
科尔沁不同沙地类型植被动态特征及其与凋落物的关系研究
2021-01-09 06:42陈银萍李晓辉罗永清王旭洋牛亚毅
草地学报 2021年1期
陈银萍,李晓辉,罗永清,王旭洋,牛亚毅
(1.兰州交通大学环境与市政工程学院,甘肃 兰州 730070; 2.中国科学院西北生态环境资源研究院,甘肃 兰州 730000)
凋落物是植被-土壤系统物质循环和能量流动的关键环节[1]。植被通过凋落物的产生将存储于体内各器官的有机物质释放到环境中,通过风力作用实现凋落物的再分配,在水热等环境因素的影响下,环境中的微生物将凋落物进行分解,使存储于凋落物中的有机物质再次转化为无机物或小分子有机物释放到环境中,并通过根系吸收,再次被植物利用[2]。因此,在一定程度上,地表凋落物的量决定着土壤肥力的大小和植被生长状况。植被特征是影响凋落物产生的主要因素,植物生物量对凋落物的量有一定的决定作用[3]。北京东林山地区的研究表明,氮添加可通过促进灌木生长从而导致凋落物产量增加[4];中国森林凋落物的对比分析表明,温度、降水等环境因素与森林总凋落物及其组分均呈显著的正相关关系[5]。另外,凋落物的再分配过程是植被-土壤系统物质循环的关键,如凋落物形成过程中的风力搬运是影响地表凋落物量的重要因素。已有研究表明,在科尔沁沙地流动、半固定和固定沙丘,植被生长状况较好的丘间低地,凋落物运移量显著低于迎风坡和坡顶[6]。因此,植被盖度和密度可能通过调节近地面微环境,减少风对凋落物的搬运,影响着地表凋落物的积累。但目前的相关研究中,对风沙活动剧烈、草本植物为主的干旱半干旱地区的植被与凋落物关系的研究较少。
科尔沁沙地是我国四大沙地之一,由于人为活动的干扰,沙漠化土地面积由20世纪50年代的20%增加到80年代的53%。从20世纪80年代后期开始,由于土地利用方式的调整和沙漠化治理措施的加强,土地沙漠化逐步实现逆转[7-8]。在该过程中,由于沙漠化治理措施和土地利用方式的差异,以及本世纪以来气候特征变化(气温整体升高、降水分布不均、极端气候事件增加等)的加剧[9],该地区景观类型多样化,流动沙丘、半固定沙丘、固定沙丘等景观单元交错分布。这些景观单元特殊的分布格局导致科尔沁沙地植被-土壤与物质循环格局的异质性加剧。而作为植被与土壤物质循环的重要环节,凋落物的产生和分配也受到这种格局的强烈影响[6]。不同类型生境的凋落物[10-11]、植被[12]和土壤性质[13]等均存在显著的差异性。目前的研究大多关注于土壤性质[13-15]、植被特征[12]及二者的关系[16-20],而凋落物方面的研究,尤其是其与植被关系方面的研究相对较少。
综上所述,本研究以位于科尔沁沙地东南缘的奈曼沙漠化研究站长期监测样地(流动沙丘、固定沙丘和封育草地)为对象,研究了2005-2016年间凋落物量与植被特征的年际动态,旨在分析当地植被-凋落物关系,为生态恢复提供一定的数据支持和理论依据。
1 材料与方法
1.1 研究区概况
本研究区位于我国东北部的科尔沁沙地东南缘,地处内蒙古自治区东部的通辽市奈曼旗(42°58′ N,120°43′ E)境内,平均海拔345 m。该地区属于温带大陆性半干旱气候,多年平均降水量(1961-2017年)为343 mm,年均降水量波动性较大,变异系数为25.7%,其中86.3%发生在5-9月份。年均温为6.7℃,月平均最高和最低气温分别为24.3℃(7月)和-12.6℃(1月),年均蒸发量1 935 mm。土壤类型为风沙土。植被类型包括灌木黄柳(salixgordejevii)、差巴嘎蒿(Artemisiahalodendron)和小叶锦鸡儿(Caraganamicrophylla),一年生草本沙米(Agriophyllumsquarrosum)、狗尾草(Setariaviridis)、五星蒿(Bassiadasyphylls)、大果虫实(Corispermummacrocarpum)、猪毛菜(Salsolacollina)和黄蒿(Artemisiasacrorum)等,多年生草本主要有糙隐子草(Cleistogenessquarrosa)、达乌里胡枝子(Lespedezadavurica)、羊柴(Hedysarumfruticosum)和冷蒿(Artemisiafrigida)等。地貌以平缓的流动沙丘、半流动沙丘、固定沙丘、丘间低地和甸子地镶嵌分布。
1.2 研究方法
本研究在中国科学院奈曼沙漠化研究站流动沙丘辅助观测场(流动沙丘:MD)、固定沙丘辅助观测场(固定沙丘:FD)和沙地综合观测场(封育草地:G)中进行。植被组成的测定方法为:在3种类型的观测场中分别随机设置10个10 m×10 m的大样方,每个样方各设置1个1 m×1 m的小样方,于生长季的高峰期(8月下旬)进行植被调查,分别统计样方内植物的株数和每种物种的盖度,统计结束后齐地面收割地上活体部分,带回实验室内冲洗烘干(65℃烘干48小时)计算每个样方的地上活体生物量。凋落物观测样方设置与植被组成测定方法一致,即在每种类型的观测场随机设置10个10 m×10 m的大样方,每个样方各设置1个1 m×1 m的小样方,在每年生长季开始前(4月底),收集地表所有凋落物,清除立枯物和植物活体部分,然后做好标记,随后逐月回收进入各样方的凋落物,将收回的凋落物冲洗干净,然后56℃烘干48小时,测定生物量。本研究共计调查植被组成样方360个(3类样地×10样方×12年),凋落物样方330个(3类样地×10样方×11年,2006年数据缺失)。
1.3 数据分析
采用SPSS 20.0进行数据分析,不同类型样地的植被特征采用单因素方差分析(one-way ANOVA)进行,LSD法进行多重比较,不同样地植被组成的年际动态进行二因素方差分析(two-way ANOVA);不同样地凋落物特征采用三因素方差分析(three-way ANOVA)进行;采用线性回归分析(Linear Regression Analysis)法分析植被特征与凋落物量的关系,不同样地植被特征与凋落物量的关系通过相关分析(Pearson Correlate Analysis)完成。
2 结果与分析
2.1 植被特征
2005-2016年的平均值分析表明,3类样地的植被特征存在极大的差异性。流动沙丘样地植被密度显著低于固定沙丘样地和封育草地(P<0.05),后二者的差异性不显著(图1-A)。3类样地的植被盖度间均存在显著差异性(P<0.05),其中流动沙丘样地植被盖度最低,其次为固定沙丘样地,封育草地最高(图1-B)。地上生物量与盖度类似,3类样地间均存在显著的差异性(P<0.05),具体表现为从流动沙丘到封育草地,生物量呈现逐渐增加的现象(图1-C)。
年际动态分析表明,植被密度、盖度和地上生物量均具有强烈的年际波动性。2005年的植被密度较高,随后几年植被密度较低且波动较小,2010-2013年间的植被密度增加,2013年达到最大值(328.3株·m-2),此后逐渐下降(图2-A)。与密度相比,植被盖度的波动性较小,最大盖度为最小盖度的1.64倍,从12年的年际动态来看,植被盖度整体表现为两个明显的增减过程,第一个出现在2005-2008年,第二个出现在2009-2015年,其中2012年的植被盖度最大(图2-B)。与盖度类似,植被生物量的波动性较小,最大生物量和最小生物量分别为141.94 g·m-2和50.48 g·m-2,其中2005—2008年,植被生物量呈现明显的下降趋势,2008年的生物量最小,随后逐年增加,2012年达到最大,此后再次逐渐下降(图2-C)。
不同类型样地植被特征的年际动态分析表明,植被密度、盖度和生物量均存在显著的年和样地类型的2因素交互效应,表明各类样地的年际动态呈现出不同的特征。固定沙丘和封育草地样地的植被密度具有相似的年际动态,整体呈现“降-升-降”的特征且波动范围较大,最大密度均出现在2013年;而流动沙丘样地的波动性较小,最大密度出现在2014年(图3-A)。固定沙丘和封育草地样地植被盖度的年际动态整体呈现为“M”型波动且波动范围较大,其中固定沙丘样地在2005和2013年的盖度较高,封育草地样地在2006和2012年的盖度较高;而流动沙丘样地植被盖度的波动范围较小,且整体呈现为逐年增加的趋势(图3-B)。封育草地样地生物量的年际波动最大,整体呈现倒“V”型特征,2012年的生物量最大,随后逐渐下降;固定沙丘样地的波动性较封育草地小,其中2005-2011年呈现缓慢下降的趋势,随后又表现为“增-减-增”的趋势;流动沙丘样地的波动性最小,其中2005年的生物量最大,此后趋于稳定,生物量的变化范围为5.27~14.77 g·m-2(图3-C)。
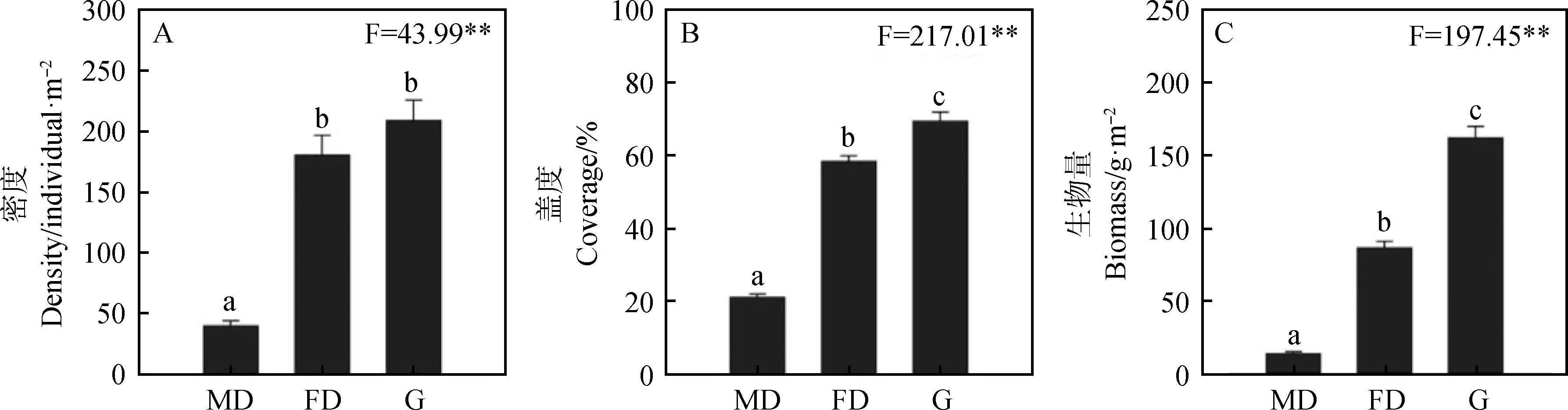
图1 科尔沁沙地植被特征的生境差异Fig.1 Vegetation characters among different habitats in Horqin Sandy Land注:不同生境MD、FD和G分别表示流动沙丘、固定沙丘和封育草地,不同小写字母表示差异显著(P<0.05),**表示差异显著(P<0.01),下同Note:Different habitats of MD,FD and G in figures indicated mobile dune,fixed dune and fencing grassland,respectively. Different lowercase letters indicated the differences was significant at the 0.05 level,** indicate signifiacnt difference at the 0.01 level, the same as below
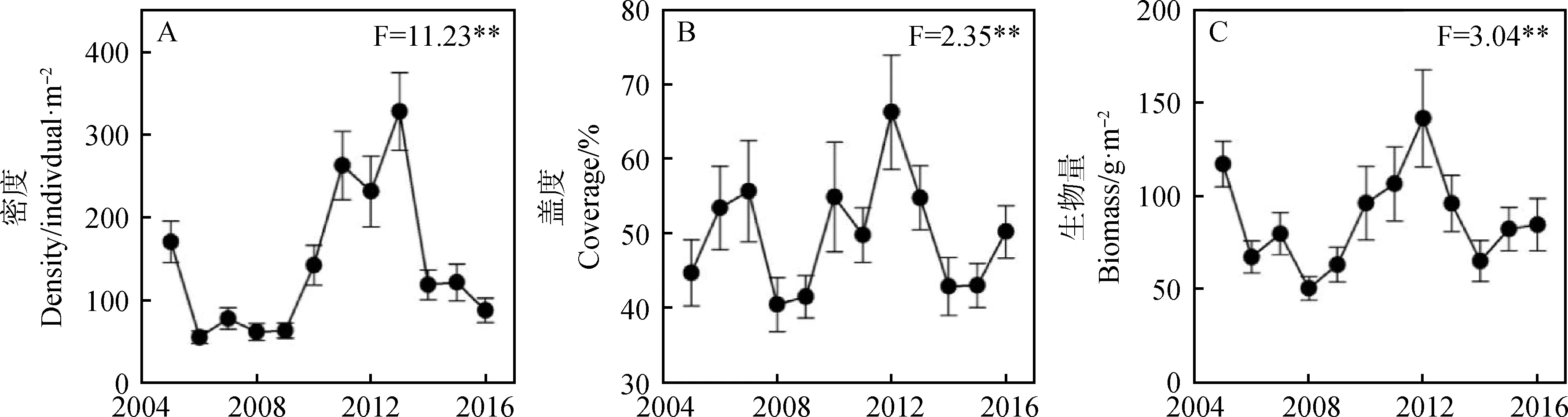
图2 科尔沁沙地植被特征的年际动态Fig.2 Interannual dynamics of vegetation characters in Horqin Sandy Land
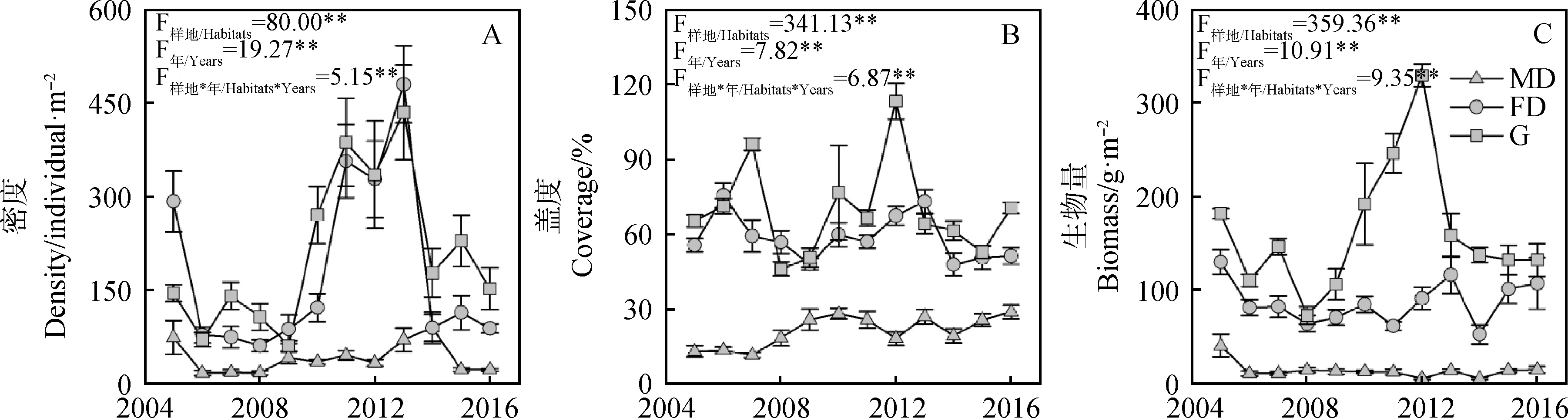
图3 科尔沁沙地不同类型样地植被特征的年际动态Fig.3 Interannual dynamics of vegetation characters in different habitats of Horqin Sandy Land
2.2 凋落物特征
年际动态分析显示,除2005年的凋落物量大幅度低于其他年份外,2006-2016年变化较小,整体呈逐步增加的趋势(图4-A)。月动态分析显示,科尔沁沙地凋落物量在生长季内整体呈“U”型特征,其中4月的凋落物量最高(203.73 g·m-2),5-7月的量较低且波动较小(11.42~27.20 g·m-2),生长季末的凋落物量再次增加(图4-B)。与植被特征类似,从流动沙丘样地到封育草地,凋落物量具有明显的逐步增加的趋势(图4-C),流动沙丘、固定沙丘和封育草地样地的凋落物量(4-9月累计)分别为8.29,64.33和111.49 g·m-2,且三者间差异显著(F=160.81,P<0.001)。
三因素方差分析表明,科尔沁沙地凋落物量受年、月及样地类型3因素的显著影响,同时各因素间均存在显著的交互作用(P<0.001)(表1)。
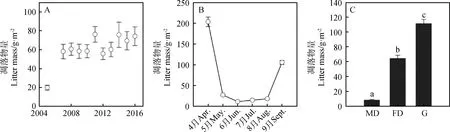
图4 科尔沁沙地凋落物量的年动态、月动态及样地间变化Fig.4 Variations of litter mass dynamics among year,month and habitat in Horqin Sandy Land注:不同小写字母表示差异显著(P<0.001)Note:Different lowercase letters indicated the differences was significant at the 0.001 level

表1 科尔沁沙地不同类型样地、年和月凋落物量的三因素方差分析结果(F值)Table 1 Result from three-way ANOVA (F value) of litter mass among habitats,year and month in Horqin Sandy Land
2.3 植被与凋落物的关系
回归分析显示(图5),科尔沁沙地凋落物(4-9月累计量)与生长季高峰期的植被密度、盖度及地上生物量呈极显著的线性关系,表明植被生产力的高低显著影响科尔沁沙地凋落物量。
各类型样地植被特征与凋落物量(4-9月累计量)的相关分析则显示(表2),植被密度、盖度及地上生物量与凋落物的相关性较小,表明在大区域尺度上,植被生产力与凋落物量密切相关,但在不同类型的小区域尺度上,二者的关系不显著,可能存在其他影响因素。
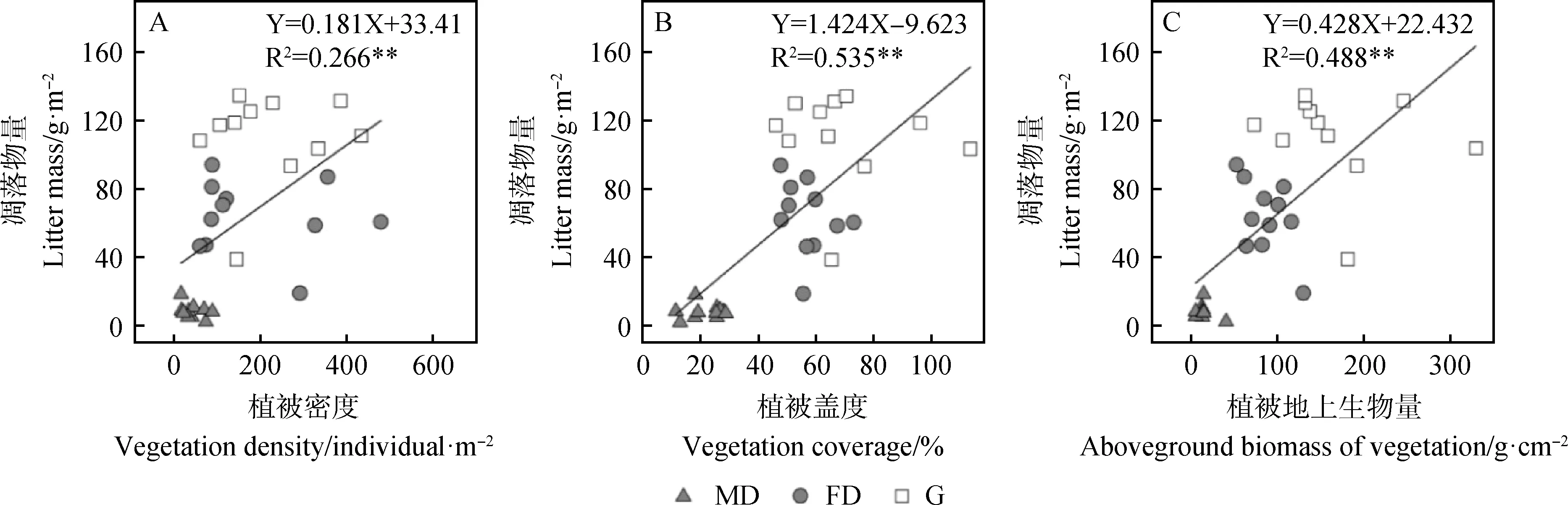
图5 科尔沁沙地植被特征(8月份)与凋落物(4-9月累计)量的回归分析(n=33,2006年凋落物数据缺失)Fig.5 Result of regression analysis between vegetation characters and litter mass at the top of the growing season (August) in Horqin Sandy Land(n=11,and litter mass data in 2006 was missing)
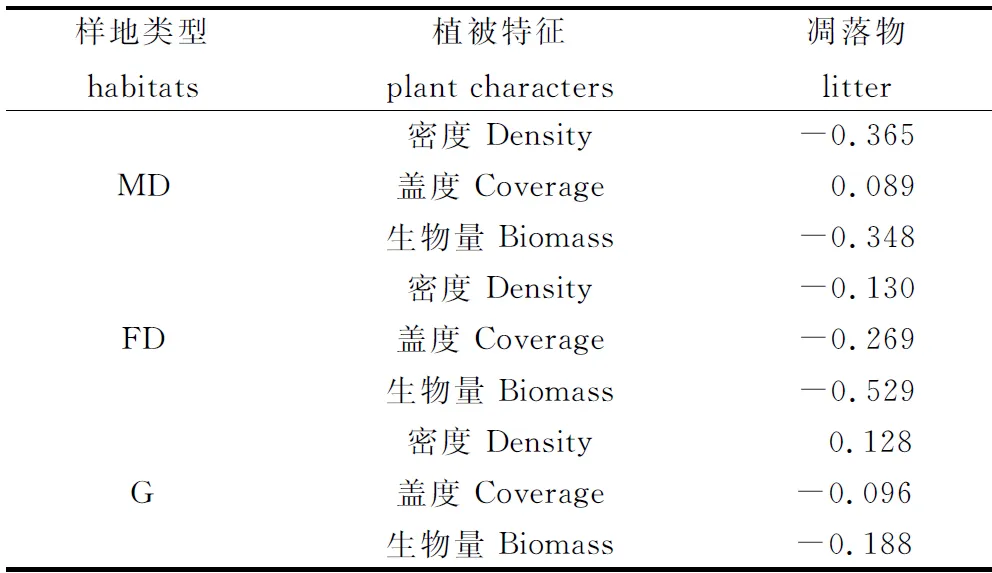
表2 科尔沁沙地植被特征与凋落物量的相关性分析(Pearson指数,n=11,2006年凋落物数据缺失)Table 2 Relationship between plant characters and litter mass in different habitats of Horqin Sandy Land(Data in this table means the Pearson’s correlate index. n = 11,and litter mass data in 2006 was missing)
3 讨论
3.1 植被特征
本研究结果表明,科尔沁沙地植被特征存在极显著的年际变化,这可能与该区的降水波动性有关。已有研究表明,降水是限制该区植被生产力的主要因素[21-22]。该区降水动态显示,2012年总降水量和4-9月降水量分别为430.40 mm和382.20 mm,大幅度高于研究期间的平均降雨量,2005-2016年均降雨量和4-9月降雨量分别为299.08 mm和258.03 mm(数据来源:http://nmd.cern.ac.cn/)。因此,2012年较高的降雨量可能是造成该年植被地上生物量高于其他年份的主要因素。与植被地上生物量相比,植被盖度的波动相对较小,但2012年的盖度最高,可能也是由于较高的降水量所致。另外,植被密度的年际动态与地上生物量和盖度有所不同,最大密度出现在2013年。导致这一现象的原因可能是由于降水的滞后效应,一方面,上一年度相对偏高的降水量可促进植被生产力的提高及种子产量,土壤中种子密度的增加为次年幼苗形成提供保障;另一方面,相对较高的降水量使土壤水分含量提高,存贮于土壤中的水分可为次年植被萌发和幼苗生长提供有利条件[23-24]。土壤水分的滞后效应也可能是次年地上生物量和盖度较高的主要原因[25],2013年全年降水量和4-9月降水量分别为研究期间降水量的88.81%和82.32%,但该年度植被地上生物量和盖度为研究期间平均值的109.72%和109.99%。此外,植被密度与盖度和生物量年际动态的差异可能与物种组成有关。科尔沁沙地的群落组成以一年生短命植物为主,且具有较大的波动性[8],不同年份群落组成差异可能是导致植被密度与盖度和地上生物量年际动态不同步的另一原因。
3.2 凋落物
本研究结果表明,2005年的凋落物量显著低于其他年份。这可能与此前几年连续较小的降雨量有关,2000-2004年连续5年的平均降水量仅为多年平均降水量的74.48%,因此,相对较低的降水量限制了植被的生长,导致凋落物量较低。同样,2012年的凋落物量相对较低,可能是2011年较低的降水量(全年降雨量和4-9月降雨量分别为230.6和215.1 mm)限制了植被生长,导致凋落物量较低。
从月动态来看,4月的凋落物量最大,可能是由于冬季的降雪、冻融作用及较大的风力作用等导致立枯物质转变为地表凋落物[26],9月的凋落物量也高于5-8月,可能由于生长季末期,大量一年生草本枯死和多年生物种器官凋落所致。
3.3 植被特征与凋落物的关系
凋落物来源于植物器官的凋落,因此,植被生长与凋落物的形成具有密切关系[3],植被生产力与凋落物量的关系也在许多研究中得到了验证。如在北京东林山地区,氮添加显著增加了凋落物的产量,其原因是由于氮添加促进了灌木的生长[4]。本研究中,植被地上生物量、盖度和密度与凋落物量呈显著的线性关系,表明在科尔沁沙地,植被生长决定着凋落物的产生。但是,从不同类型样地植被特征与凋落物的相关分析来看,二者不存在显著的正相关关系,表明在小区域尺度上,地表凋落物的形成还受到其他因素的影响。相关研究发现,风力因素是影响科尔沁沙地凋落物分配的重要因素,流动沙丘、半固定沙丘和固定沙丘凋落物运移量均与风速呈显著的二次正相关关系[6]。据此推断,在小生境范围内,风对凋落物的运移作用导致植被特征与凋落物量的相关性不显著。另外,在风力作用下,低植被覆盖区(流动沙丘)一部分杂质可能向高植被覆盖区(封育草地)运移,因此,封育草地凋落物中的杂质可能有一部分来源于低植被覆盖区。但是,本研究中凋落物在不同类型样地间的运移量尚无法确定,后续研究可采用标记法等进行精确定量。
4 结论
综上所述,科尔沁沙地植被特征存在极显著的样地间差异,2005-2016年植被密度、盖度和生物量均具有明显的年际动态,同时年际动态和样地类型间存在显著的交互效应;凋落物量受样地类型、年、月三因素及其交互效应的影响;科尔沁沙地植被特征与凋落物量呈极显著的线性关系,但各类型样地的植被特征与凋落物量的关系不显著。表明植被生长显著影响科尔沁沙地凋落物的产生,但这种影响可能是由于样地类型差异引起,在小区域尺度范围内,凋落物量与植被特征的相关性不显著,凋落物量可能还受其他因素的影响。
猜你喜欢
内蒙古林业(2022年6期)2022-06-27
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
黑龙江工程学院学报(2020年5期)2020-10-21
农家科技下旬刊(2020年1期)2020-03-17
水土保持研究(2019年6期)2019-10-19
草业学报(2019年2期)2019-02-25
现代农业科技(2017年13期)2017-08-09
辽宁林业科技(2017年4期)2017-06-22
