
植物表型可塑性对环境因子的响应研究进展
2021-01-12 15:52:56朱润军李仕杰杨畅宇程希平
西南林业大学学报 2021年1期
朱润军 杨 巧 李仕杰 杨畅宇 程希平
(西南林业大学地理与生态旅游学院,云南 昆明 650233)
植物的表型可塑性是同种基因型对不同环境因子响应而产生的不同表型特征[1]。植物通过表型可塑性适应不同的环境,可塑性的强弱是衡量植物适应环境能力的指标之一,当前已成为生态学研究的一个重要领域[2]。表型可塑性强的物种通过调整形态、生长、生物量分配等特性,在不利条件下获取更多的环境资源,增强自身对环境的适应能力,从而拥有更广的生态幅[3],特别是在气候变化的大背景下植物表型可塑性对气候变化的响应与适应具有十分重要的意义。另外,表型可塑性包括形态、生理、生活史特征等方面,可以表现在单株的植物上,也可跨越世代[4]。基因和环境都会引起表型的变异,促使植物可塑性响应的非生物环境因子包括光照、水分和土壤养分等;生物环境因子包括竞争、动物采食及微生物等[5]。植物的表型结构是各个构件在空间上的排列、组合,是基因与环境共同影响的结果,是自身生长与环境胁迫相权衡的表现,植物的可塑性研究是通过观察与实验来认识和理解不受基因影响的外部生长过程[6]。植物通过表型结构的变化,增强其对环境资源的获取能力,提高生长适应度。研究植物的空间结构与构件特征,并反映植物的生长进程与环境的关系是植物表型可塑性研究的一个重点方向。
植物的表型受环境的影响,如何准确把握不同生境条件下植物的表型可塑性,是当前本学科的一大热点问题。本研究着重阐述了有关植物表型可塑性对气候、地理以及生物等主要环境因子的响应,为植物物种对环境变化的适应策略提供参考,为理解植物的生存策略提供参考依据。
1 可塑性对气候环境的响应
植物的可塑性与环境的联系十分紧密,性状的调整是其适应环境变化的有效途径,也是最直观的表现之一。植物表型可塑性的研究通常以植物个体为对象,在一种或多种影响植物生长发育的主要环境因子下,观察、记录其表型发展趋势[7]。表型可塑性研究的实验方法有很多[8],主要有5种常用实验方法,其主要优缺点见表1。

表1 表型可塑性研究的实验方法Table 1 Experimental method of phenotypic plasticity research
植物性状的形成是外界环境作用的结果,光照和降水作为重要的气候因素,对植物的可塑性产生直接影响。光照能显著影响植物株高,分枝特征、叶片分布等植物形态特征,一般来说,植物的叶片大而薄且柔软,比叶重较小,叶柄较长等特点是其应对弱光环境的策略。如林窗环境下,闽楠(Phoebe bournei)幼林将更多的生物量分配给枝叶,增大叶面积,并促进冠长、冠幅和侧枝的生长[9]。植物叶片的功能结构也会发生调整,新疆猪牙花(Erythronium sibiricum)在弱光环境下形成较弱的叶表皮角质层,叶片大而薄,叶肉有海绵组织等[10]。光照的不足,往往使植物出现基径变细、株高增加的现象,将更多的生物量用于垂直方向的生长。植物还可以通过水平生长的方式扩大冠幅,最大限度的截获有效光合辐射[11]。在光照充足的条件下,植物则增加对地下部分生物量的投入,增加对水分和养分的利用,因此,光照环境变化是特定性状形成的重要原因之一。
水分也可显著影响植物的生长发育和形态特征,缺乏土壤水分会延缓或抑制植物的正常生长。随着水分的减少,植物倾向于增加根部的生物量,向更深的地下延伸以汲取足够的水分。水分长期匮乏时,植物总叶片数降低、叶片缩小甚至脱落,防止水分过度散失。节节麦(Aegilops tauschii)缺水时茎、叶生物量下降,降低水分蒸发;而根生物量增加,拓展地下空间以获取足够的水分[12]。干旱胁迫减弱了植物的基径、株高和叶片的生长,干旱程度越高,对其生长的抑制作用越显著,这是缺失水分带来的负面影响。而水分过多同样会阻碍植物正常生长,如持续水淹的中华蚊母树(Distylium chinense)的比叶面积显著降低,叶片将多数干物质用来形成保卫细胞,增加叶厚,阻止叶片外部水分进入内部,维持叶片水分平衡[13]。水分的盈亏不仅影响植物的生长,还对表型的重构具有一定的作用。
温度和CO2浓度作为气候环境的一部分,对植物可塑性产生的影响也不可忽视。增温促进植物积累更多的生物量,加速早稻(Oryza sativa)的生长[14]。长期处于较低温度下的植物,叶片与茎的生物量降低,而比叶面积和叶片厚度均比高温下的小和厚[15]。在某些特定条件下,主动和被动的可塑性响应可能会同时产生。较高的CO2浓度提高了植物的光合速率,消耗了更多的水分,而植物通过降低保卫细胞膨压,减小气孔开度降低蒸腾速率的方式,又减少了水分的耗损,使自身水分维持平衡[16]。
2 可塑性对地理环境的响应
植物可塑性不仅受到气候干扰,其所生长的地理环境也会带来一定影响。可塑性取决于具体的物种和环境。其中一个关键特征是响应可预期,并间接地遵循于基因的协调。因生境限制而导致的表型改变,有利于种群或个体的生存和繁衍。
矿质元素作为重要的地理因素在植物生长过程中不可或缺,养分缺失将严重影响植物正常发育。植物在高养分下叶片长度和宽度明显高于低养分,其他部分生物量也随之增加[17]。随着氮、磷水平的上升,飞机草(Chromolaene odorata)通过产生较长的分枝和叶片来形成、扩大自身的冠层结构,大大提高光合作用面积[18]。随着养分在植物体内的富集,会进一步促进植株发育生长,生物量、株高等均显著提高。黄顶菊(Flaveria bidentis)根、茎、叶的生物量和叶面积指数均与土壤氮含量成正比[19]。而且,氮的形态也会对植物生长产生一定的影响,如与同样施加氮元素,使用有机氮与无机氮的植株相比,根冠比增大,生物量分配也存在差异[20]。有些特征的响应是适应性的,而另一些则是对资源匮乏的消极响应。在养分匮乏时,植物与在缺水环境下有相似的表现,即叶片小而厚、较致密且比叶重较大[21]。
地理空间差异也会对植物表型产生影响,如文冠果(Xanthoceras sorbifolium)果实大小与纬度呈正相关[22]。从沿海到内陆,酸枣(Ziziphus jujubavar.spinosas)根初生结构表皮细胞的厚度和宽度逐渐增大[23]。即便是相同特征,在面对同一地理环境因子时也可能表现出不同的响应趋势(图1)。植物的比叶面积在高海拔地区缺水条件下增加,低海拔缺水条件下则减小[24]。另外,环境污染也会对植物生长产生影响,如煤粉尘的加剧,显著降低了羊柴(Hedysarum laeve)幼苗的叶片数和株高[25]。植物性状时刻进行着权衡,并随环境变化做出的相应的调整,有利于种群或个体的生存和繁衍。
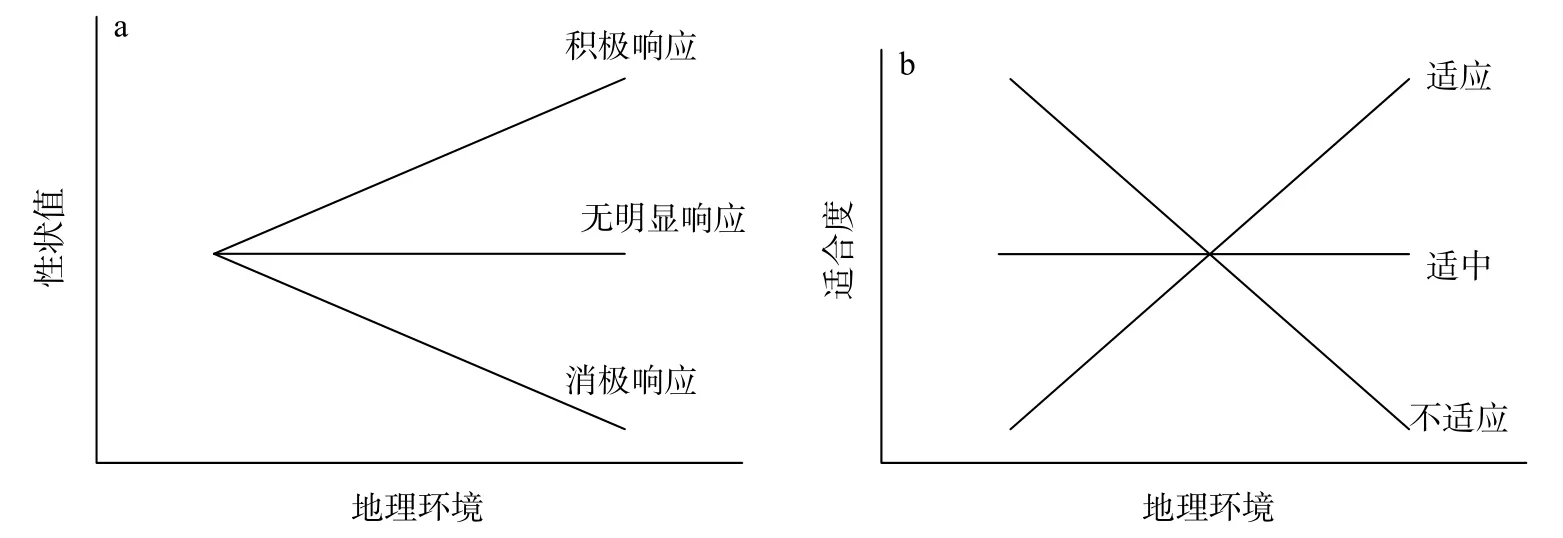
图1 植物性状值、适合度与环境的关系Fig.1 The relationship between plant trait value or fitness and environment
3 可塑性对生物环境的响应
植物通过自身可塑性的机制抵抗、促进或缓冲环境带来影响的权衡结果,促使不同物种占据不同的生态位,有利于生态环境多样性的维持。生物环境对可塑性的影响主要体现在植物间的竞争以及动物采食等。竞争者的出现可能加强了植物对光的争夺,但养分分流会削弱某些性状的发育,如混生小子虉草(Phalaris minor)株高大于单生种群,但分枝数、主穗长度和剑叶面积均比单生种群小[26]。植物可塑性存在种间差异,若外来植物的可塑性强于本地植物,就会占据更多的资源,获得生长优势,出现植物入侵现象。加拿大一枝黄花(Solidago canadensis)具有多而长的根茎向周围延伸从而占据更多的资源,提高了适合度,这可能是其具有较强入侵能力的重要原因[27]。入侵植物通过灵活调整各构件资源的分配,更多地获取有限的资源。成功入侵的原因预计有两种:一是入侵种可以快速适应不同的环境,其表型可塑性明显高于本地种群,斜率比本地种更陡;二是入侵植物与本地种对异质环境的适应速度大致相同,但其性状适合度始终高于本地植物(图2)。植物任何与适应性相关的表型可塑性,在其入侵中发挥着巨大作用。

图2 入侵植物与本地植物可塑性变化趋势Fig.2 The change trend of invasive plants and native plants plasticity
动物的行为同样会诱发可塑性的表达。植物往往会调整叶片形态和化学性状,例如模拟天敌采食则降低了薇甘菊(Mikania micrantha)的叶面积及叶片干质量[28]。放牧强度的加剧,导致植物的株高、叶面积和叶干质量均显著降低,并且出现俯卧生长现象,这些都是主动适应的策略[29]。与植物共生的微生物也可能对可塑性产生影响。接种吡咯伯克霍尔德氏菌(Burkholderia cepacia)的植株生长与光强呈正相关,原因可能是微生物对光同化产物有更高的要求[30]。有益菌还减轻了高粱(Sorghum bicolor)、小麦(Triticum aestivum)和玉米(Zea mays)植株干旱胁迫以及小麦植株盐胁迫的有害影响,并促进茎长、生物量和叶绿素含量的增加[31-32]。由于种间或种内竞争,以及对适应生境的获取,在不同程度上改变植物表型可塑性的表达,以促进种群演化发展。
4 展望
表型可塑性是植物为适应不同环境条件在生理和形态上发生的变化,表现出积极或消极的响应。由于研究描述植物形态结构的直观性,越来越受到关注,并且对理解环境与植物之间的相互作用有着积极的意义。植物在各个器官上生物量分配的格局差异以及表型上的不同,体现了植物对不同环境的适应策略。在植物的生长过程中,光照成为制约植株生长的主要因子时,植物通过增加枝叶的相对生长来获取更多的光资源。水分或养分成为主要的制约因子时,植物则增加根系的生物量分配以最大程度汲取土壤中的水分或养分,植物生物量的分配方式与最优分配假说一致[33]。对于其他环境因素,植物同样会产生可塑性响应增强对环境的适应能力,有利于生长和繁殖,从而拥有更广的生态幅。
在全球气候变化的背景下,植物将会面临光照、水分等环境因素的时空挑战。目前植物可塑性的研究虽取得了长足的进展,但仍然有很多问题需要解决。基于此前研究中存在的不足,今后应在以下几个方面加强研究:1)自然环境复杂多样,不同环境因子往往交互影响,多种环境因子对植物可塑性的综合影响需要进一步探索,另外还应加强对不同时间、空间植物可塑性的研究。2)植物的生长对策会随着环境的变化而改变,则需要进一步探究植物可塑性变化的动态过程与响应机制。3)当前植物可塑性的研究主要停留在宏观方面,在分子、细胞层面上对其调控的机理还需深入探讨。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
新疆农业科学(2020年1期)2020-02-14 03:22:46
现代园艺(2017年21期)2018-01-03 06:41:32
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国麻业科学(2015年5期)2015-12-28 06:22:11
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
西南医科大学学报(2015年1期)2015-08-22 13:01:48
