
光质对琼枝生长和生理特性的影响
2020-12-28 03:15童立豪吴翔宇曾俊邢诒炫顾志峰石耀华唐贤明
中国渔业质量与标准 2020年6期
童立豪,吴翔宇,曾俊,邢诒炫,顾志峰,石耀华,唐贤明*
(1.海南省海洋与渔业科学院,海南省热带海水养殖技术重点实验室,海口 571126; 2.广西科学院广西红树林研究中心,广西红树林保护和利用重点实验室,广西 北海 536007; 3.海南大学海洋学院,海口 570228)
光是植物进行光合作用的原动力,光质是光的重要属性[1]。光质对藻类的生长发育、形态建成和光合作用等具有调控作用,同时,藻体也会根据光质的变化而产生不同的适应机制[2]。人工光源常用于代替或补充自然光源的不足,已经成为控制植物生长发育与作物栽培环境的重要手段[3]。LED光源是目前最新的节能冷光源[4],与传统光源相比具有亮度高、能耗低、寿命长、体积小、无辐射、无污染及耐受恶劣环境条件等优势[5],其在藻类研究中的应用已成为热点。其次,LED纯色光源的发射光谱波长可控且带宽窄,与藻类光合器官所吸收的光谱匹配度高,通过合理优化可达到精准调控藻类形态发育与新陈代谢、提升产量的目的[6]。在过去的研究中,常通过彩色薄膜和滤光片来改变入射光以研究光质对海藻生理的影响[7],然而这些方法具有带宽大,费用高的缺点。
琼枝(Betaphycusgelatinae),隶属于红藻门、真红藻纲、杉藻目、红翎菜科、琼枝藻属(Betaphycus),是热带、亚热带性海藻,分布在中国、菲律宾、日本和印度尼西亚等地,在中国分布于海南岛、东沙群岛和台湾岛等热带海区[8],是海南重要的热带资源之一。琼枝在生态和经济上具有较高的价值,藻体在生长过程中可提供固碳释氧、吸收营养盐及调节生物多样性等生态系统服务功能。藻体中的卡拉胶被广泛应用于食品、日化、医药制造等产业[9]。同时,琼枝富含生物活性物质,具有广谱性病毒抑制活性[10-11]、抗凝血和抗血栓活性[10, 12-13]、抗真菌活性[14]、抗炎症和抗肿瘤活性[15]等作用。
目前,海南的琼枝供不应求,加之人类活动与全球气候变化给近岸生态系统造成持续的压力[16],如污染物、营养物流入沿岸水域,使得海水透明度降低[17],直接限制了海藻的光合作用,进一步加剧琼枝市场供需不平衡的现象。人工养殖琼枝是解决该矛盾的有效途径,利用LED光源能够满足藻体对不同光质的需求,增加产量。不同种大型海藻对光质的需求存在差异,如Kim等[18]发现,红光、绿光下生长的红藻龙须菜(Gracilariatikvahiae)具有较高的生长速率;Franklin等[19]与Barufi等[20]发现,蓝光可增加红藻鹿角菜(Chondruscrispus)与江蓠(Gracilariabirdiae)的藻红蛋白与藻蓝蛋白含量。关于琼枝的研究多集中在光照强度[21]、温度[22]和盐度[23]等方面,尚未有关于光质对其生长发育的研究报道。本研究以琼枝为实验材料,研究其在不同光质条件下的生长、形态、颜色和光合色素的差异性,可为科学种养琼枝,提高产量提供参考依据。
1 材料与方法
1.1 实验材料
琼枝取自海南省海洋与渔业科学院琼海科研基地(110°40′07.2″E, 19°21′58.2″N),挑选颜色正常、生长旺盛的藻体洗净,在自然海水中暂养7 d后开始实验。暂养温度28 ℃,盐度28,光照系统由传统的荧光灯提供,光照强度72 μmol/(m2·s),光周期为光亮(L)∶黑暗(D)=12 h∶12 h。
1.2 实验设置
挑选健康无溃烂的琼枝2~3 g(鲜质量)于1 L玻璃烧杯中培养。设置红、黄、蓝、绿、紫和白,共6个不同光质,每个处理设置3个重复;采用多排密集LED灯管(型号为81331,中国浦田公司,中山)提供不同光质,通过距离远近调节光照强度,使光照强度统一维持在72 μmol/(m2·s)。温度、盐度和光周期与暂养条件相同。每7 d更换1次海水并称重,实验持续35 d。
1.3 研究方法
1.3.1 生长速率和增重率计算
每7 d测量琼枝的鲜质量,称量前用吸水纸快速吸干藻体表层水。根据下列公式计算增重率(weight gain rate,WGR)和相对生长速率(relative growth rate,RGR):
WGR(%)=(Mt-M0)/M0×100
式(1)
RGR(%·d-1)=(lnMt-lnM0)/t×100
式(2)
式中:t为培养时间(d),M0为初始鲜重(g),Mt为t天后的鲜重(g)。
1.3.2 形态性状的测定
实验前,将生长状态一致的实验藻体进行随机分组,实验35 d后,计算各处理组的主枝上的分枝数和齿芽数,使用游标卡尺测量各样本最长的3个分枝的长度和宽度。
1.3.3 颜色参数的测量
实验第35 d,用相机(佳能EOS6D,日本)对藻体进行无差别图片采集,后将待测图片导入CSE-1成像色度检测分析系统中,随机选取3个点,获得相应的颜色参数Lab(CIE 1976)和三刺激值XYZ(CIE 1931)(L为明度,数值介于0~100之间;a指颜色的红绿特征,“+a”为红,“-a”为绿;b指黄蓝特征,“+b”为黄,“-b”为蓝;X、Y、Z分别指红、绿、蓝三原色值,表示颜色的光色度特性)。计算各处理组L、a、b、X、Y和Z值的平均值,分别作为该藻体颜色参数和三刺激值。
1.3.4 色素和藻胆蛋白的提取和分析
实验第35 d提取和分析藻体色素和藻胆蛋白含量。
叶绿素a(Chla)和类胡萝卜素(Car)质量分数的测定采用无水甲醇提取法。根据Porra[24]的公式计算叶绿素a的含量;根据Parsons和Strickland[25]的公式计算类胡萝卜素的含量。
藻红蛋白(PE)和藻蓝蛋白(PC)质量分数的测定采用磷酸缓冲液提取法。根据Beer和Eshel[26]的公式计算藻红蛋白和藻蓝蛋白的含量。
1.4 统计分析
采用Excel 2019和SPSS 18.0统计软件进行数据处理及统计分析。用One-way ANOVA(Turkey)和t-检验进行显著性检验(P<0.05)。
2 结果与分析
2.1 不同光质下琼枝的生长情况
琼枝在紫光下的生长情况优于其他光质,35 d的RGR为(1.22±0.05)%/d,WGR为(44.60±3.43)%;其次为蓝光,在该光质下,RGR和WGR分别为(1.19±0.11)%/d和(42.14±5.45)%;绿光条件下琼枝无明显生长,RGR仅为(0.66±0.03)%/d。各光质下,琼枝的RGR依次为紫光>蓝光>白光>红光>黄光>绿光,其中,紫光、蓝光、白光和红光无显著差异(P>0.05),黄光和绿光无显著差异(P>0.05)。从WGR分析,紫光下藻体的WGR分别是绿光、黄光、红光、白光和蓝光下藻体WGR的2.12倍、1.83倍、1.22倍、1.18倍和1.05倍(表1)。
紫光、蓝光、白光和红光的WGR在35 d的实验期内,总体趋势基本一致,紫光WGR从一开始就明显高于其他光质。在前14 d各处理组的增重情况均较为缓慢,从第14 d到28 d,紫光、蓝光、白光和红光的WGR呈线性增长,28 d后开始放缓;而黄光和绿光始终保持缓慢增长的趋势(图1)。
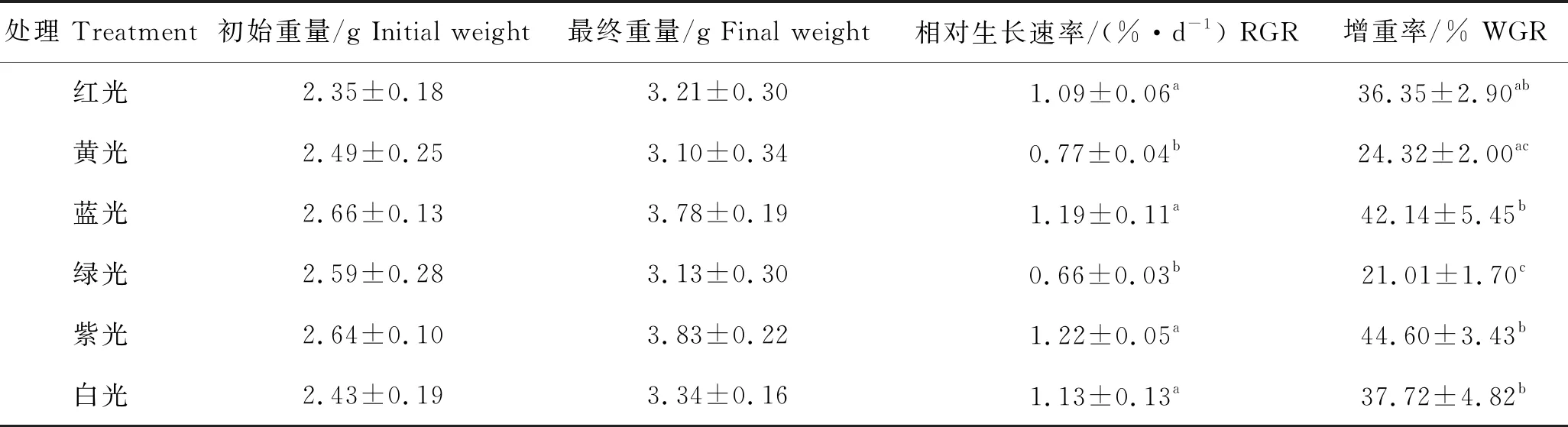
表1 不同光质下琼枝的生长变化Tab.1 Growth changes of B. gelatinae under different light qualities

图1 不同光质下琼枝增重率随时间的变化Fig.1 WGR changes of B. gelatinae with time under different light qualities
2.2 不同光质下琼枝的外部形态、颜色特征
经过35 d的实验,不同光质条件下琼枝的外部性状差异明显(表2)。藻体的分枝数,以蓝光下的分枝数最多,为(8.33±0.47)个;其次为白光和红光;绿光的分枝数最少,仅为(2.33±0.47)个;而黄光、紫光下的分枝数与绿光相近,且无显著差异(P>0.05)。齿芽数最多的为蓝光和白光,二者差异不显著(P>0.05);其次为红光和黄光,二者差异不显著(P>0.05);最少的为紫光和绿光,二者差异不显著(P>0.05)。紫光下的藻体分枝长度最长,且宽度最宽,分别为(57.13±4.51)mm和(6.70±0.21)mm;其次为蓝光,分枝长度和宽度分别为(48.23±2.92)mm和(5.83±0.25)mm。

表2 不同光质下琼枝的外部性状Tab.2 External characters of B. gelatinae under different light qualities
对不同光质处理的藻体颜色差异进行参数分析(表3)。蓝光下藻体的三刺激值XYZ最大,且与各组均存在显著差异(P<0.05)。明度L也以蓝光最大(45.69±4.78),与各组差异显著(P<0.05)。各光质下藻体的红绿特征a均为正值,偏红色,其中以蓝光和紫光下的红绿特征a最小,颜色为淡红色,其他光质的藻体颜色均为深红色。在黄蓝特征b上,各光质下的显著性差异并不明确,均为正值,特征偏黄。

表3 不同光质下琼枝的颜色参数Tab.3 Color parameters of B. gelatinae under different light qualities
2.3 不同光质下琼枝的色素和藻胆蛋白含量
藻体中的叶绿素a在黄光、绿光和白光处理下的含量显著高于蓝光和紫光处理(P<0.05),含量最低处理组为蓝光,仅59.65 μg/g。藻体中的类胡萝卜素含量总体偏低,在17.55~29.82 μg/g的范围内,但黄光、绿光和白光下的类胡萝卜素含量依然显著高于蓝光和紫光(P<0.05)(图2)。

图2 不同光质下琼枝的色素含量不同字母表示存在显著性差异(P<0.05)。下同。Fig.2 Pigment contents in B. gelatinae under different light qualitiesDifferent superscript letters means that there are significant differences for the data (P<0.05). The same below.
蓝光和紫光下,藻体中的藻红蛋白含量显著低于黄光、绿光和白光下的藻红蛋白含量(P<0.05),与叶绿素a、类胡萝卜素含量的显著性相一致。而藻蓝蛋白的含量偏低,各光质下的含量相近,除了紫光和白光存在显著性差异外(P<0.05),各光质下的含量均无显著性差异(P>0.05)(图3)。

图3 不同光质下琼枝的藻胆蛋白含量Fig.3 Phycobiliprotein contents in B. gelatinae under different light qualities
3 讨论
3.1 光质对琼枝生长的影响
光是藻类生长发育最重要的生态因子,不仅是光合作用的能源物质,还可调控藻体的生长发育过程。光质作为光的重要属性,对藻类的生长发育、形态建成和光合作用等具有重要的调控作用。有关光质对大型海藻生长和生理的影响已有报道,但其结果因种类不同而存在差异。在对坛紫菜(Pyropiahaitanensis)自由丝状体[27]、龙须菜(Gracilarialemaneiformis)和铜藻(Sargassumhorneri)[3]的研究中均发现蓝光处理后的藻体具有较高的生长速率。同时,也有研究表明角叉菜(Chondrusocellatus)[28]、脐形紫菜(Porphyraumbilicalis)[29]和江蓠[30]在红光下有较高的生长速率,红藻龙须菜在红光和绿光的照射下生长速率都较高[18]。本研究中,不同光质对琼枝的生长影响较为明显,紫光和蓝光的照射有利于琼枝的生长,其生长速率和增重率都较其他光质大,绿光和黄光处理则生长较为缓慢。造成本研究与其他研究的差异,可能是因为研究对象不同,不同种海藻在长期自然生存中,受不同生长环境的影响,形成了不同的光适应机制,对不同光质的利用存在差异所致。此外,不同的实验光源和实验方法也可能对实验结果产生一定的影响。
3.2 光质对琼枝形态、颜色建成的影响
光质在藻类的整个生长过程中起着重要的调控作用,光色的差异不仅会影响藻体的生长过程,不同光色照射下藻体形态亦会发生变化。在海带的研究中,海带的柄和固着器的延伸率在红光下显著高于蓝光[31],蓝光可以促进海带幼苗长度和宽度的增加[32]。本研究发现,蓝光和白光有利于琼枝分枝和齿芽的形成,紫光能明显增加分枝的长度和宽度,其次为蓝光。综上,蓝光既有利于琼枝的分枝、发芽又能够促进藻体分枝的生长。
藻类的颜色主要是基于其捕光色素复合体含有的光合色素颜色[3],通过对颜色的分析判断能够更好的了解藻体的生长状态,而利用CSE-1成像色度检测分析系统可以对颜色进行深入的分析,使颜色实现数据化和形象化。在藻体颜色参数Lab的分析中,蓝光照射的藻体明度L较大,红绿特征a则相对其他光质处理的藻体而偏向于绿原色,在黄蓝特征b中偏向于蓝原色,与其他光质处理的藻体,存在明显的差异。分析形态、颜色上的差异性变化,可能是藻类为适应不同光质,满足自身生长的一种适应机制,也可能是在不同光质下,藻体中基因表达、酶活变化的一些调控。
3.3 光质对琼枝光合色素的影响
研究证明,不同光质既影响藻类的生长、形态特征,也影响着藻体中光合色素含量和组成比例[30, 33]。在对石莼(Ulvalactuca)的研究中发现红光能够提高类胡萝卜素的含量[3],而蓝光较红光更利于红藻紫菜(Porphyraleucosticta)中藻胆蛋白的合成[34],同时蓝光下条斑紫菜(Porphyrayezoensis)的藻胆蛋白含量高于其他光质下的含量。本研究中,琼枝在绿光和黄光下的生长速率显著低于紫光和蓝光,但其叶绿素a、类胡萝卜素和藻红蛋白的含量却显著高于紫光和蓝光,说明绿光和黄光下有利于光合色素的积累。这种差异的出现,可能是由于不同藻类的光合色素系统对不同光质的吸收和利用存在差异,藻体根据自身生长需要而对光合色素的含量和比例进行相应的调整。琼枝光合色素系统对绿光和黄光的吸收利用有限,光能利用效率低,不能满足藻体正常的光合需要,为了使藻体能够正常生长,藻体需要合成更多的光合色素来进行光合生长。
4 结论
本研究通过对不同光质条件下琼枝的生长和生理特征进行研究,发现不同光质对琼枝生长、生理指标的影响差异显著。紫光和蓝光的照射有利于琼枝生长速率的提高;蓝光有助于琼枝藻体发芽和分枝的形成,紫光条件下琼枝藻体的分枝易于伸长和长粗;叶绿素a、类胡萝卜素和藻红蛋白易于在黄光、绿光和白光条件下的藻体中积累。因此,为了更好的促进琼枝的养殖生长,在对琼枝光质需求的调节上,可以考虑在养殖初期照射蓝光,促进琼枝的发芽和分枝的形成;在养殖中后期照射紫光,促进琼枝藻体分枝进一步伸长、长粗,提高生长速率。本研究可为深入研究琼枝的生物学性状和人工栽培技术提供有益指导,促进琼枝的科学种养。
猜你喜欢
上海农业学报(2022年1期)2022-03-08
Plasma Science and Technology(2021年8期)2021-08-05
中国美容医学(2021年5期)2021-06-22
中国中药杂志(2016年21期)2017-02-16
中国中药杂志(2016年21期)2017-02-16
东方教育(2016年14期)2017-01-16
作文与考试·小学低年级版(2016年10期)2016-09-10
意林·少年版(2016年11期)2016-09-10
祝你幸福·知心(2016年3期)2016-03-29
安徽文学(2009年7期)2009-08-11
