
产葡萄糖异构酶菌株的筛选及酶学性质研究
2020-12-22 08:55:02廖贵芹程国军吴和涛滕雪刘莹
中南民族大学学报(自然科学版) 2020年6期
廖贵芹, 程国军, 吴和涛, 滕雪, 刘莹
(1华中科技大学 生命科学与技术学院,武汉 430074;2中南民族大学 生命科学学院,武汉 430074)
葡萄糖异构酶(GIase)能将D-葡萄糖、D-木糖等转化为D-果糖、D-木酮糖等相应酮糖[1].它是工业上从淀粉大规模制备高果糖浆(HFCS)的关键酶[2-4],其酶活的高低直接关系到高果糖浆的产量和生产成本.
目前国产葡萄糖异构酶主要问题是酶学性质不理想,即酶活低,热稳定性差、耐受金属离子的能力差.葡萄糖异构酶大多数来自于细菌、放线菌,小部分来自真菌[5-6].因此,筛选和构建高生产性能的商业用葡萄糖异构酶一直是研究的热点.为获得高产葡萄糖异构酶的优良菌种,本文从土壤中分离得到一株产葡萄糖异构酶活比较高的细菌菌株,并对其部分酶学性质进行了研究,为后期优化产葡萄糖异构酶的条件奠定了理论基础.
1 材料与方法
1.1 材料和仪器
本文中21份土样采自湖北省等邻近省份,来源于含水量较低、有机质丰富的土壤[7].可见分光光度计(V-1000,上海翱艺仪器有限公司);超声波细胞粉碎机(JY92-IIN型,宁波新芝生物科技有限公司,破碎条件:变幅杆Φ3,功率为40%, 工作时间4 s,停30 s,共238 s);测序(北京三博远志公司).
1.2 培养基
(1)细菌初筛培养基:D-木糖5 g·L-1,硫酸铵2 g·L-1,柠檬酸钠1 g·L-1,MgSO4·7H2O 0.2 g·L-1,K2HPO44 g·L-1,KH2PO46 g·L-1,琼脂粉20 g·L-1,pH 7.2,115 ℃灭菌20 min[8].
(2)细菌种子培养基:牛肉膏3 g·L-1,蛋白胨10 g·L-1, NaCl 10 g·L-1, pH 7.2,121 ℃灭菌20 min.
(3)细菌发酵培养基:D-木糖2 g·L-1,蛋白胨5 g·L-1,酵母膏2 g·L-1,MgSO4·7H2O 1 g·L-1,NaCl 10 g·L-1, K2HPO41 g·L-1,pH 7.2,115 ℃灭菌20 min.
(4)放线菌初筛培养基:可溶性淀粉20 g·L-1,KNO31 g·L-1,MgSO4·7H2O 0.5 g·L-1,NaCl 0.5 g·L-1, K2HPO40.5 g·L-1,FeSO40.01 g·L-1,K2Cr2O70.03 g·L-1,琼脂粉20 g·L-1,氨苄青霉素2 mg·L-1,121 ℃灭菌20 min.
(5)放线菌种子培养基:蛋白胨5 g·L-1,葡萄糖10 g·L-1,NaCl 5 g·L-1,pH 7.2~7.4,115 ℃灭菌15 min.
(6)放线菌液体发酵培养基:玉米浆20 g·L-1,玉米粉10 g·L-1,MgSO4·7H2O 1 g·L-1,K2HPO41 g·L-1,麸皮水解液配制,115 ℃灭菌30 min.
1.3 方法
1.3.1 菌株的筛选
土样进行梯度稀释,采用涂布平板法,接种在细菌初筛培养基平板上,对单菌落采用平板划线分离法进行纯化[9-10].将纯化的菌株逐个用细菌种子培养基培养,培养过夜,再按约10%的接种量(所有菌种保证菌液浓度一致,可按需适当稀释)接种至细菌发酵培养基中,进行摇瓶液体发酵培养24 h.接着用超声波破壁法制备胞内、胞外初酶液,采用改进的硫酸咔唑法测定每个菌株的胞内、胞外初酶液的活性大小,最终选出一个产酶活性最大的细菌菌株.
筛选放线菌菌株时与细菌的操作相似,土样梯度稀释液涂布在放线菌初筛培养基平板上,纯化分离.再将菌株逐个接种至放线菌初筛培养基的试管斜面,培养至产孢稳定期,接着制备浓度为107cfu·mL-1的孢子悬液,按1%接种量接种到放线菌种子培养基,培养48 h[7].将种子液按10%的接种量转移至放线菌液体发酵培养基,摇瓶培养48 h.与细菌相同,制备胞内、胞外初酶液,通过改进的硫酸咔唑法测定酶活,选出一个产酶活性最大的放线菌菌株.
1.3.2 粗酶液的制备
发酵液于4000 r·min-1,10 min,4 ℃离心,收集上清即为胞外粗酶液.沉淀用三乙醇胺-盐酸缓冲液(pH 8.0)-10 mmol·L-1MgSO4洗涤2次,再加入10倍沉淀体积的缓冲液-10 mmol·L-1MgSO4重悬菌体,超声破碎.12000 r·min-1,20 min,4 ℃离心收集上清即为胞内粗酶液[11].
1.3.3 酶活的测定
测定酶活采用改进的硫酸咔唑法[11].250 μL 0.6 mol·L-1D-葡萄糖中加入经适当稀释的100 μL粗酶液、200 μL 0.025 mol·L-1三乙醇胺一盐酸缓冲液,在70 ℃反应15 min,再加入50%三氯乙酸50 μL终止反应;立即加入3 mL体积分数为70%的硫酸(冰浴冷却)、100 μL质量浓度为2.4 %的半胱氨酸-盐酸盐、100 μL质量浓度为0.12%的乙醇-咔唑溶液.混匀后60 ℃反应30 min,取出用流水冷却;在可见分光光度计上,使用波长为560 nm,以试剂空白为参比,光程为1 cm石英杯测其吸收度.
分别吸取50 μg·mL-1的果糖标准溶液0、0.1、0.2、0.3、0.4、0.5、0.6 mL,按上实验方法依次加入硫酸、半胱氨酸盐酸盐、乙醇咔唑溶液,60 ℃显色30 min,测定吸收度,绘制工作曲线.葡萄糖异构酶酶活单位定义为:在上述方法的反应条件下,每分钟生成1 μmol果糖的酶量规定为1个活力单位(U).
1.3.4 菌落形态特征观察与系统发育分析
菌体形态特征鉴定通过采用结晶紫和碘液先后染色,乙醇脱色,番红复染的方法对菌株进行革兰氏染色,采用高倍显微镜观察菌体结构特征.
将纯化的菌株振荡培养至对数期,离心收集菌体,抽提基因组DNA.PCR扩增菌体基因组16S rDNA.引物16S-F: 5′-AAGGAGGTGATCCAGCC-3′,引物16S-R: 5′-AGAGTTTGATCCTGGCTCAG-3′.PCR程序参考文献[12],PCR扩增产物经用琼脂糖凝胶回收后,克隆到pMD18-T载体,转化大肠杆菌DH5α的感受态细胞.通过Blast程序,将由所测得的16S rDNA序列与GeneBank数据库中的16S rDNA序列进行相似性比较分析,选取同源性高的菌株后运用ClustalX 2.1进行比对分析,采用Mega 5.0程序,用 Neighbor-Joining算法,构建系统发育树[13].
1.3.5 酶活性质初步研究
酶学性质初步研究包括研究pH、温度、金属离子等对葡萄糖异构酶酶活力的影响[14].研究pH对葡萄糖异构酶酶活的影响:葡萄糖和粗酶液中加入200 μL的pH分别为7.0、7.4、7.8、8.2、8.6、9.0的缓冲液中,70 ℃反应15 min,按改进的硫酸咔唑法测定酶活[11].温度对葡萄糖异构酶酶活的影响:调节酶促反应的温度分别为60、65、70、75、80、85、90 ℃,缓冲液均为三乙醇胺一盐酸缓冲液(pH 7.4)-10 mmol·L-1MgSO4,测定酶活.金属离子对酶活力的影响:在最适温度和pH条件下,向三乙醇胺-盐酸缓冲液中分别添加终浓度为1 mmol·L-1Co2+, 5 mmol·L-1Mg2+, 1 mmol·L-1Ca2+, 1 mmol·L-1Mn2+, 1 mmol·L-1Fe3+,然后测定葡萄糖异构酶活力.
2 结果与分析
2.1 菌种的筛选和发酵
对采集的21个土壤样品进行筛选, 得到28株细菌和35株放线菌,依次命名为XJ-1~XJ-28,FXJ-1~FXJ-35.其中XJ-27、FXJ-8的筛选平板见图1a和图1b.所有细菌、放线菌分别液体培养发酵后制备胞内、外粗酶液.

图1 XJ-27(a)和FXJ-8(b)初筛平板图Fig.1 Preliminary screen plate diagram of XJ-27(a)and FXJ-8(b)
2.2 标准曲线的绘制及酶活力测定
采用改进的硫酸咔唑法测定28株细菌和35株放线菌的胞内、外粗酶液,测定结果见表1和表2.结果显示:细菌菌株XJ-27胞外酶活最高,为242.1 U·mL-1;放线菌菌株FXJ-8胞外酶活次之,为183.3 U·mL-1.

表1 28株细菌的胞内、外酶活统计Tab.1 Statistics of intracellular and extracellular enzyme activities of 28 strains of bacteria

表2 35株放线菌的胞内、外酶活统计Tab.2 Statistics of intracellular and extracellular enzyme activities of 35 strains of actinomyces
2.3 菌落形态特征观察与系统发育分析
产葡萄糖异构酶胞外酶活最高的XJ-27细菌菌株在培养基上菌落表面突出,为不透明淡黄色、边缘整齐,与培养基结合疏松,易挑取,见图1a.经革兰氏染色确定该菌株为革兰氏阳性菌,呈杆状,有芽孢,见图2.经BLAST与NCBI数据库中登录的序列比对分析,XJ-27的16S rDNA序列与Bacillus属的16S rDNA序列均有99%的相似性,可鉴定该菌株为Bacillus属细菌.菌株进一步选取Bacillus属的标准菌株构建进化树,结果表明,XJ-27菌株与BacilluscoagulansATCC7050距离最近,同处一筹(图3).因此,XJ-27菌株在系统发育地位上属于: Bacteria,Fimicutes,Bacillales,Bacillaceae,Bacillus.

图2 菌株XJ-27的革兰氏染色显微图(1000×)Fig.2 Gram staining micrograph of XJ-27(1000×)

图3 菌株XJ-27的16S rDNA序列的系统发育树 Fig.3 Phylogenetic tree of XJ-27 based on 16S rDNA sequence
2.4 酶活性质的测定
温度和缓冲液条件不变,在不同pH条件下测定XJ-27胞外酶活.以pH 7.4下测得的酶活作为参照(记为100%),结果如图4所示,该酶最适作用pH为7.0~7.8.pH和缓冲液条件不变,在不同温度下测定XJ-27胞外酶活,以70 ℃下测得的酶活作为参照(记为100%),结果如图5所示,该酶最适温度为70~80 ℃.不同离子下测定XJ-27胞外酶活,以含5mmol·L-1Mg2+缓冲液测得的酶活为参照(记为100%),结果如图6所示.XJ-27胞外酶活性依赖于Mg2+或Co2+.Ca2+、Mn2+、Fe3+金属离子对酶活力无明显的激活作用,Mg2+和Co2+有激活作用,Mg2+的效果最明显.

图4 XJ-27胞外酶的最适pHFig.4 Optimal pH of extracellular enzyme of XJ-27
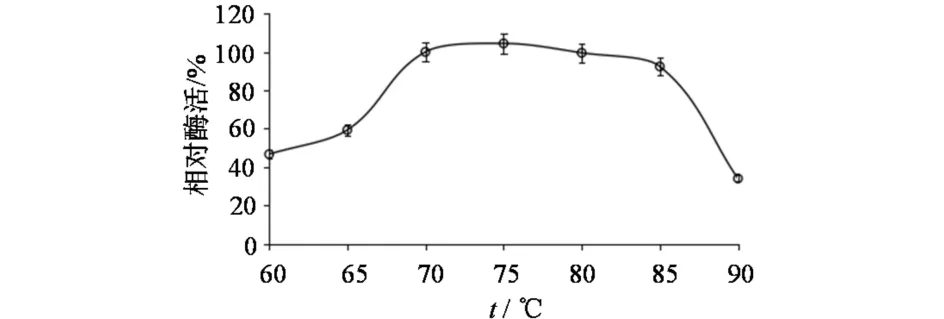
图5 XJ-27胞外酶的最适温度Fig.5 Optimal temperature of extracellular enzyme of XJ-27
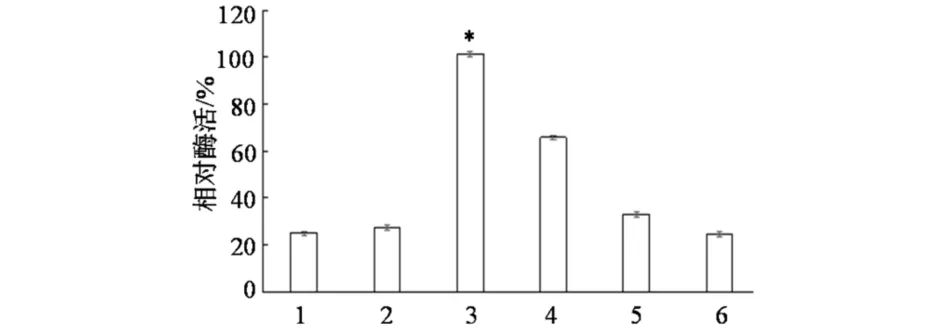
1)未加任何金属离子; 2)1 mmol·L-1 Ca2+; 3)5 mmol·L-1 Mg2+;4) 1 mmol·L-1 Co2+ ; 5) 1 mmol·L-1 Mn2+; 6)1 mmol·L-1 Fe3+*表示与未加任何金属离子相比,其酶活有显著性差异,P<0.05图6 金属离子对XJ-27胞外酶活力的影响Fig.6 Effect of metal ions on extracellular enzyme activity of XJ-27
3 讨论
本文筛选产葡萄糖异构酶菌株时采用改进的硫酸咔唑法测定酶活, 该法十分灵敏.测定时葡萄糖浓度及稀释倍数很关键, 因为葡萄糖浓度过高会对结果产生干扰.
细菌具有发酵产酶时间短、所产葡萄糖异构的酶的适用范围较广等优势.据报道,产细菌葡萄糖异构酶菌种主要集中在凝结芽孢杆菌(B.coagulans)和节杆菌(ArthrobacterNRRL B3724)[8],本研究筛选出的具最高胞外酶活的菌株XJ-27即为凝结芽孢杆菌.XJ-27酶活比张帆研究的大肠杆菌葡萄糖异构酶酶活(41.1 U·mL-1)高5.8倍[15],具有较强的应用研究前景.
在HFCS生产过程中, 葡萄糖转化率约为50%,而工业应用要求果糖含量高于50%.由于葡萄糖异构酶催化的葡萄糖异构化为果糖的反应为一可逆过程,在高温条件下,此反应的平衡偏向于果糖的生成[16-17].本文从土壤中筛选出的一株高产胞外葡萄糖异构酶的细菌菌株XJ-27在pH 7.4时其最适反应温度为70~80 ℃.因此,进一步提高其反应温度可增加果糖得率.提高酶的反应温度可从酶蛋白的分子结构方面进行研究,通过在分子中引入二硫键提高蛋白质的热稳定性,是蛋白质工程的常用方法[18-20].葡萄糖异构酶的活性二聚体或四聚体解聚成亚基结构可能是其热不稳定的重要原因之一.在亚基间构建二硫键,增强亚基间的相互作用,也有可能改善酶的热稳定性.
酶的最适pH一般由酶活性中心部位的静电环境,或者总的表面电荷所决定的,因此,改变酶作用的最适pH,可考虑通过改变静电环境来实现.对XJ-27菌株的葡萄糖异构酶进行分子结构研究,可使蛋白质工程方法定向改造葡萄糖异构酶变得切实可行.
猜你喜欢
辽宁化工(2022年8期)2022-08-27 06:03:04
石油炼制与化工(2021年12期)2021-12-14 06:26:38
江苏农业科学(2020年2期)2020-04-17 09:53:15
祝您健康(2019年7期)2019-07-12 03:11:52
天然产物研究与开发(2018年9期)2018-10-08 03:25:38
塑料助剂(2018年6期)2018-03-25 05:59:16
山东化工(2018年1期)2018-03-10 02:56:49
江苏农业科学(2017年10期)2017-07-21 17:00:28
湖北农业科学(2015年17期)2015-10-09 22:07:40
医学研究杂志(2015年8期)2015-06-22 14:00:54
