
喀斯特湿地水葫芦(Eichhornia crassipes)生长及表型可塑性特征对不同富营养化水体的响应*
2021-01-12 06:58滕秋梅孙英杰沈育伊张德楠徐广平周龙武张中峰黄科朝何成新
湖泊科学 2021年1期
滕秋梅,孙英杰,沈育伊,张德楠,徐广平,3 ,周龙武,张中峰,黄科朝,何 文,何成新
(1:广西壮族自治区中国科学院广西植物研究所,广西喀斯特植物保育与恢复生态学重点实验室,桂林 541006) (2:广西壮族自治区中国科学院广西植物研究所,广西植物功能物质研究与利用重点实验室,桂林 541006) (3:中国地质科学院岩溶地质研究所,自然资源部/广西岩溶动力学重点实验室,桂林 541004)
水体富营养化是我国近40年来众多湖泊和河流水域所面临的主要问题和挑战之一[1],在人类活动的影响下,这种演化过程大大加快,富营养化引起的环境问题日益严重[2-4]. 研究表明,城市周边的湿地是整个景观物质流的汇聚地,积累了较多的沉积物,伴随湿地常常出现水体富营养化[5],进而给外来入侵植物创造了一定的空生态位[6].
水葫芦,学名凤眼莲(Eichhorniacrassipes),原产于南美洲,属雨久花科,它还有其他俗名,如凤眼蓝、水凤仙、水风信子等[7]. 作为主要的外来入侵物种之一,多个国家和地区受到水葫芦入侵的危害[8],对区域生物多样性和生态系统构成了巨大的威胁[9]. 桂林会仙喀斯特湿地是中国最大的岩溶湿地,在调节地区气候、涵养水源和生物多样性保护等方面发挥着重要作用,也是响应全球变化和人类活动较为敏感的生态系统[10]. 近年来随着湿地旅游业的开发利用,会仙喀斯特湿地过多地受到开垦、养殖、污水排放等人类活动的影响,出现水体富营养化问题[11-12]. 水葫芦入侵面积在急剧扩张,主要集中在富营养化水域中[11],并形成高密度单优势水葫芦群落,是会仙湿地危害严重的入侵植物.
植物表型可塑性(plant phenotypic plasticity)通常指同一基因型植物适应不同生境而改变形态、生理和行为的特性[13]和不同基因型在环境作用下能表达的表型范围[14],是植物适应环境的重要方式,对其分布、拓殖、存活能力都具有重要的影响[15],是植物适应异质环境最直观的策略之一. 研究植物表型可塑性具有重要的生态学意义[16-17]. 水生植物可以通过形态结构调节、分株间物质传输与共享、相连分株形态和生理功能特化等策略来适应异质的生境[18-20],这有利于其拓宽生态位,占据更多样的生境[21]. 陆生植物的表型可塑性受到植物生态学界的关注较多,但水生植物表型可塑性的研究仍显薄弱[20].
研究表明,一般在不同生境下物种的表型可塑性可能会发生适应性分化[22],表型可塑性对外来物种的入侵也起到了关键作用[23]. 水生植物在喀斯特湿地生态系统物质循环中起着关键作用,水体富营养化引起了湿地水生植物多样性的降低,水葫芦入侵对喀斯特湿地生态系统可能产生了重要影响. 因此,研究喀斯特湿地入侵植物水葫芦的表型可塑性特性,对于深入探讨喀斯特湿地生态系统保护和植被恢复具有重要的意义. 目前,对于水葫芦的研究主要集中在对鱼类、水质和生物群落等的负面影响[24],蓝藻水华聚集对水葫芦生理生态的影响[25],对水体营养盐的去除及生理响应[26],对富营养化水体中氨氧化和反硝化微生物的影响[27],对重金属(镉、锌、铅、汞)的吸附[28]等方面. 在桂林会仙喀斯特湿地,不同富营养化水体对水葫芦表型可塑性的影响尚不清楚. 因此,我们试图回答以下科学问题:1)在桂林会仙喀斯特湿地,水葫芦生长及表型可塑性特征对不同富营养化水体有怎样的响应趋势?2)表型可塑性是否促进了水葫芦在富营养化水体中的生长及生态适应性?在不同富营养化水体条件下,探讨水葫芦生长及表型可塑性的响应特征,有助于认识水葫芦的生态适应性,可提高对水葫芦的预警和综合防治,以期为桂林会仙喀斯特湿地环境保护和生态修复提供理论依据.
1 材料与方法
1.1 研究区概况
会仙岩溶湿地(25°01′30″~25°11′15″N,110°08′15″~110°18′00″E,图1)位于桂林市临桂区会仙镇,东至雁山区,西至四塘乡,海拔150~160 m,总面积约 120 km2,是以草本沼泽和湖泊为主的喀斯特湿地,是国内为数不多的中低海拔大型喀斯特湿地之一 ,现已规划为我国最大的喀斯特湿地公园. 该区属亚热带季风气候区,年均气温16.5~20.5℃,极高气温达38.80℃,极低气温为-3.30℃,年均降雨量为1890.4 mm. 降雨时空分布不均,多集中在每年的3-8月,形成了春夏季雨多而集中、秋冬季少雨干旱的特点. 土壤以红黄壤和红壤为主,集中分布于洼地、平原和缓坡,土壤层薄甚至基岩裸露. 湿地植被以挺水植被和沉水植被为主,植物种类较多,且生长茂盛,盖度常可达80%~95%,主要建群种有芦苇(Phragmitescommunis)、华克拉莎(Cladiumchinense)、五刺金鱼藻(Ceratophyllumoryzetorum)、石龙尾(Limnophilasessiliflora)等[29].
图1 桂林会仙喀斯特湿地区位Fig.1 Location of the Huixian karst wetland in Guilin
1.2 实验方法
前期调查中发现,会仙喀斯特湿地入侵植物水葫芦主要分布在水体富营养化区域[11],每年3-4月为水葫芦的萌芽阶段,6月为水葫芦群落的旺盛生长期,8月为水葫芦暴发的高峰期,12 月下旬水葫芦开始枯萎和逐渐死亡. 在前期实验基础上,2012年于会仙湿地水葫芦的生长旺盛期(6月)和暴发高峰期(8月),在湿地具有代表性的轻度富营养化、中度富营养化和重度富营养化水域中,各选择3个面积为30 m×30 m、有水葫芦入侵的典型水域作为重复,共9个试验小区,随机采集水葫芦,以植株为单位,每个试验小区采集15株. 生物量的计算是将实测样方面积的生物量转换为单位面积的生物量(g/m2)[19]. 按照各个水域面积及水体生态环境特点,采用浮标法确定采样点,同时在不同水域各中心点用采水器采集水样,每个采样点取10 处瞬时混合且深度为40 cm的表层水,样品采集后用保温密封瓶4℃保存带回实验室分析.
参考湖泊富营养化评价方法及分级标准[30-32],依据叶绿素a(Chl.a)、总磷(TP)、总氮(TN)、透明度(SD)和高锰酸盐指数(CODMn) 参数的测定结果,运用综合营养状态指数法对研究区湿地水体健康状况进行评价,综合营养状态指数计算公式为:
TLI(Σ) =ΣWj·TLI(j)
(1)
各参数营养状态指数的计算公式为:
TLI(Chl.a)=10(2.5+1.086 ln Chl.a)
(2)
TLI(TP)=10(9.436+1.624 ln TP)
(3)
TLI(TN)=10(5.453+1.694 ln TN)
(4)
TLI(SD)=10(5.118-1.94 ln SD)
(5)
TLI(CODMn)=10(0.109+2.661 ln CODMn)
(6)
式中,Chl.a单位为mg/m3,SD单位为m;其他指标单位均为mg/L;TLI(Σ)表示综合营养状态指数;TLI(j)代表第j种参数的营养状态参数;Wj为第j种参数的营养状态指数的相关权重,详细见参考文献[30-31]. 综合营养状态指数TLI(Σ)<30为贫营养,30≤TLI(Σ)≤50 为中营养,50

表1 研究区综合营养状态指数特征
参照《水和废水监测分析方法》[33],采用《水质总有机碳的测定-燃烧氧化非分散红外吸收法》(HJ 501-2009)、《水质总氮的测定-碱性过硫酸钾消解紫外分光光度法》(HJ 636-2012)、《水质氨氮的测定-纳氏试剂分光光度法》(HJ 535-2009)、《水质硝酸盐氮的测定-紫外分光光度法》(HJT 346-2007)分别分析水样中总有机碳、总氮、氨氮和硝态氮浓度. 总磷浓度采用浓硫酸-高氯酸消煮,钼锑抗比色法(Agilent 8453 紫外-可见分光光度计,美国)测定;叶绿素a浓度使用 95%乙醇提取后测定;高锰酸盐指数采用酸性高锰酸钾法测定;透明度采用塞氏盘法测定.
在湿地不同富营养化水域中,每小区选择20株水葫芦进行挂签标记,每隔5 d观测一次,记录水域面积和水葫芦抽出的分株数,统计每月的分株总数,然后在生长旺盛期和生长暴发期不同时间间隔内,计算单位面积每天的分株速率大小. 各处理随机选取5株水葫芦,将其捞出洗净后置之于阴凉处沥干,10 min后测定各株水葫芦匍匐茎和叶柄的数目,以新叶叶长达2 cm以上作为判定长出新叶的标准. 记录水葫芦的叶长(LL)、叶宽(LW)、株高(SL)、根长(RL)等形态指标;采用便携式叶绿素仪(SPAD-502PL,日本)测定水葫芦叶片叶绿素含量(SPAD);将水葫芦根、茎和叶分开放入烘箱中在105℃杀青30 min,之后在85℃烘干72 h至恒重,用电子天平(精度0.01 g)称量各器官生物量. 以干重平均值估算生物量[18-19,34],由下列公式计算各器官生物量比. 用表型可塑性指数来解释不同处理下表型可塑性的大小,取值为0~1,其值越大表示表型可塑性越大[35].
分株速率=分株总数/间隔时间
(7)
叶生物量比=叶重/植株总重
(8)
茎生物量比=茎生物量/植株总重
(9)
根生物量比=根重/植株总重
(10)
根冠比=根生物量/地上部分生物量
(11)
表型可塑性指数=(最大值-最小值)/最大值
(12)
1.3 数据分析
采用Microsoft Excel 2016软件整理数据;利用 SPSS 22.0软件对各指标分别进行方差分析. 比较前,先检验数据是否符合正态分布. 对于服从正态分布的数据,使用单因素方差分析(One-way ANOVA)检验不同处理间水葫芦形态和生物量分配的差异(LSD,α=0.05)及显著性;对于不服从正态分布的数据,首先进行数据的转换,转换后若数据服从正态分布,使用单因素方差分析检验相关变量间是否存在显著差异. 若仍不服从正态分布,则使用非参数检验中的 K 个独立样本检验法. 用Canocofor Windows 5.0 软件对不同处理下的水葫芦各形状和环境因子以及生长周期等指标进行主成分分析(principal component analysis,PCA).
2 结果与分析
2.1 不同富营养化试验小区中水体的理化性质
由表2看出,在水葫芦生长旺盛期(6月)和暴发高峰期(8月),水体总有机碳、总氮、总磷、氨氮、硝态氮和叶绿素a浓度在不同富营养化水体间具有显著差异(P<0.05),均表现为重度富营养化水体中的浓度最高,中度富营养化水体次之,轻度富营养化水体最低. 暴发高峰期水体的主要理化指标值高于生长旺盛期,如在暴发高峰期水体的总有机碳浓度比生长旺盛期高53.85%,比中度富营养化水体高56.41%,比轻度富营养化水体高53.26%;在暴发高峰期水体的TN浓度比生长旺盛期高46.77%,比中度富营养化水体高27.93%,比轻度富营养化水体高8.19%. 说明在水葫芦暴发高峰期,水体中过多的营养物质可能加剧了水葫芦的疯狂生长,水体富营养化程度的加重, 导致了水葫芦入侵加剧.

表2 不同富营养化试验小区中水体的主要理化指标*
2.2 不同富营养化水体对水葫芦各类性状的影响
2.2.1 不同富营养化水体对水葫芦株高、根长、匍匐茎数和分株速率的影响 从表3可以看出,在水体同一富营养化条件下,暴发高峰期水葫芦的株高、根长、匍匐茎数和分株速率要高于生长旺盛期. 在暴发高峰期,轻度富营养化水体中水葫芦的株高、根长、匍匐茎数和分株速率分别比生长旺盛期高95.70%、90.74%、200.00%和88.80%,中度富营养化水体中水葫芦的株高、根长、匍匐茎数和分株速率分别比生长旺盛期高102.22%、69.84%、50.00%和31.98%,而在重度富营养化水体中,则分别比生长旺盛期高128.17%、81.10%、80.00%和47.48%. 水葫芦的株高、根长、匍匐茎数和分株速率在同一生长时期不同富营养化水体间存在显著差异(P<0.05). 在生长旺盛期和暴发高峰期,株高和根长均随着富营养化程度的增大而减小,且轻度富营养化水体显著高于中度和重度富营养化水体(P<0.05),如生长旺盛期轻度富营养化水体中水葫芦的株高是中度和重度富营养化水体中的1.27和1.79倍,在暴发高峰期水葫芦的株高是中度和重度富营养化水体中的1.22和1.53倍. 虽然在暴发高峰期水葫芦根长和株高相比生长旺盛期有所增加,但随水体富营化程度的加重,却呈现降低趋势. 相反,匍匐茎数和分株速率却随着富营养化程度的加重而增加,表现为重度富营养化水体>中度富营养化水体>轻度富营养化水体,且重度富营养化水体高于中度富营养化水体,中度富营养化水体高于轻度富营养化水体(P<0.05). 可见,随水体富营养化程度的加剧,水葫芦的响应策略表现为株高和根长减小,呈现低矮化,主要通过增加匍匐茎数和分株速率来提高其繁殖能力,促进其种群面积增大,以提高其对外界环境和资源条件的适应性.

表3 不同富营养化水体对水葫芦株高、根长、匍匐茎数和分株速率的影响
2.2.2 不同富营养化水体对水葫芦叶性状的影响 由表4可知,在不同富营养化水体,水葫芦叶性状存在显著差异(P<0.05),叶柄数、叶柄宽、叶片数、叶长和叶宽均随着水体营养化程度的加剧而增大,如在生长旺盛期重度营养化水体中水葫芦的叶柄数、叶柄宽、叶片数、叶长和叶宽分别是轻度营养化水体的3.00、2.14、1.79、2.33和1.74倍. 叶绿素含量与之相反,随着富营养化程度的加深而降低,在生长旺盛期轻度富营养化水体中水葫芦的叶绿素含量是重度富营养化水体的2.05倍,在暴发高峰期轻度富营养化水体中,水葫芦的叶绿素含量则是重度富营养化水体的1.30倍. 在暴发高峰期的轻度、中度和重度富营养化水体中,水葫芦的叶绿素含量分别是生长旺盛期轻度、中度和重度富营养化水体的2.01、2.33和3.17倍,但均随富营养化程度加重而趋于减小,表明可能主要受水体营养盐水平的影响. 与株高等指标相似,水葫芦的各叶性状指标值在暴发高峰期高于生长旺盛期.

表4 不同富营养化水体对水葫芦叶性状的影响
2.3 不同富营养化水体对水葫芦生物量的影响
2.3.1 不同富营养化水体对水葫芦生物量积累的影响 不同富营养化水体对水葫芦生物量的影响如图2所示. 不同富营养化水体中水葫芦的总生物量分别与根生物量、茎生物量和叶生物量之间差异显著(P<0.05),在生长旺盛期和暴发高峰期,总生物量、根生物量、茎生物量和叶生物量均随着富营养化程度的增加而增大. 在生长旺盛期,重度富营养化水体水葫芦的总生物量(130.23 g/m2)、根生物量(42.71 g/m2)、茎生物量(48.52 g/m2)和叶生物量(39.0 g/m2)比轻度富营养化水体的分别高22.0%、20.49%、19.89%和26.33%,在暴发高峰期,重度富营养化水体水葫芦的生物量分配大小比轻度富营养化水体则分别高22.94%、27.52%、18.71%和24.35%,表明富营养化程度的加重增加了水葫芦的生物量,水葫芦对水体富营养化表现出一定的表型可塑性. 就水葫芦不同器官的生物量而言,在生长旺盛期同一富营养化程度下,水葫芦不同器官生物量积累大小关系表现为茎>根>叶;而在暴发高峰期,生物量积累情况为茎生物量(71.47~87.92 g/m2)最大,叶生物量(51.25~67.75 g/m2)次之,根生物量(43.56~60.10 g/m2)最小,这也说明生物量积累与暴发高峰期水葫芦的匍匐茎数和分株速率较大有关(表3). 可见水葫芦生物量的分配主要趋向于茎部分. 在不同富营养化程度下,暴发高峰期的总生物量(166.28~215.78 g/m2)高于生长旺盛期(101.57~130.23 g/m2),增幅为38.92%~39.88%,其中根、茎和叶生物量的增幅分别为22.04%~28.94%、44.81%~46.88%和42.44%~43.94%. 总体上,水葫芦的生物量在重度富营养化水体中较高,暴发高峰期比生长旺盛期高.

图2 不同富营养化水体对水葫芦生物量积累的影响(图中数据为平均值±标准误,图中左面小写字母a、b、c分别表示生长旺盛期(6月份)轻度、中度、重度富营养化水体间各指标的差异显著性;图中右面d、e、f分别表示暴发高峰期(8月份)轻度、中度、重度富营养化水体间各指标的差异显著性,字母相同表示差异不显著(P>0.05),字母不同表示差异显著(P<0.05). TB表示总生物量,RB表示根生物量,SB表示茎生物量,LB表示叶生物量. 下同)Fig.2 Effect of different eutrophic waters on biomass accumulation of Eichhornia crassipes
2.3.2 不同富营养化水体对水葫芦生物量分配比的影响 在同一生长时期不同富营养化水体间,水葫芦各器官生物量的分配表现出一定差异性(图3). 在生长旺盛期,根生物量比、茎生物量比、根冠比和茎根比在不同富营养化程度水体间均差异不显著(P=0.061),叶生物量比、叶根比和茎叶比间则存在显著差异(P<0.05). 水葫芦生物量在茎中的分配占绝对优势,占全株的37.26%~38.27%,根次之(32.80%~33.44%),叶最少(28.29%~29.94%). 各部位生物量贡献的大小关系依次是茎>根>叶,生长旺盛期水葫芦的生物量贡献主要来自茎和根.
在暴发高峰期,茎生物量比、叶生物量比和根生物量比的范围分别为40.75%~42.43%、30.82%~31.40%、26.17%~28.43%,此时期各部位的生物量贡献的大小关系表现为茎>叶>根,说明该生长时期水葫芦的生物量主要分配在水上部分(茎和叶). 除叶生物量比、叶根比差异不显著之外,其他器官的配比关系在不同营养化程度水体间部分呈现出显著差异,如轻度富营养化水体中水葫芦的茎根比、茎生物量比、茎叶比均显著大于重度富营养化水体(P<0.05),而根生物量比则是轻度富营养化水体显著低于重度富营养化水体(P<0.05). 暴发高峰期水葫芦的叶根比、茎根比和茎叶比均高于生长旺盛期.
在生长旺盛期,根冠比随着水体富营养化程度的增加呈现减小的趋势,大小关系表现为轻度富营养化水体>中度富营养化水体>重度富营养化水体,说明分配给冠部植株的生物量相对较大;暴发高峰期则相反,随着富营养化程度的增加根冠比呈现升高的趋势,重度富营养化水体显著大于中度和轻度富营养化水体(P<0.05),说明分配给根系的生物量相对增加. 而生长旺盛期根冠比大于暴发高峰期,表明生长时期不同,水葫芦根冠的生物量分配有所差异,暴发高峰期分配给冠部植株的生物量相比生长旺盛期要大;这可能是由于水葫芦为快速生长采取的生存策略不同,与在暴发高峰期生物量主要集中在叶和茎部位,用于水平方向的拓展,这与相对应的冠部植株生物量的增加有关,与匍匐茎数和分株速率增加(表3)也是相一致的.
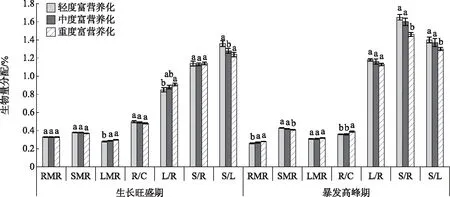
图3 不同富营养化水体对水葫芦生物量分配的影响(RMR表示根生物量比,SMR表示茎生物量比,LMR表示叶生物量比,R/C表示根冠比,L/R表示叶根比,S/R表示茎根比,S/L表示茎叶比)Fig.3 Effect of different eutrophic waters on biomass allocation of Eichhornia crassipes
2.4 不同富营养化水体中水葫芦表型可塑性指数的变化
由表5可知,水葫芦的表型可塑性指数在0.009~0.792之间,各器官的可塑性指数在不同富营养化水体间有明显差异性. 在生长旺盛期,除叶长和分株速率外,其他性状的可塑性指数均在重度富营养化水体达最大值. 从不同富营养化水体中水葫芦形态可塑性指数的平均值来看,叶长的可塑性指数在生长旺盛期最高(0.356),说明该时期水葫芦叶长的可塑性最大;生长旺盛期叶片数的可塑性指数最小(0.168),叶长可塑性指数是叶片数的2.12倍. 在暴发高峰期,除叶宽和叶柄数外,其他性状的可塑性指数均为在重度富营养化水体大于中度和轻度富营养化水体,其中匍匐茎数的可塑性指数最大(0.480),株高的最小(0.113).
生物量分配方面,除茎干重外,在生长旺盛期和暴发高峰期,其他性状的可塑性指数均在重度富营养化水体下达最大值. 叶干重、茎干重和根干重的可塑性指数分别小于叶重比、茎重比和根重比,即水葫芦根干重、茎干重和叶干重的可塑性指数均小于相应的生物量比. 生长旺盛期和暴发高峰期的根冠比可塑性指数均值最小. 总体上,在水葫芦各性状参数中,叶重比的可塑性最大,根冠比最小;不同富营养化水体中重度富营养化的可塑性最大;在不同生长时期之间,表型可塑性没有明显的规律.

表5 富营养化水体中水葫芦形态和物质分配的可塑性指数
2.5 水葫芦表型可塑性与不同富营养化水体间的主成分分析
图4为水葫芦形态指标、生物量分配与不同富营养化水体理化性质间的主成分分析(PCA)(用来表示植物功能性状和环境因子之间的排序图). 图中实心圆形的分布情况表示水葫芦生长时期+营养化程度对植物形态构件的影响情况;虚线箭头连线与排序轴夹角的大小表示环境与排序轴相关性的大小;实线箭头连线与排序轴夹角的大小表示植物形态和生物量指标与排序轴相关性的大小;实线箭头和虚线箭头之间夹角的大小表示该形态指标或者生物量指标与该环境因子的相关性大小;夹角越接近 0°或180°代表两者相关越显著,夹角越接近90°代表两者相关越弱[35]. 图中前两轴解释了总方差的86.78%,表明排序结果是可信的,能够较好地反映不同富营养化下水葫芦性状参数与环境因子之间的关系. 总生物量、根生物量、叶生物量、叶片数、叶宽等均与PCA2轴(解释了86.78%)显著相关(夹角<90°,下同),茎根比、根长、株高和茎叶比均与PCA1轴(解释了62.97% )显著相关. 生长旺盛期和暴发高峰期分布在主成分两轴的两侧,说明水葫芦表现出一定的表型可塑性,并且与暴发高峰期存在较高的相关性. 在生长旺盛期,表型可塑性与茎生物量比和根生物量比具有显著的相关关系. 水体总有机碳、总氮、总磷、硝态氮、氨氮和叶绿素a均与第1排序轴呈显著正相关,同时与茎生物量、叶宽、叶片数、匍匐茎数、叶柄数、分株速率、叶生物量和叶生物量比等显著相关,与根长、株高、叶生物量比和茎叶比等呈显著负相关. 碳氮比与第2排序轴显著正相关,与茎叶比和根长等呈显著相关. 水葫芦功能性状参数不同,与环境因子的相关性有所差异,不同生长时期和不同富营养化水体的理化因子共同显著影响了水葫芦的表型可塑性.
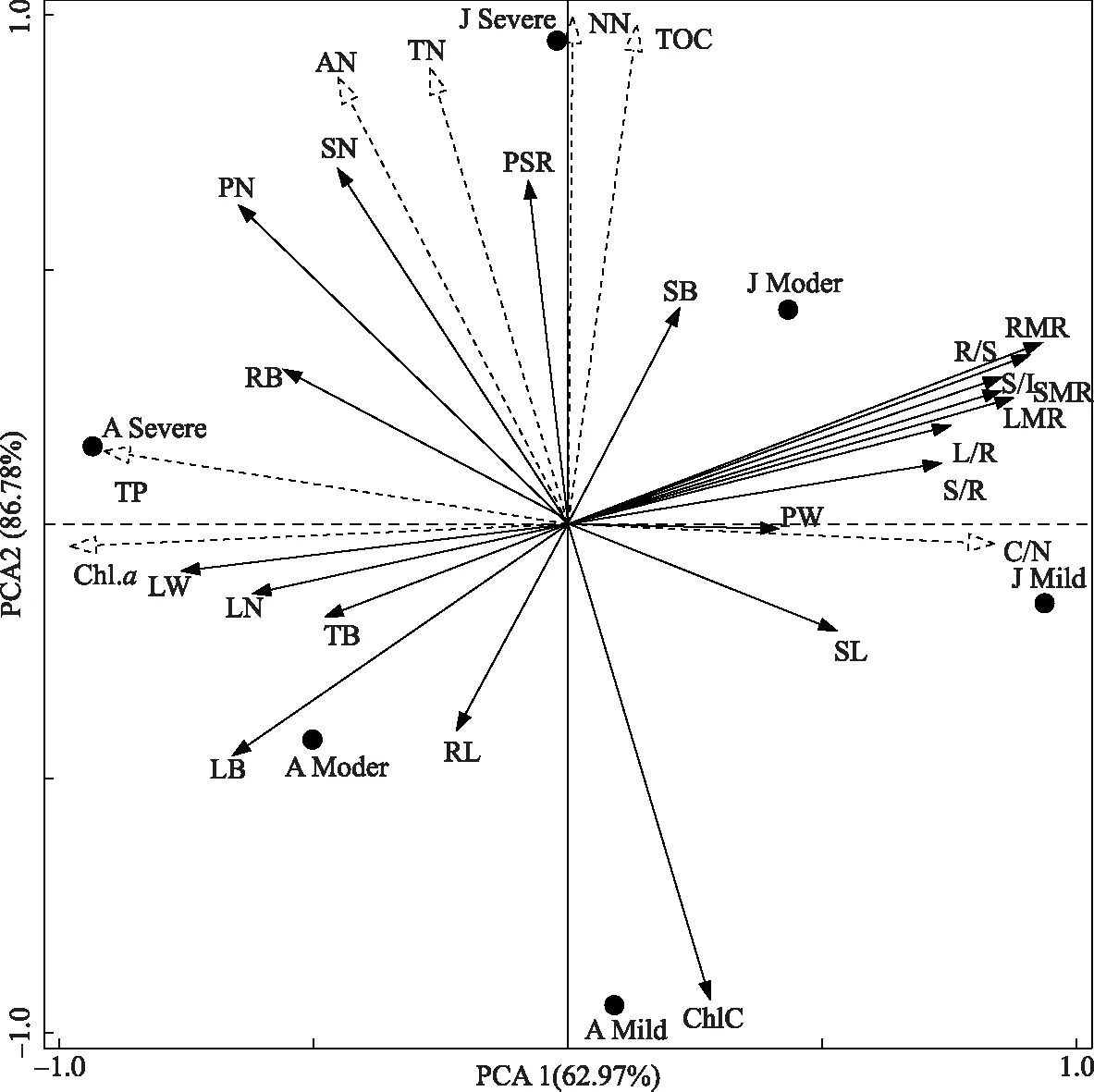
图4 水葫芦形态、生长指标与富营养水体理化性质的主成分分析(PN:叶柄数;PW:叶柄宽;LN:叶片数;LW:叶宽;SL:株高;RL:根长;SN:匍匐茎数;PSR:分株速率;SB:茎生物量;LB:叶生物量;RB:根生物量;TB:总生物量;RC:根冠比;SMR:茎生物量比;LMR:叶生物量比;RMR:根生物量比;L/R:叶根比;S/R:茎根比;S/L:茎叶比;ChlC:叶绿素含量;J表示6月(生长旺盛期);A表示8月(暴发高峰期);Mild、Moder和Severe分别表示轻度、中度和重度富营养化)Fig.4 Principal component analysis of morphology, growth index, eutrophication and physi-chemical property of water for Eichhornia crassipes
3 讨论
3.1 水葫芦株高、根长、匍匐茎数和分株速率对不同富营养化水体的响应特征
在不同的环境条件下,植物对资源的获取和分配能力是其竞争成败的重要因素,而植物的表型可塑性是提高其资源获取能力的重要途径,植物性状(包括地上和地下部分)发生的变化是植物适应异质环境最直观的策略之一[15]. 在各类性状参数中,株高指示了植株的生长情况,与植物邻体间的竞争能力和适应能力有关;根系长度决定了植物所能延伸的深度,直接影响植物对资源的获取能力和竞争力[15];匍匐茎数和分株速率可使植物有选择地将分株置于资源充足的良性斑块,或者逃离资源稀少的劣质斑块[36]. 本研究结果表明,不同富营养化水体对水葫芦株高、根长、匍匐茎数和分株速率的影响有所差异. 随着富营养化程度的增加,水葫芦的株高和根长趋于下降,匍匐茎数和分株速率随之升高,呈现低矮化现象,水平方向的种群扩张面积趋于增大,且重度富营养化水体下水葫芦的匍匐茎数和分株速率显著高于轻度富营养化水体,暴发高峰期高于生长旺盛期. 这与刘晓娟等[37]的观点相似,其指出生活在资源较丰富环境中的植物体具有较高的扩散和定植能力,以快速抢夺生存所必需的资源,因而植物体会将更多的能量分配给繁殖和生长;与之相反,生活在资源限制中的植物体会将能量分配给维持植物在不利生境中的结构. 刘旻慧等[26]的研究表明,当环境中营养盐浓度较低时,水生植物可通过提高根系中相关酶活性,满足自己对营养的高效吸收与利用. 在会仙喀斯特湿地,随着水体富营养化程度的加剧,虽然植株高度相对有所降低,但没有抑制水葫芦的生长,也没有减少水葫芦分枝数量,匍匐茎数和分株速率与水葫芦的繁殖密切关联. 在重度富营养化水体中,水葫芦将营养物质主要投入到增加匍匐茎数和分株速率方面,力争进行种群的扩张,以增加种群面积和占据更为有利的生境,尤其在暴发高峰期会出现水面分布面积剧增的现象.
高岩等[38]的研究结果表明,水葫芦既可以吸收水体中的营养物质,又可通过根系吸收底泥中的营养. 与其类似,本研究中,在轻度富营养水体中,水葫芦的响应策略为增加其株高和根长,随着富营养化程度的增大,虽然轻度富营养化下的营养物质相对较少,但其较高的株高有利于获取更多的光能,而且根长的增加有利于伸入水体吸取更多的营养物质. 水葫芦可通过株高、根长、匍匐茎和分株速率等表型可塑性特征来调节对富营养化水体养分的利用以促进其自身对必需资源最大限度地获取. 研究表明,植物的叶长、叶宽、叶柄长、叶片数和叶面积等叶性状参数可随着海拔、水分、光环境和温度等环境因子的变化而变化[38-40]. 而Gallagher等[41]的研究指出,并没有证据证明植物的叶片性状与入侵植物的生存能力存在联系;同一种植物,入侵种和本土植物在叶片性状上没有显著的差异. 本研究中,水葫芦叶柄数、叶片数、叶长和叶宽均随着水体富营养化程度的加深而增大,这与陈勇等[40]的结论相似;叶绿素含量则表现为随着富营养化程度的加深而降低,表明水葫芦植株体可能受到了高富营养化的胁迫,这与其株高和根长趋于降低类似. 在氮、磷浓度胁迫下,植物能够对其有所反应,并通过自身的调节机制,做出形态和生理上的适应性响应策略,以增加其在外界胁迫条件下的生存机会[42]. 植物叶片表现出的差异性,一方面可能与植物的生物学特性有关;另一方面,在重度富营养水体中,水葫芦为了在营养充足的基础上快速扩大其种群面积,通过优化叶长、叶宽、叶柄数、叶炳宽和叶片数等结构来满足自身最大限度地获取养分,以保持在群落中的最大适合度,其叶重比、叶长的可塑性指数表现为较大,因为适合度趋向于随着环境中营养物质总量的增加而增大[43]. 另外,叶柄数的增加有利于水葫芦叶片的运动以吸取更多的营养物质. 在会仙喀斯特湿地,水葫芦的叶片性状随水体富营养化程度的增加反映了其响应环境变化的生存策略,水葫芦的叶性状具有较强的可塑性,且促进了水葫芦在不同富营养化水体中的生态适应性.
3.2 水葫芦生物量分配及表型可塑性对不同富营养化水体的响应特征
生物量的分配格局是植物关键的形态特征之一. 研究表明,入侵植物比本土植物具有更强的可塑性响应[44],生物量的重新分配(从地下到地上)和较强的可塑性有利于植物在频繁干扰的环境中入侵成功[45],形成有利于自身生长发育的适应策略[46]. 水体中含有的营养物质是水生植物生长发育的重要生态因子,水生植物为了实现对养分提取有效性和摄取效率的最大化,必然改变其构件生物量的分配以适应水中养分的差异,从而形成多种因水中养分的异化所引起的能量投资策略[47]. 本实验比较了3个不同富营养化水体中水葫芦生物量积累的变化趋势,发现水葫芦生物量积累随着外界环境的变化而变化,总生物量、根生物量、茎生物量和叶生物量在重度富营养化程度水体中均显著高于中度和轻度富营养化程度水体,水葫芦在生长旺盛期的生物量分配大小关系为茎>根>叶,在暴发高峰期的则为茎>叶>根,茎生物量大于根和叶,水葫芦生物量分配比例的变化,有利于其在不同富营养化水体中的生长以达到最佳状态. 水葫芦将较多的能量分配到茎部,表现为茎生物量的增加,有利于占据更多的空白生境,在外界环境营养充足的前提下扩张其种群面积. 有研究表明,营养物质的增加将会使陆生植物风毛菊(Saussureanivea)的根生物量减少,叶生物量增加[48],与此相类似,本研究中暴发高峰期水葫芦根生物量积累较少. 这可能是在高养分时,水葫芦作为浮水植物,把更多的资源分配于茎叶等器官的生长,相对减少了对根系长度的营养物质投入,而在低养分时,水葫芦却具有较高的根生物量,这有利于从水体或底泥中吸收养分,提高其生存能力.
植物的性状间存在比较普遍的权衡关系,主要包括叶长与叶宽等叶片性状间、叶片与枝条及树干性状间、叶片与根性状间、树干与根性状间、繁殖性状与数量间等[49]. 本研究中,不同富营养化水体和不同生长时期对水葫芦性状间的权衡关系均有一定的影响,在生长旺盛期,不同富营养化水体间的根生物量比、茎生物量比、根冠比和茎根比没有显著差异;但在暴发高峰期,除叶生物量比和叶根比无显著差异外,其他器官间均具有显著差异,在轻度富营养化水体较高. 同一种植物在不同的环境和时期会有多种表型可塑性,以充分有效地利用资源,高效的适应环境[50]. 不同生长时期的水葫芦在不同富营养化程度下的响应策略和表型可塑性表现出差异性,这也是其为适应环境变化而调整自身行为的表现. 包宇等[49]的研究表明,根冠比随着养分的增加而降低;林超等[51]的研究得出,沉水植物苦草的根冠比随着营养盐浓度的增加而升高. 本研究中,水体富营养化程度的升高加剧了水葫芦快速生长,这与陈潇等[7]的研究结果一致. 在不同生长时期,水葫芦为适应生长而对根系和地上植株的资源分配采取了不同的生存策略.
在生长旺盛期,根冠比呈现出随着富营养化程度升高而降低的趋势,在暴发高峰期则呈现出随富营养化程度升高而升高的趋势,根冠比在暴发高峰期小于生长旺盛期,说明在暴发高峰期根系分配生物量相对减少,而水葫芦冠部植株生物量增加,水葫芦通过器官生物量分配对不同富营养化水体产生了可塑性响应,而表型可塑性提高了水葫芦应对环境胁迫的能力,以适应外界高营养的胁迫. 这也反映了水葫芦生长与生物量分配对水体富营养化环境的内在协调性及其适应性,与挺水植物芦苇对盐碱胁迫的可塑性响应相似[52]. 植物表型可塑性是同一基因型植物在面对不同环境条件下,通过调节自身性状特征以适应环境变化的能力[35],同时环境因素也会引起植物形态可塑性的差异[53]. 多个入侵实例表明,表型可塑性强的外来种通常更容易形成入侵[54],而湖泊富营养化有利于外来水生植物的生长与无性繁殖,增强其入侵性,如水葫芦的匍匐茎长度会随营养水平提高而增加[55]. 本研究中,水葫芦能适应富营养化的生境,表现出较强的表型可塑性,这与湿地外来植物香菇草(Hydrocotylevulgaris)相类似[56]. 在同一生长时期不同富营养化水体中,水葫芦的叶长、匍匐茎数、叶柄数和叶宽等以及生物量的可塑性指数不同,重度富营养化下的表型可塑性指数比中度和轻度高,表明重度富营养化水体中水葫芦的表型可塑性比中度和轻度富营养化水体大,这可能与重度富营养化水体环境中营养元素含量高有关. 主成分分析显示(图4),水体总有机碳、总氮、总磷、硝态氮、氨氮和叶绿素a均与水葫芦茎生物量、叶宽、叶片数、匍匐茎数和分株速率等参数具有显著的相关性,水葫芦表型可塑性与水体富营养化程度关系密切. 这与众多研究表明植物的表型可塑性受总氮、总磷、水解氮、速效钾等因素影响的结论相似[51,57-58],入侵植物较本土植物常表现出较高的生长速率、资源利用效率、表型可塑性和环境胁迫耐受能力等,有利于取得竞争优势[59]. 在水生生态系统中,多数成功入侵的外来植物均为具有较强克隆繁殖能力的克隆植物[18],而环境因子是通过怎样的配比使水葫芦在不同富营养化水体中处于最佳的生长适合度,还需进一步的探讨. 会仙喀斯特湿地地处亚热带季风气候区,夏季高温多雨、冬季寒冷少雨的气候特点适合水葫芦生长,其水文系统具有的地表、地下双重空间结构,湿地的水情多变,水文动态变化快,每年漓江流域内其他水流及旅游干扰携入的悬浮有机物等带来的营养物质亦可能加剧水葫芦的大量繁殖.
在对会仙喀斯特湿地水葫芦进行控制和治理时,可根据其在不同富营养化条件下的表型可塑性响应特征,有针对性地采取不同措施来控制其生长、繁殖和扩张. 本研究是探讨基于水葫芦的形态性状(根长、株高和叶片等)、克隆性状(匍匐茎数和分株速率)以及生物量的积累和分配等表型可塑性对不同富营养化程度水体的响应特征,而植物的许多性状普遍存在可塑性[46],为更全面地了解水葫芦在不同富营养水体中的生长策略,还需结合水葫芦在光合作用以及物候、生活史、异质光照下的克隆整合、种子数量及其活力等多方面的相关反馈,特别是根系和叶片的解剖结构、基因组学的分子机制,以便更深入地反映水葫芦对不同富营养化程度水体的入侵机制.
4 结论
1)在会仙喀斯特湿地水葫芦的生长旺盛期和暴发高峰期,水葫芦可以通过形态、生长变化和生物量分配等进行资源分配,不同富营养化水体中的水葫芦产生了有利于其存活和入侵的表型可塑性响应特征. 水葫芦的总生物量、根生物量、茎生物量和叶生物量的大小关系表现为重度富营养化水体>中度富营养化水体>轻度富营养化水体,茎生物量大于叶生物量和根生物量;水葫芦在不同富营养化水体中的表型可塑性提高了其在富营养化水体环境中的生态适应性.
2)在水体轻度富营养化条件下,水葫芦通过增加其株高和根长来调节自身生长;重度富营养化条件下水葫芦的响应策略则表现为提高其匍匐茎数和分株速率,增强其克隆生长能力,提高扩散能力. 水葫芦叶重比的可塑性指数最大,根冠比最小;水葫芦在重度富营养化水体中的表型可塑性最大. 不同富营养化水体显著影响了水葫芦的表型可塑性,水葫芦的表型可塑性与湿地水体总有机碳、总氮、总磷浓度等环境因子密切关联. 水体富营养化程度的提高和水葫芦的表型可塑性共同加剧了水葫芦在会仙喀斯特湿地的入侵.
致谢:感谢刘建春、于倩倩、郭永红、田垒等在实验样品分析方面提供的帮助.
猜你喜欢
散文诗世界(2022年5期)2022-05-23
昆明医科大学学报(2022年1期)2022-02-28
农家参谋(2021年10期)2021-12-05
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
林业与生态(2019年3期)2019-05-21
今日农业(2019年13期)2019-01-03
红蜻蜓·低年级(2017年12期)2018-03-06
东方教育(2016年4期)2016-12-14
学生天地·小学中高年级(2015年11期)2016-01-14
