状及水文效应研究[D].泰安:山东农业大学,2014.
[28] 林文树,穆丹,王丽平,等.针阔混交林不同演替阶段表层土壤理化性质与优势林木生长的相关性[J].林业科学,2016,52(5):17-25.
[29] 秦倩倩,王海燕,李翔,等.长白山云冷杉针阔混交林半分解层凋落物生态功能[J].林业科学研究,2019,32(1):147-152.
[30] BRADFORD M A,BERG B,MAYNARD D S,et al.Understanding the dominant controls on litter decomposition[J].Journal of Ecology,2016,104:229-238.
[31] 朱金兆,刘建军,朱清科,等森林凋落物层水文生态功能研究[J].北京林业大学学报,2002,24(5):30-34.
[32] 蒋云峰.长白山针阔混交林主要凋落物分解及土壤动物的作用[D].长春:东北师范大学,2013.
[33] 刘世荣.中国森林生态系统水文生态功能规律[M].北京:中国林业出版社,1996.
[34] 常雅军,曹靖,李建建,等.秦岭西部山地针叶林凋落物层的化学性质[J].生态学杂志,2009,28(7):1308-1315.
[35] 李小双,赵安娜,党承林,等.昆明西山云南油杉针阔混交林的群落结构及其更新特征研究[J].云南大学学报(自然科学版),2013,35(4):549-557.
[36] YAN Y,ZHANG C,WANG Y,et al.Drivers of seeding survival in a temperate forest and their relative importance at three stages of succession[J].Ecology & Evolution,2015,5(19):4287-4299.
[37] SEIWA K,ETOH Y,HISITA M,et al.Roles of thinning intensity in hardwood recruitmentand diversity in a conifer,Criptomeria japonica plantation:a 5-year demographicstudy[J].Forest Ecology & Management,2012,269(2):177-187.
[38] 米爽,宋子龙,秦江环,等.抚育采伐对吉林蛟河针阔混交林幼苗更新的影响[J].北京林业大学学报,2019,41(5):159-169.
[39] GILADI I,ZIV Y,MAY F,et al.Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem[J].Journal of Vegetation Science,2011,6:983-996.
[40] HE F L,LAFRANKIE J V,SONG B.Scale dependence of tree abundance and richness in a tropical rain forest,Malaysia[J].Landscape Ecology,2002,17:559-568.
[41] 汪殿蓓,暨淑仪,陈飞鹏,等植物群落物种多样性研究综述[J].生态学杂志,2001,20(4):55-60.
[42] 马志远,石玲,吴相菊,等.长白山次生针阔混交林乔木多样性维持机制研究[J].北京林业大学学报,2014,36(6):93-98.
[43] 辛营营,韦新良.青山湖针阔混交林优势树种竞争的数量研究[J].浙江农林大学学报,2011,28(4):601-606.
[44] 杜秀芳,汤孟平,潘建勇,等.临安区不同森林类型竞争指数比较研究[J].生态学报,2020,40(12):4064-4072.
[45] 田超,杨新兵,刘阳.边缘效应及其对森林生态系统影响的研究进展[J].应用生态学报,2011,22(8):2184-2192.
[46] 奚为民,钟章成,毕润成.四川缙云山森林群落林窗边缘效应的研究[J].植物生态学与地植物学学报,1993(3):42-52.
[47] 刘彦春,张远东,刘世荣,等.川西亚高山针阔混交林乔木层生物量、生产力随海拔梯度的变化[J].生态学报,2010,30(21):5810-5820.
[48] 丘菊秀.针阔混交林的优势及应用分析[J].绿色科技,2020(7):119-121.
[49] 宋德利.人工针阔混交林水源涵养能力的试验[J].防护林科技,2014(2):36-37,104.
[50] 张光灿,刘霞.林冠截留降雨模型研究进展及其述评[J].南京林业大学学报,2000,24(1):64-68.
[51] 郝新朝,安裕伦,伍显,等.贵州省森林生态系统水源涵养功能价值评估[J].安徽农学通报,2019,25(Z1):94-97,130.
[52] 刘涌.混交林的优点及混交原则相关问题分析[J].科技创新与应用,2014,(12).
[53] 秦娟,唐红心,杨雪梅.马尾松不同林型对土壤理化性质的影响[J].生态环境学报,2013,2(4):598-604.
[54] 陈杰.白桦+红松人工混交林生态效益的研究[J].防护林科技,2019(9):27-28,36.
[55] 张发根,张文斌,赖根伟,等.混交林下多花黄精仿野生种植技术探讨[J].华东森林经理,2019,33(4):22-24.
[56] 臧丽鹏,张广奇,何跃军,等.近自然经营对黔中地区马尾松灌草群落多样性及物种共存的影响[J].山地农业生物学报.2021,40(3):21-28.
[57] 钟梁,高友英,孙浩,等.扶河上游生态公益林4种森林类型枯落物层和土壤层水文效应[J].南方林业科学,2017,45(6):5-8.
[58] LIU C L C,KUCHMA O,KRUTOVSKY K.Mixed-species versus monocultures in plantation forestry:Development,benefits,ecosystem services and perspectives for the future[J].Global Ecology and Conservation,2018,15:e00419.
[59] 刘琼.森林康养产业发展的必然性可行性和路径选择[J].经济师,2021(2):36-37,39.
[60] BRAVO-OVIEDO N,PRETZSCH H,AMMER C,et al.European mixed forests:definition and research perspectives[J].Forest Systems,2014,23(3):518-533.
[61] 于瀚,欧静,漆倩,张智叶.黔中地区马尾松-杜鹃植物群落多样性研究[J].山地农业生物学报,2020,39(2):14-20.
[62] 陈福军.发展林下经济推进林下资源可持续发展利用的探索[J].农业开发与装备,2020(7):39.
[63] 黃美水.谈林下经济产业现状及发展重点[J].百科论坛电子杂志,2019(16):227.
通讯作者:张健(1980—)男,博士,副教授,主要从事微生物生态学方面研究,E-mail:zhangjian12102@163.com.
猜你喜欢
林业与生态(2021年4期)2021-05-28
当代经济管理(2016年11期)2016-12-14
商情(2016年40期)2016-11-28
科教导刊(2016年27期)2016-11-15
科技视界(2016年18期)2016-11-03
企业导报(2016年9期)2016-05-26
林业与生态(2014年5期)2014-07-07
中国经济信息(2004年16期)2004-08-28
