
林下植被清除对不同径级中龄杉木生长速率的影响机制*
2021-01-05 02:47:16陈奕帆付晓莉王辉民戴晓琴陈伏生卜文圣
林业科学 2020年11期
陈奕帆 付晓莉 王辉民 戴晓琴 寇 亮 陈伏生 卜文圣
(1.中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室千烟洲生态站 北京 100101; 2.中国科学院大学资源与环境学院 北京 100049; 3.江西农业大学林学院 南昌 330045)
杉木(Cunninghamialanceolata)是我国南方最重要的常绿针叶树种之一,具有3 000多年的培育历史,在我国亚热带地区广泛分布,具有较好的耐用性和较高的单位产量,是重要的用材林树种(Shietal., 2010)。过去20~30年,杉木人工林多以高密度、短轮伐期经营为主,经营重点集中在中小径材上。随着人们对多样化木材材种需求的升高,大径材供需矛盾凸显(叶功富等, 2006)。因此,研究杉木中龄林的科学经营方式对充分发挥中龄杉木的生长潜力、培育杉木大径材具有重要意义。
林下植被清除是人工林经营管理中的一项重要措施(Lebeletal., 2008),其目的是为了提高木材质量和产量(Stokesetal., 2014)。林下植被清除主要用于幼林抚育阶段,因为在幼林阶段,林下植被与乔木对水分、养分的竞争尤为激烈,清除林下植被对乔木生长最有利(Stokesetal., 2014)。随着乔木的生长,高大乔木本身会阻碍林下植被发展(Busseetal., 1996),清除林下植被对乔木生长的促进作用可能会减弱(Busseetal., 1996; Zhangetal., 2013; Stokesetal., 2014)。但已有研究多关注林下植被清除对林分水平乔木生长的影响(Busseetal., 1996; Takahashietal., 2003; Takahashietal., 2006; Zhangetal., 2013; Stokesetal., 2014),而同一林分中相同林龄不同径级乔木对同一管理措施的响应机制仍不明确。
影响植物个体生长的机制有化学内稳性、资源利用效率、碳分配模式等(Evans, 1989; Jacobetal., 1991; Lietal., 2009; Garrishetal., 2010; Xieetal., 2018; 任书杰等, 2012)。氮(N)、磷(P)是保持植物结构、维持植物生理代谢功能的重要元素,是生产力形成的重要限制因素(Jacobetal., 1991; Lietal., 2009; 任书杰等, 2012)。N、P元素可通过影响CO2同化速率和水分利用效率(WUE)、光合氮素利用率(PNUE),进而影响植物生长(Evans, 1989; Garrishetal., 2010; 李勇, 2011)。N/P既可用来判断林木生长过程中土壤N的供应状态(Herbertetal., 2003),还能反映植物个体、群落及生态系统的养分限制情况(Gusewell, 2004; Zhangetal., 2004)。此外,碳水化合物是树木细胞结构的主要组成成分,也是植物生长代谢过程中的重要能量来源(郑云普等, 2014)。碳水化合物在植物体内的存在形式主要包括结构性碳水化合物和非结构性碳水化合物(NSC)。其中,结构性碳水化合物(木质素、纤维素等)作为植物细胞组成成分主要用于植物生长,而NSC是植物生命代谢的重要碳库,为植物各项生理功能的正常发挥提供物质基础(潘庆民等, 2002)。在植物生长过程中,大部分光合产物不会直接用于新组织形成和代谢过程,而是以NSC的形式储存在植物体内,在植物需要时被消耗以维持植物正常生长和代谢(Kozlowski, 1992)。NSC含量的大小能够反映植物抵御外界逆境的能力(Xieetal., 2018)。在干旱胁迫时,气孔的关闭减少碳固定,使树木依赖储存的NSC进行代谢活动来抵抗逆境和维持渗透压(Mcdowell, 2011)。
本研究以中龄杉木人工林为对象,依托林下植被管理实验平台,研究不同径级中龄杉木化学计量比、碳的光合同化速率、水和氮的利用效率及碳分配模式(如结构性碳与NSC间的分配,NSC在不同器官间的分配),探讨不同径级中龄杉木生长速率对林下植被管理措施的响应机制,为中龄杉木的科学管理策略提供理论支撑。
1 研究区概况
样地位于江西省泰和县中国科学院千烟洲石溪林场(115°03′29.9″E,26°44′29.1″N)。气候属于典型的亚热带大陆性季风气候,年均气温约17.1 ℃,年降水量1 377.4 mm(Yangetal., 2015)。研究区在7-10月存在着明显的季节性干旱(Yangetal., 2017)。土壤为典型低盐基湿润弱育土,土壤母质为红色砂岩和泥石风化物(Kouetal., 2015),pH值为4.87,有机碳、总氮、可溶性有机碳和有效氮含量在自然条件下约为19.1 g·kg-1、1.4 g·kg-1、32.9 mg·kg-1和15.3 mg·kg-1(Yangetal., 2019)。样地坡度约15°,土壤厚度50~100 cm。
2 研究方法
2.1 样地设置
本试验依托2013年建立的杉木人工林林下植被管理实验平台,杉木林于1998年种植。2013年的本底调查显示: 林分平均树高10.2 m,胸径9.8 cm,郁闭度0.75,林下植被以芒萁(Dicranopterisdichotoma)、钩藤(Uncariarhynchophylla)、南烛(Vacciniumbracteatum)、盐肤木(Rhuschinensis)、红花檵木(Loropetalumchinense)、短柄枹(Quercusserrata)、大青(Clerodendrumcyrtophyllum)、杜鹃花(Rhododendronsimsii)为主(Yangetal., 2019),林下植被约为6 236 kg·hm-2。本试验包括2个处理: 保留林下植被(CK),清除林下植被(UR,刈割并移出样地)。每个处理有6块样地重复,样地大小15 m×30 m,样地随机排列。2013年1月移除UR样地林下植被,每个月对样地进行维护,清除UR样地中萌生的草本及灌木。至2016年,UR样地林下植被仅有少量草本及零星灌木。2016年,林分密度为2 143棵·hm-2,平均树高为13.7 m,胸径14.7 cm。2016年,CK处理林木胸径分布: 11~12 cm(4.3%)、12~13 cm(8.5%)、13~14 cm(15.0%)、14~15 cm(25.5%)、 15~16 cm(15.0%)、16~17 cm(10.6%)、17~18 cm(8.5%)、18~19 cm(12.8%); UR处理林木胸径分布: 11~12 cm(16.7%)、12~13 cm(27.1%)、13~14 cm(14.6%)、14~15 cm(0)、15~16 cm(2.1%)、16~17 cm(22.9%)、17~18 cm(12.5%)、18~19 cm(4.2%)。CK处理中胸径为11~12 cm的林木少; UR处理没有胸径为14~15 cm的林木,且15~16 cm及18~19 cm两个范围内的林木少。为确保不同处理间具可比性,即相同径级CK和UR处理都有多个样树重复,所以重点关注12~14 cm和16~18 cm两个径级。其中,12~14 cm和16~18 cm两径级杉木占CK处理总林木数的42.6%,占UR处理总林木数的77.1%。
2.2 样品采集与分析
基于样地内林木径级分布调查,每个样地选8棵涵盖整个径级分布范围的样树,12个样地共计96棵样树。于2016年8月、2018年8月用胸径尺测定每棵样树的胸径,分别记录为D2016、D2018。每棵样树胸径处均有固定树牌以确保数据的连续性和可比性。相对生长速率(RGR)的计算方法: RGR=0.5×(D2018-D2016)/D2016。
于2016年8月(旱季)使用Li-6400便携式光合仪(Li-Cor,Lincoln,Nebraska,USA)测定叶片光合参数。在每棵样树的树冠中下向阳部位选取1个树枝,选择健康且成熟的叶子进行光合参数的测定,以获取每颗样树叶片最大瞬时光合速率(Amax)、蒸腾速率(Tr),并计算WUE(WUE=Amax/Tr)。然后,将树枝、叶片分别按照新(1年)与老(2年、3年)分开,测定各叶龄对应的叶片面积。将枝条和叶片样品烘干,估算叶片比叶面积(SLA)。用生长锥在每棵样树胸高处的树干阴阳两面取约4 cm深的样品,去树皮后混合成1个树干木质部样品,冷藏杀青后65 ℃烘干。将所有烘干后的植物样品用球磨仪粉碎。
采用酶解法测定各样品中的NSC(包括可溶性糖(SS)和淀粉)(Kozlowski, 1992)。为确定林木碳水化合物是优先用于存储还是生长,计算每棵树的NSCtrunk/RGR和NSCleaf/RGR(trunk: 树干,leaf: 树叶)。其中,为确定光合固定的碳在存储与生长间分配的相对优先性,对NSC和RGR的数值均相对各自最大值进行了归一化(Saffelletal., 2014)。NSCtrunk/RGR和NSCleaf/RGR值越大表示光合固定的碳越优先用于存储,而非生长(Saffelletal., 2014)。
叶片全氮采用元素分析仪(德国elementar公司)测定,全磷采用电感耦合等离子体光谱仪(ICP-OES 5300DV,PerkinElmer)测定。用SLA将叶片N质量百分比含量换算成单位叶面积N含量(Narea),然后计算PNUE(PNUE=Amax/Narea)。
2.3 数据处理
利用SPSS18.0,采用双因素方差分析法分析径级和林下植被处理对RGR、Amax、WUE、PNUE、NSC及其组分的影响,采用t检验分析同一径级不同处理间参数的差异性,显著水平为P<0.05。数据在分析前均进行方差同质性检验,以满足正态分布。采用多元逐步回归法分析不同因素(化学计量、资源利用效率、碳分配)对于相对生长速率的影响。利用SigmaPlot12.5软件进行绘图。
3 结果与分析
3.1 林下植被清除对中龄杉木生长的影响
林分水平上,杉木RGR在UR处理为2.54%,CK处理为3.24%,即清除林下植被显著降低了中龄杉木相对生长速率(P=0.035)。清除林下植被对中龄杉木生长的影响因径级不同而异(图1A)。为确保每个径级内均有样树重复,分析中主要关注林下植被管理对12~14 cm和16~18 cm两个径级的中龄杉木生理生态属性的影响。清除林下植被对12~14 cm径级的中龄杉木相对生长速率无显著影响,但会减缓16~18 cm径级的中龄杉木生长(图1B)。
3.2 中龄杉木光合参数和叶片养分的径级差异
清除林下植被对12~14 cm和16~18 cm径级的中龄杉木Amax、WUE、PNUE均无显著影响(表1),这表明清除林下植被没有改变中龄杉木的叶片光合固碳能力、叶片氮素光合效率和水分利用效率。对两径级的中龄杉木叶片全N、全P含量和N/P也无显著影响(表2),表明清除林下植被不会影响中龄杉木的养分限制情况。但两径级的中龄杉木叶片全N、N/P差异显著。16~18 cm、12~14 cm径级的中龄杉木叶片N/P分别为14.00和12.89。

图1 林下植被清除和径级对中龄杉木相对生长速率的影响Fig.1 Effects of understory removal and DBH classes on relative growth rate of middle-aged Chinese fir不同小写字母表示在某一径级不同处理间差异显著(P<0.05)。下同。Different lowercase letters indicated significant differences among treatments in a given DBH class (P<0.05).The same below.
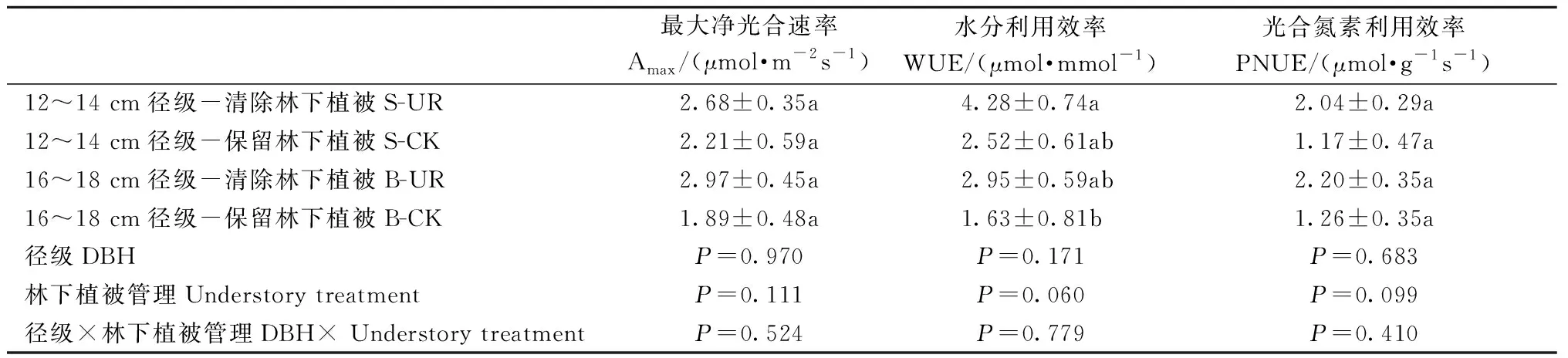
表1 径级和林下植被清除对中龄杉木光合参数的影响①Tab.1 Effects of DBH class and understory removal on photosynthetic parameters of middle-aged Chinese fir
3.3 中龄杉木非结构性碳水化合物分配的径级差异
清除林下植被对12~14 cm和16~18 cm两径级NSCleaf、NSCbranch和NSCtrunk均无显著影响(表3),但对16~18 cm径级中龄杉木NSC在不同年龄叶片中的分配影响显著(图2,3)。对于16~18 cm径级的中龄杉木而言,清除林下植被显著降低NSCnl(图2A)、NSCnl/NSCol(图3A),SSnl(图2B)、SSnl/SSol(图3B)(nl: 新叶,ol: 老叶),而对新叶淀粉无显著影响(P=0.667)。以上结果表明, 清除林下植被主要是通过降低16~18 cm径级的中龄杉木新叶可溶性糖的含量及分配比例,进而降低NSC向新叶的分配。对于12~14 cm径级中龄杉木,清除林下植被显著增加了SSnl/SSol(图3B),而对NSCnl(图2A)、SSnl(图2B)、NSCnl/NSCol无显著影响(图3A)。以上结果表明: 尽管清除林下植被有促进该径级中龄杉木可溶性糖向新叶分配的趋势,但并未造成新叶SS和NSC含量发生显著变化。
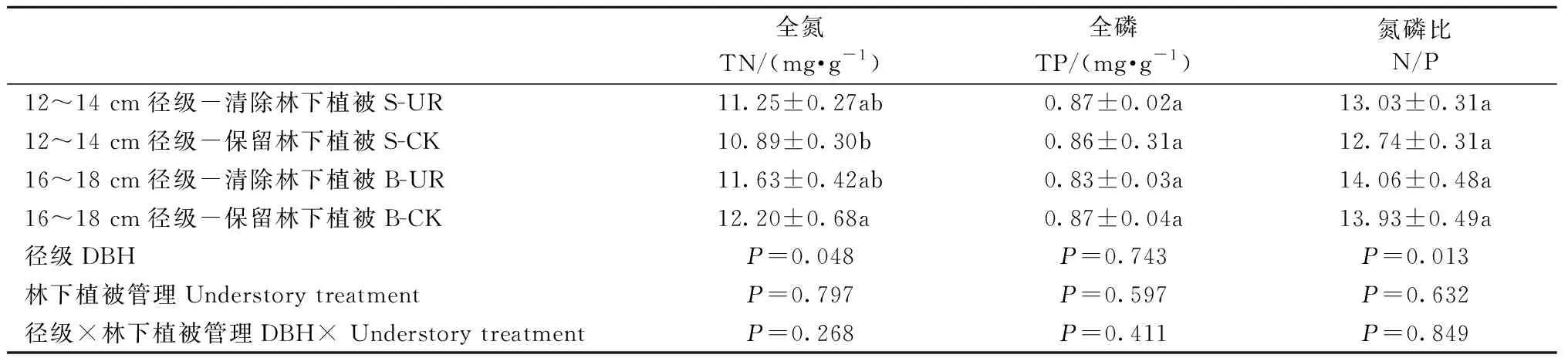
表2 径级和林下植被清除对中龄杉木叶片养分含量的影响Tab.2 Effects of DBH class and understory removal on leaf nutrient of middle-aged Chinese fir

表3 径级和林下植被清除对中龄杉木叶、枝、干非结构性碳水化合物的影响Tab.3 Effects of DBH class and understory removal on NSCs of leaf, branch,trunk of middle-aged Chinese fir
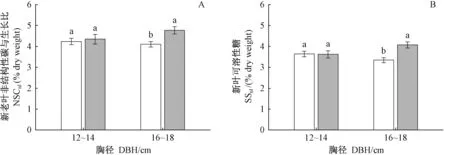
图2 林下植被清除和径级对中龄杉木新叶NSC和可溶性糖的影响Fig.2 Effects of understory removal and DBH classes on NSC and SS in new foliage of middle-aged Chinese fir下角的nl、ol分别为新叶、老叶。The inferior figures nl and ol means newleaf and old leaf respectively.
对于16~18 cm径级的中龄杉木,清除林下植被对NSCtrunk和NSCleaf无显著影响(表3),但减小了其相对生长速率(图1B),导致NSCtrunk/RGR和NSCleaf/RGR比值增大(图4A,4B)。以上结果表明, 清除林下植被后,16~18 cm径级的中龄杉木光合固定的碳优先用于存储,而非生长。清除林下植被对12~14 cm径级的中龄杉木NSCtrunk/RGR和NSCleaf/RGR无显著影响(图4A,4B),说明清除林下植被没有改变12~14 cm径级的中龄杉木光合固定的碳在存储与生长间的分配模式。
3.4 影响中龄杉木生长速率的主控因子
多元逐步回归结果表明: 光合固定的碳在存储与生长间的分配模式是影响中龄杉木生长的主要因素(表4)。NSCleaf/RGR是影响12~14 cm径级的中龄杉木生长速率的主要驱动因子,NSCleaf/RGR越大越不利于杉木生长。对于16~18 cm径级的中龄杉木,NSCleaf/RGR与SSnl共同调控杉木生长速率,NSCleaf/RGR与RGR呈负相关,SSnl与RGR呈正相关,且NSCleaf/RGR对生长速率的解释度高于SSnl。
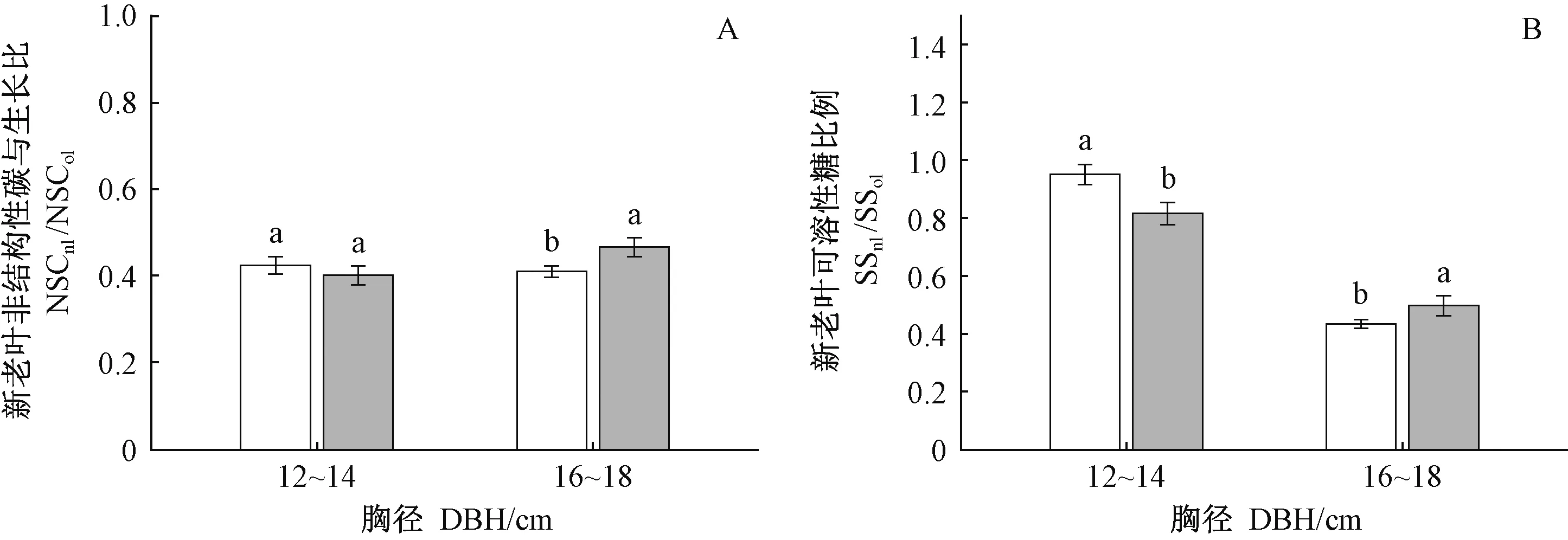
图3 林下植被清除和径级对中龄杉木叶片NSC和可溶性糖在新、老叶间分配的影响Fig.3 Effects of understory removal and DBH classes on allocations of NSC and soluble sugar between the leaves of middle-aged Chinese fir
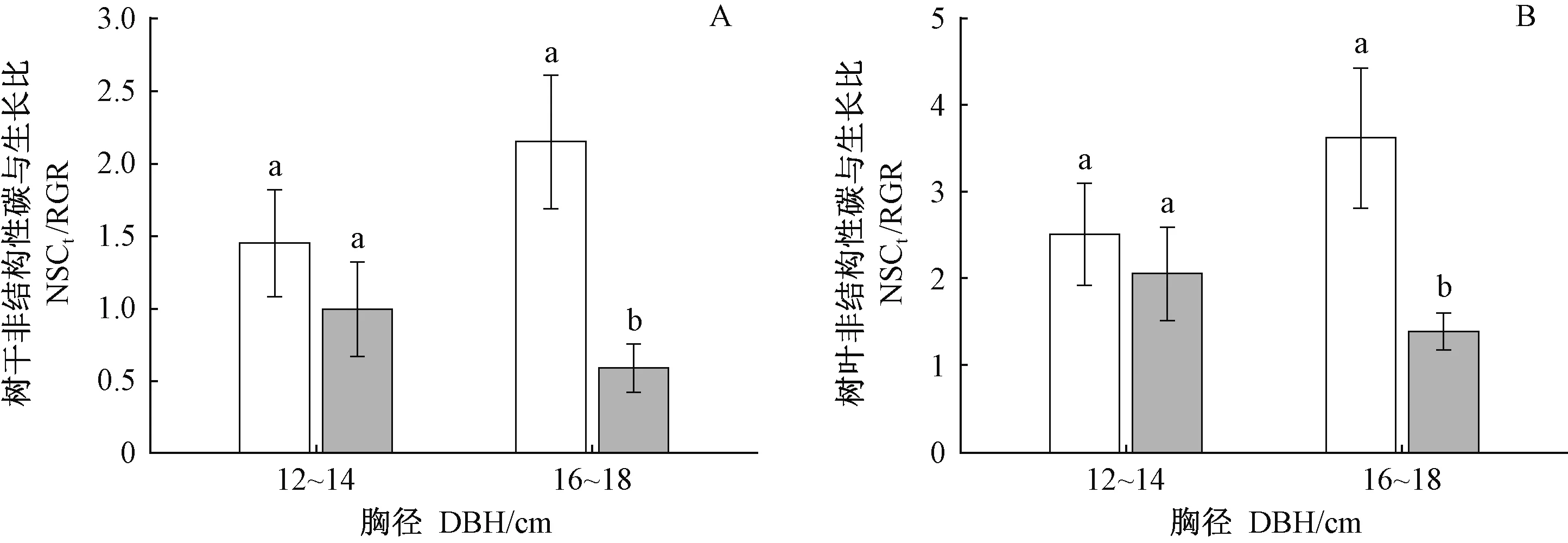
图4 林下植被清除和径级对中龄杉木碳存储与生长关系的影响Fig.4 Effects of understory removal and DBH classes on the relationships between carbon storage and growth of middle-aged Chinese fir
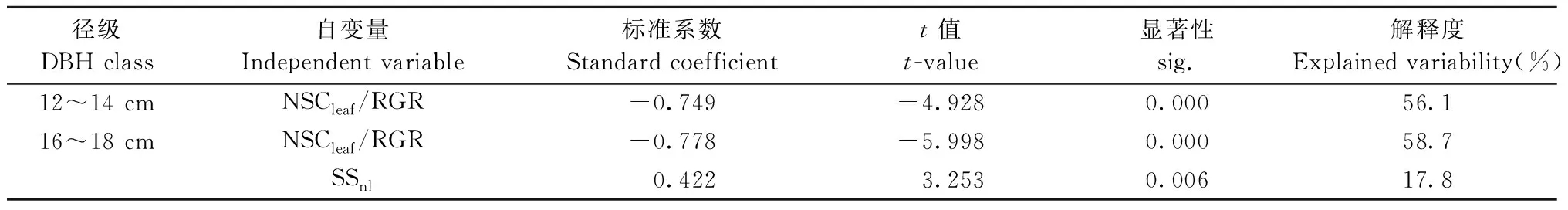
表4 中龄杉木相对生长速率多元逐步回归结果Tab.4 Results of stepwise regression analysis for relative growth rate of middle-aged Chinese fir
4 讨论
4.1 林下植被清除对林分水平乔木生长的影响
当林下植被与乔木竞争激烈土壤水分、养分时,清除林下植被一般可促进保留木生长(Takahashietal., 2003; Takahashietal., 2006; Lebeletal., 2008; Zhangetal., 2013; Stokesetal., 2014)。但本研究结果与此相反,Wan等(2014)研究4年生桉树(Eucalyptusrobusta)林中清除林下植被减慢了乔木生长(Wanetal., 2014)与本研究相似。这说明清除林下植被对乔木生长的影响存在不确定性,可能是因林下植被与乔木间的竞争会随乔木生长而相对减弱(Busseetal., 1996; Zhangetal., 2013),或使二者关系主要表现为协同关系。
首先,林下植被覆盖能有效减少土壤侵蚀(袁正科等, 2000),林下植被也可将渗入土壤深层的养分运移到表层土壤以供其他植物利用(Jobbágyetal., 2004)。其次,林下植被可缓解土壤酸化、优化土壤微生物群落结构、提高真菌比例(Fuetal., 2015)。此外,土壤碳分布与林下植被细根密切相关,林下植被细根多的土壤斑块的碳含量高(Liaoetal., 2014),且土壤表层中林下植被吸收根的量高于乔木(Jiangetal., 2018),林下植被吸收根分泌物能提高土壤微生物活性、加快凋落物分解(郑琳琳等, 2017),因此林下植被细根对促进养分释放有重要意义。清除林下植被会减少根系分泌物,导致β-1,4-葡糖苷酶活性及其最大反应速率降低,碳获取酶活性下降(Yangetal., 2018; Yangetal., 2019)。因此,清除杉木人工林林下植被可显著降低土壤全N、全P及可溶性C含量、减小土壤真菌含量、减缓土壤养分循环速率(Wuetal., 2011; Liuetal., 2012; Zhaoetal., 2012)、使植物光合固定的碳优先用于地下根系生长而非地上生长(Reichetal., 2014)。
清除林下植被会使太阳辐射能、蒸散发量、地表空气流动等环境因子进行系统内重新分配(吴彦等, 2004; 臧润国, 2016),土壤温度及水分分布特征发生变化(Hoggetal., 1991; Takahashietal., 2003; Matsushimaetal., 2007; Wuetal., 2011),间接影响表层土壤有机质的分解及氮矿化过程。研究发现, 清除林下植被可使10 cm处土壤温度升高(Hoggetal., 1991; Matsushimaetal., 2007; Wuetal., 2011)、提高土壤微生物活性和凋落物分解速率、加快N的矿化与硝化、提高N有效性、促进乔木生长(杜忠等, 2016)。而同在本试验样地开展的研究发现,林下植被清除后土壤温度、水分均无显著变化(杨洋等, 2016)。因此,土壤温度、水分不是本研究中乔木生长减缓的原因。
4.2 林下植被清除对林木个体化学计量、资源获取及利用效率的影响
不同林下植被物种的根际效应不同(Daietal., 2018; 莫雪丽等, 2018),且不同乔木个体周围的林下植被物种可能不同。因此,清除林下植被对不同乔木个体周围土壤养分释放的影响可能会有差异。但本研究中的林木样树均是随机取样,可排除这个潜在影响因素。
氮和磷在植物的生理功能中起着至关重要的作用,是陆地生态系统中限制植物生长的最重要的元素(Chapin, 1980; Reichetal., 1997)。叶片N/P可表征植物养分受限情况。通常,叶片N/P>16表示植物受P限制; 叶片N/P<14表示植物受N限制; 叶片N/P处于14和16之间表示N、P均不受限制或者均受限制(Koerselmanetal., 1996)。本研究中12~14 cm径级的中龄杉木受N限制,而16~18 cm径级的中龄杉木N、P化学计量平衡较好。但林下植被清除并未影响两径级的中龄杉木的养分状况(表2)。这可能是因为清除林下植被一方面减少了单位土体养分释放的总量,另一方面排除了林下植被对土壤养分消耗这一分量(即清除林下植被后土体养分仅供乔木所用)。
在一个植物群落中,长势好的植物个体通常会有较高的资源获取能力或资源利用效率,如碳同化速率较大、水分利用效率或氮利用效率高等(Funketal., 2007; 刘鹏等, 2019)。尽管清除林下植被可增大乔木冠层叶片的最大净光合速率(Kitaoetal., 2003; Kobayashietal., 2006; Sirdah, 2017),减小林分水分利用效率(张浩, 2017)。但本研究中林下植被清除未改变林木的资源吸收和利用效率(表1)。
4.3 林下植被清除对林木个体碳分配的影响
碳水化合物主要以可溶性糖的形式在韧皮部运输,参与新陈代谢,为树木生长提供重要能源(Chapin, 1990)。树木遇到干旱等逆境胁迫时,用可溶性糖调节渗透压,以维持树木细胞的正常活性(Salaetal., 2012)。新叶的CO2光合能力、净同化量与净光合强度高于老叶,对林木的碳固定及生长意义重大(张小全等, 2000; 谢国阳, 2002; 许忠坤等, 2008)。清除林下植被后,16~18 cm径级树木新叶的NSC和可溶性糖含量均下降,可导致干旱季节杉木叶片渗透压调节能力下降,进而影响杉木生长。其次,林下植被清除后,16~18 cm径级的中龄杉木相对提高了树干、叶片NSC存储量,进而相对地减少了用于生长的结构性碳的分配(图4)。即当光合吸收的碳不变时,16~18 cm径级中龄杉木光合同化的碳优先用于存储,而非生长。综上可知,对于16~18 cm径级的中龄杉木而言,清除林下植被后,叶片(尤其是新叶)NSC及可溶性糖的降低及光合固定的碳优先用于存储而非生长共同导致了其生长缓慢(表4)。
对于12~14 cm径级的中龄杉木,叶片SSnl/SSol显著增加(图3)。这可能是由于该径级的中龄杉木受N限制显著,而植物面对资源不足时将固定的碳优先向获取资源能力强的器官或组织中分配(Chapinetal., 1987)。然而,限制12~14 cm径级的中龄杉木生长的主要因子是NSCleaf/RGR(碳存储与生长间的关系),而非叶片NSC(或可溶性糖)在新、老叶之间的分配比例(表4),且林下植被清除对该径级中龄杉木NSCleaf/RGR无显著影响。因此,林下植被清除没有显著改变12~14 cm径级的中龄杉木的生长速率。
4.4 本研究的局限与不足
本研究是用2016年杉木生理生态指标来解释其后续2年的生长,这存在一定的不确定性,因为随着林下植被管理措施的年限延长,林木生长的响应规律可能会发生变化。因此,林木生长对林下植被管理措施的长期响应规律还需未来更多研究进行验证。
此外,林业生产中通常是割倒林下植被后将残余物归还到林分中。林下植被割后归还比林下植被正常生长更有利于增加土壤酶活性,因为产生的林下植被地上、地下植物残体比活根和正常凋落物输入提供的碳源更多(杨洋等, 2016)。而本研究是持续地将林下植被割倒并将产生的残余物移出到林分外,这可能导致水土流失、不利于土壤水分和微生物群落结构等土壤微环境的维持(Zhaoetal., 2013)。因此,将林下植被残余物移出到林分外也可能导致林下植被去除未促进林木生长。未来研究需增加割倒林下植被但不移出的处理,并设计不同的林下植被刈割强度梯度,进一步探讨林下植被管理对林木生长的影响机制。
5 结论
基于本研究的观测数据,表明清除并移除林下植被会减缓中龄杉木的生长速率。但不同径级的中龄杉木生长速率对林下植被管理措施的响应不同,即林下植被清除会降低16~18 cm径级的中龄杉木的相对生长速率,而对12~14 cm径级的中龄杉木相对生长速率无显著影响。林下植被管理对中龄杉木生长影响的径级依赖性主要受碳分配(不同形态间,相同形态不同器官间)机制调控,而非化学计量、资源获取及利用效率等机制影响。清除林下植被导致16~18 cm径级的中龄杉木新叶NSC(尤其是可溶性糖)含量降低,且使林木光合产物优先用于存储而非生长,进而减缓了该径级的林木生长速率。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
农民致富之友(2020年8期)2020-05-11 05:54:38
现代园艺(2017年23期)2018-01-18 06:58:24
现代园艺(2017年23期)2018-01-18 06:58:19
广西林业科学(2016年2期)2016-03-20 05:53:20
