
土壤缺钾对冬小麦碳氮代谢的影响
2020-12-17 07:37:42刘晓伟王伟伟张浩月孙若希郝培显杨学举王殿武
麦类作物学报 2020年8期
刘晓伟,王伟伟,张浩月,孙若希, 郝培显,杨学举,赵 勇,王殿武
(1. 河北农业大学资源与环境科学学院,河北保定 071000;2. 河北农业大学农学院,河北保定 071000; 3. 沧州市农林科学院,河北沧州 061001 4. 河北农业大学生命科学学院,河北保定 071000)
冬小麦在我国种植广泛。钾素在冬小麦产量和品质形成过程中起着重要作用[1]。我国耕地中缺钾土壤的面积约占25%~33%,并且缺钾范围正在逐渐扩大[2];同时,我国钾盐资源储量少,钾肥利用率低[3]。因此,研究不同冬小麦品种生理代谢对土壤缺钾的响应具有重要意义。
钾素参与植株生理生化过程[4]。碳氮代谢作为植物最基本的两大代谢过程,二者之间存在很强的交互作用[5]。玉米缺钾时,叶片的可溶性总糖含量明显上升[6]。缺钾会降低棉花叶片净光合速率及蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)等活性,且通过影响硝酸还原酶活性限制氨基酸的合成[7]。缺钾时水稻蛋白质代谢受抑,游离氨基酸含量提高[8]。缺钾时大豆叶片谷氨酸脱氢酶(GDH)活性升高,谷氨酸合成酶(GS)活性下降[9]。以上结果表明,缺钾影响作物光合作用和碳代谢过程,抑制氮同化[4]。目前有关干旱、温度等逆境胁迫对小麦产量和品质形成影响机制的研究已经很多[10-11],但针对缺钾影响冬小麦碳氮代谢影响的研究尚少,且冬小麦生育后期是籽粒充实和产量、品质形成的关键时期。基于此,本研究以冬小麦品种科农9204(耐低钾型)和石新828(低钾敏感型)为材料,分析了土壤缺钾对冬小麦开花后碳氮代谢的影响,以期为揭示冬小麦耐低钾的调控机理提供依据。
1 材料与方法
1.1 供试材料
供试冬小麦品种为科农9204(耐低钾型)和石新828(低钾敏感型)。供试土壤为潮褐土 (<0.01 mm物理性粘粒含量为44.35%)和河流故道风沙土混配的土壤(<0.01 mm物理性粘粒含量为24.13%),有机质含量2.67 g·kg-1,全氮含量0.26 g·kg-1,全钾含量17.95 g·kg-1,缓效钾含量535.00 mg·kg-1,速效磷含量4.38 mg·kg-1,速效钾含量45.00 mg·kg-1。
1.2 试验设计
试验采用盆栽方法,在河北农业大学旱棚进行,共设缺钾(K1,45.00 mg·kg-1)和正常供钾(K2,180.00 mg·kg-1)两个处理,重复3次,每重复12盆,随机排列。 所用盆子为塑料盆,直径20 cm,高21 cm。每盆装土6 kg,底施尿素(N 46.4%)0.63 g和磷酸二铵(N 18.0%、P2O546.0%)0.94 g,并于冬小麦返青期追施尿素0.63 g,正常供钾处理每盆底施硫酸钾(K2O 53.0%) 1.53 g。小麦于2018年10月15日播种,三叶期定苗,每盆14株,2019年6月5日收获。试验期间进行严格的水分管理,且每次每盆浇水量相同,其他管理按常规大田。
1.3 测定指标及方法
土壤和植株养分含量的测定参照土壤农化分析标准[12];用简易比重计法测定土壤物理性粘粒含量。于冬小麦开花后0、7、14、21、28和35 d,每天9:00-11:00,采用SPAD-502型叶绿素仪测定旗叶中段叶绿素含量;然后,每重复取7片旗叶(清洗擦干)和部分籽粒,液氮速冻后,-80 ℃保存。参照石华荣[13]的方法提取旗叶蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)酶液。参考於新建[14]的方法进行测定SS和SPS活性;采用离体法测定籽粒中硝酸还原酶(NR)活性[15]。另取7片旗叶,105 ℃杀青30 min,80 ℃烘干至恒重后用于碳、钾含量测定。可溶性总糖、蔗糖、葡萄糖、果糖含量参照张志良等[16]方法的测定。籽粒经自然风干后凯氏定氮法测定氮含量,以系数5.7折算粗蛋白质含量,清蛋白、球蛋白、醇溶蛋白和麦谷蛋白组分的提取与分离参照唐云峰的方法[17],用考马斯亮蓝G-250法测定各蛋白组分的含量[18]。
1.4 统计方法
采用Excel 2010进行数据统计与作图,采用SAS18.0进行差异显著性分析。
2 结果与分析
2.1 土壤缺钾对冬小麦旗叶和籽粒钾素含量的影响
冬小麦旗叶和籽粒钾素含量因品种、生育时期不同而有差异(表1)。花后0~35 d,旗叶钾素含量呈先升后降的趋势,籽粒钾素含量呈下降趋势。2个冬小麦品种的旗叶钾素含量,均在花后 7 d最高,花后35 d最小,籽粒钾素含量均在籽粒形成初期(花后7 d)最高。与正常供钾处理(K2)相比,缺钾处理(K1)下冬小麦旗叶和籽粒钾素含量均不同程度下降,其中耐低钾型品种科农9204的旗叶钾素含量总体上变化较小,籽粒也无显著变化,而低钾敏感型品种石新828的旗叶和籽粒钾素含量总体上变化显著,降幅分别为5.01%~65.37%和4.74%~31.83%。这说明土壤缺钾会影响冬小麦旗叶和籽粒钾素含量,且对钾敏感型品种影响较大。
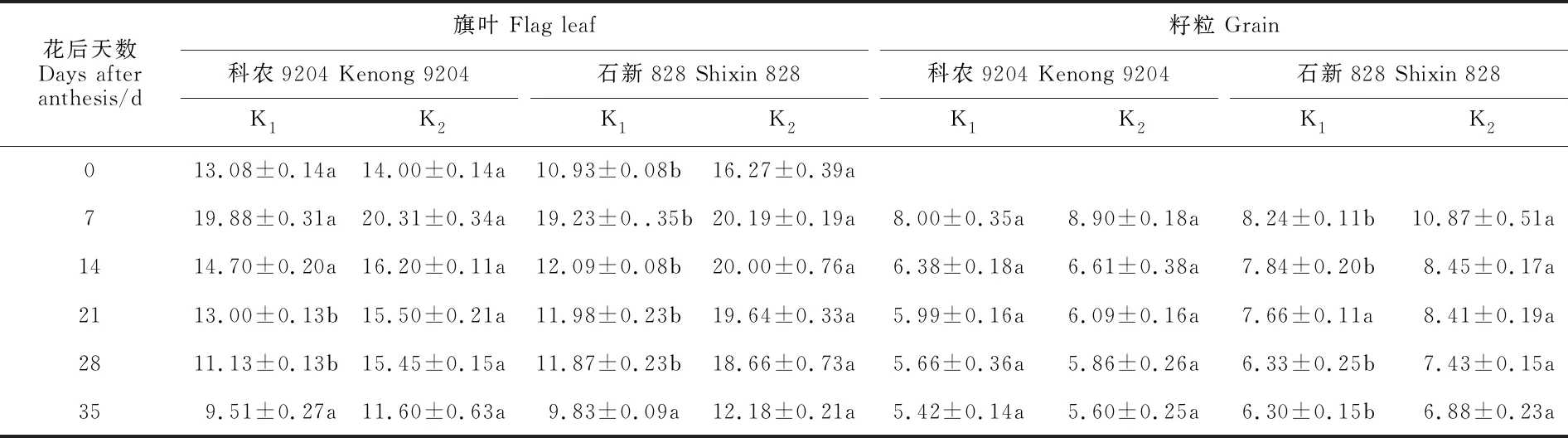
表1 土壤缺钾对冬小麦旗叶和籽粒钾含量的影响Table 1 Effect of soil potassium deficiency on potassium content in flag leaf and grain of winter wheat g·kg-1
2.2 土壤缺钾对冬小麦旗叶叶绿素含量的影响
2个冬小麦品种的旗叶叶绿素含量(SPAD值)整体上均呈先升后降趋势,且均在花后14 d达到最高值,花后35 d最小(图1)。在花后不同生育时期,缺钾时2个品种的旗叶SPAD值与正常供钾处理相比均不同程度降低,其中耐低钾型品种科农9204总体上变化不显著,而低钾敏感型品种石新828变化显著,降幅为5.27%~80.66%,表明土壤缺钾对冬小麦耐低钾型品种叶绿素的合成影响较小,而对低钾敏感型品种影响较大。
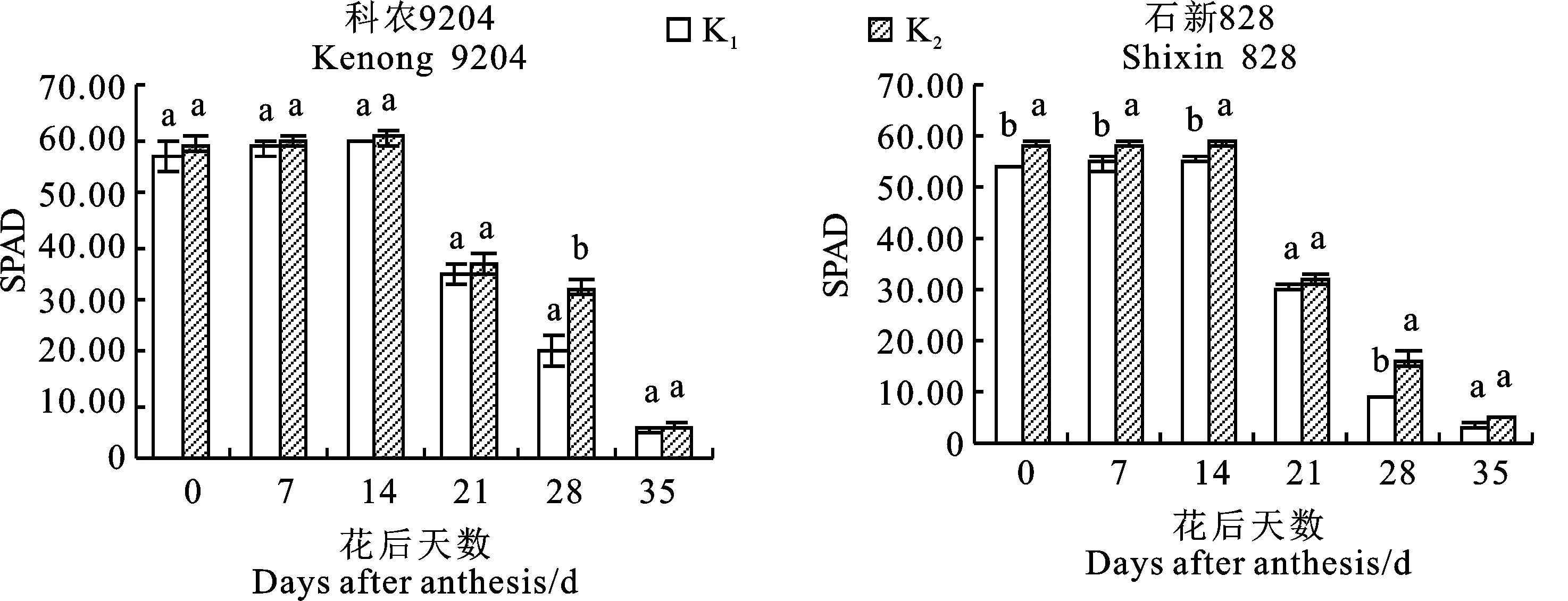
图柱上的小写字母表示同一时期不同处理间在0.05水平上差异显著。下图同。
2.3 土壤缺钾对冬小麦碳代谢的影响
2.3.1 土壤缺钾对冬小麦旗叶碳含量的影响
同一钾素处理下,2个冬小麦品种的旗叶碳含量均呈先升后降再升的趋势,其中耐低钾型品种科农9204的旗叶碳含量在花后7d最高,低钾敏感型品种石新828在花后14 d最高(表2)。缺钾处理下,两个品种的旗叶碳含量较正常供钾处理均不同程度下降,其中耐低钾型品种科农9204的两个处理间差异较小,总体上不显著,而低钾敏感型品种石新828差异较大,总体显著,降幅为3.29%~10.85%,表明土壤缺钾会影响冬小麦旗叶碳含量,其中对低钾敏感型品种影响较大。
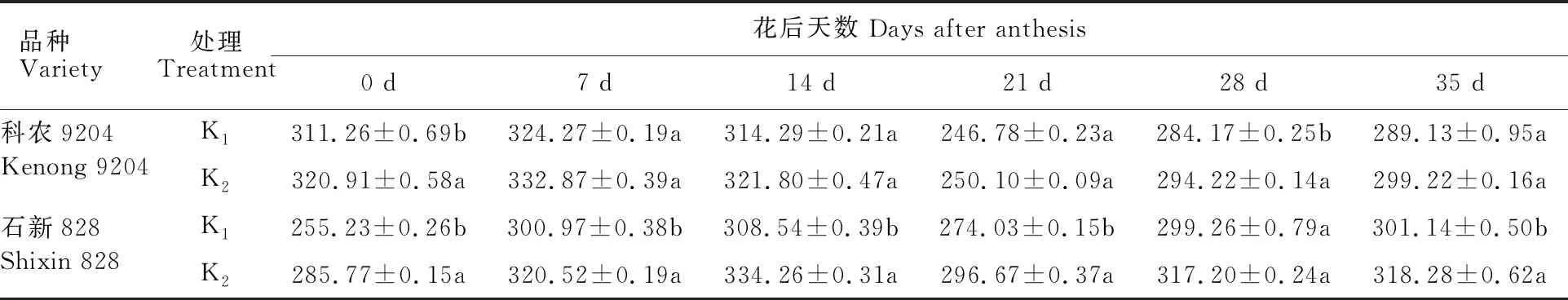
表2 土壤缺钾对冬小麦旗叶碳含量的影响Table 2 Effect of soil potassium deficiency on carbon content in flag leaves of winter wheat g·kg-1
2.3.2 土壤缺钾对冬小麦旗叶糖分含量的影响
缺钾处理下,2个品种的可溶性总糖、蔗糖、葡萄糖、果糖含量均高于正常供钾处理(图2),其中耐低钾型品种科农9204的4种糖含量与正常供钾处理差异均不显著,而低钾敏感型品种石新828的4种糖含量总体差异显著,增幅分别为 14.37%~119.08%、16.63%~68.00%、16.63%~ 68.00%、23.41%~80.37%,表明土壤缺钾影响了冬小麦糖代谢,且存在品种差异性。
2.3.3 土壤缺钾对冬小麦旗叶蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性的影响
与正常供钾处理相比,缺钾导致两个品种各时期的两种酶活性均不同程度下降(图3),其中耐低钾型品种科农9204只在花后7 d和14 d差异显著,而低钾敏感型品种石新828在花后0~21 d差异显著,降幅分别为32.28%~538.67%、48.51%~421.48%,说明土壤缺钾对冬小麦的两种酶活性有一定的抑制作用,且对敏感型品种影响较大。
2.4 土壤缺钾对冬小麦氮代谢的影响
2.4.1 土壤缺钾对冬小麦籽粒氮含量的影响
2个冬小麦品种籽粒氮含量随生育进程均呈上升趋势,在花后35 d达到最高值(表3)。与正常供钾处理相比,缺钾处理下2个品种各时期的籽粒氮含量均有所降低,其中耐低钾型品种科农9204仅在花后7 d和21 d差异显著,而低钾敏感型品种石新828除花后28 d外,在其余时期均差异显著,降低幅度为29.87%~ 90.33%。
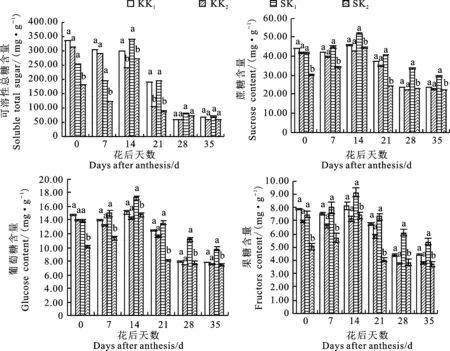
KK1和KK2分别为科农9024的K1和K1处理;SK1和SK2分别为石新828的K1和K2处理。下图同。

图3 土壤缺钾对冬小麦旗叶蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性的影响Fig.3 Effect of soil potassium deficiency on the activities of SS and SPS in flag leaves of winter wheat
2.4.2 土壤缺钾对冬小麦籽粒粗蛋白质含量及蛋白质组分的影响
2个冬小麦品种的籽粒粗蛋白质含量变化与氮含量一致,也随花后生育进程呈增长趋势(图4)。与正常供钾处理相比,缺钾处理下两个品种的籽粒粗蛋白含量均不同程度下降,其中科农9204仅在花后7和21 d差异显著,而石新828在各时期均差异显著,降低幅度为15.56%~ 66.12%,表明土壤缺钾使冬小麦籽粒粗蛋白的合成受限,对低钾敏感型品种影响较明显。
随花后生育时期的变化,2个冬小麦品种的籽粒清蛋白含量均呈下降趋势,籽粒球蛋白、醇溶蛋白和麦谷蛋白含量均呈上升趋势(图5)。与正常供钾处理相比,缺钾处理下,耐低钾型品种科农9204的籽粒清蛋白和球蛋白含量分别提高 1.47%~4.35%和10.66%~47.93%,籽粒醇溶蛋白和麦谷蛋白含量分别降低0.87%~9.14%和0.50%~4.88%,但仅醇溶蛋白含量在花后28和35 d差异显著。缺钾处理下低钾敏感型品种石新828的籽粒球蛋白含量较正常供钾处理提高2.57%~64.15%,清蛋白、醇溶蛋白和麦谷蛋白含量分别降低3.73%~26.02%、17.92%~ 38.81%和6.33%~35.81%,且不同时期差异多数达显著水平。这表明土壤缺钾影响了冬小麦 籽粒营养品质,尤其是对低钾敏感型品种影响 很大。

表3 土壤缺钾对冬小麦籽粒氮含量的影响Table 3 Effect of soil potassium deficiency on nitrogen content in grains of winter wheat g·kg-1
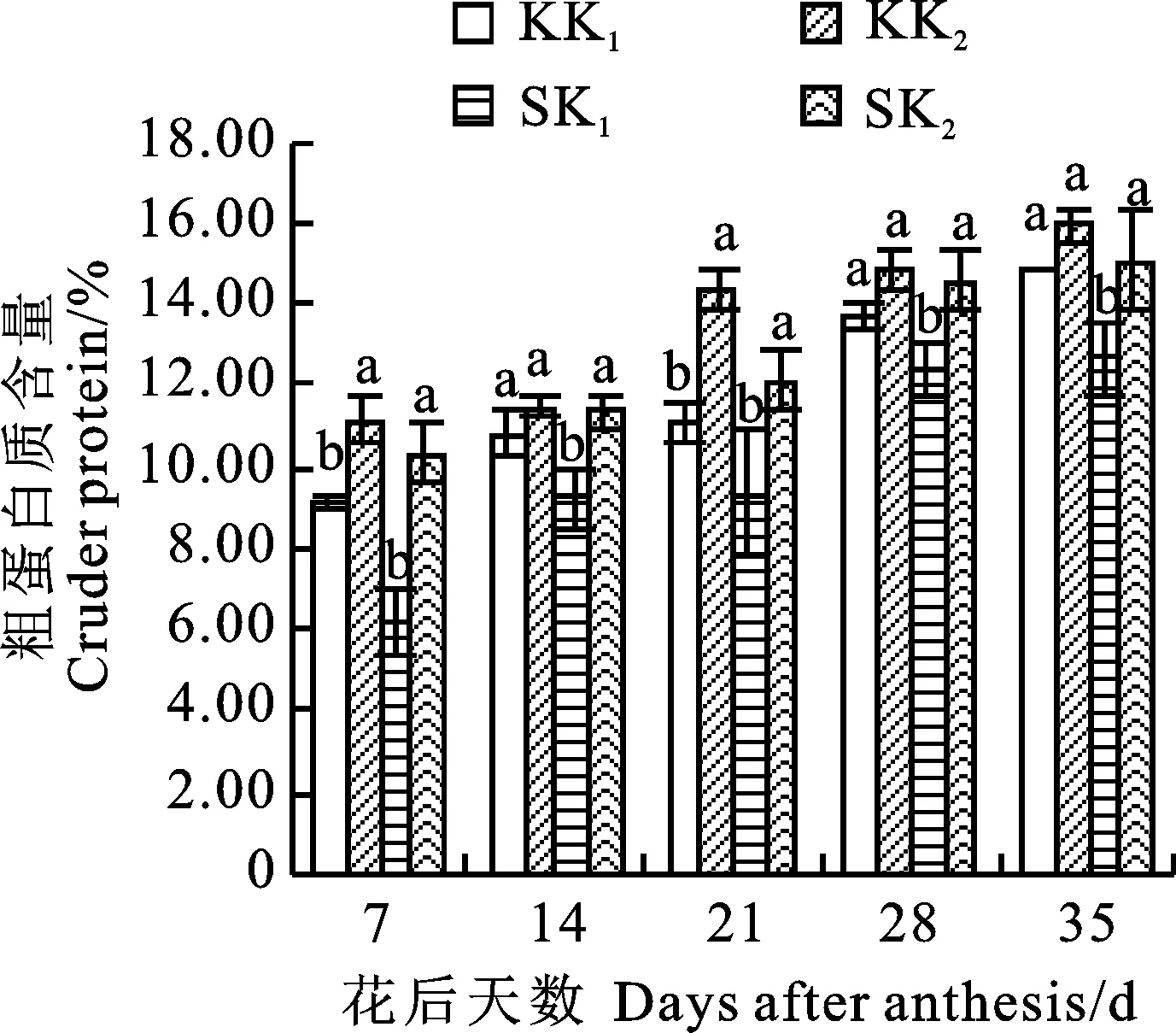
图4 土壤缺钾对冬小麦籽粒粗蛋白质含量的影响Fig.4 Effect of potassium deficiency in soil on crude protein content in grains of winter wheat
2.4.3 土壤缺钾对冬小麦籽粒硝酸还原酶(NR)活性的影响
与正常供钾处理相比,缺钾处理下两个小麦品种旗叶的NR活性均不同程度下降(图6),其中耐低钾型品种科农9204仅在花后各时期差异显著,而低钾敏感型品种石新828在花后14、21和35 d均差异显著,表明土壤缺钾对冬小麦氮代谢有一定的影响,且对低钾敏感型小麦的影响 较大。
3 讨 论
植物体内的钾素大多数来自于土壤,土壤缺钾会使得植物体内的钾素含量降低,且存在品种间钾吸收利用差异性[19]。本研究结果表明,与正常供钾相比,缺钾处理导致冬小麦旗叶和籽粒钾素含量均不同程度下降,且敏感型冬小麦总体差异显著。旗叶作为小麦的关键源器官和后期冠层的主要组成,其面积的大小和生理活性可能引起整个植株生理活性的改变[20],其叶绿素含量的多少决定了光合能力的强弱[21]。本研究发现,缺钾后冬小麦旗叶叶绿素含量降低。其原因一方面可能是缺钾使得冬小麦旗叶叶绿体数量减少,叶绿体的超微结构发生了变化[7];另一方面由于缺钾抑制了光合关键酶Rubisco及Rubisco活化酶活性,进而抑制了冬小麦的净光合速率[22]。
碳水化合物是光合作用卡尔文循环的最初产物,它既是能量的贮存者,也是植物合成其他有机物的起始物质,其含量常占植株干重的50%以上[23]。在本研究中,缺钾处理导致冬小麦旗叶碳含量下降,表明土壤缺钾影响了冬小麦碳的积累,进而影响冬小麦碳水化合物合成。研究表明,小麦叶片中可溶性碳水化合物是维系生存的重要代谢物质,能够缓解逆境胁迫对小麦植株的伤害[24]。在土壤缺钾条件下,棉花叶片葡萄糖,果糖,蔗糖和淀粉含量较正常供钾处理有所升高[4],叶片中蔗糖和淀粉含量也发生变化[22]。缺钾也会造成可溶性总糖在大豆叶片中的积累[25]。在本研究中,缺钾处理下,小麦旗叶可溶性总糖、蔗糖、葡萄糖、果糖含量有所升高,这是由于缺钾导致糖转运蛋白相关基因的表达下调,进而影响了糖的正常转运[26-27];另外,糖含量的增加能够维持细胞膨压,降低渗透式,使其适应逆境胁迫的生长环境[28]。在碳水化合物的代谢过程中涉及许多重要酶的参与,如蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)、胞质果糖-1,6二磷酸酶,均需要钾离子作为催化剂才能产生活性。SS和SPS的作用分别表现为催化蔗糖的降解以及控制蔗糖的合成。对小麦、玉米的光合碳氮代谢途径研究发现,PEPCase、SPS和NADH/NR三种酶的调节在碳、氮平衡中起着重要作用[29]。干旱胁迫对小麦灌浆期SS和SPS合成均产生不利影响[30]。本研究表明,缺钾处理的SS和SPS活性较正常供钾处理均有所降低,且敏感型冬小麦总体差异显著,推测是缺钾抑制了细胞中SS和SPS基因表达[29,31],进而影响到蔗糖的合成。

图5 土壤缺钾对冬小麦籽粒清蛋白、球蛋白、醇溶蛋白、麦谷蛋白含量的影响Fig.5 Effect of soil potassium deficiency on albumin,globulin,gliadin and glutemin contents in grains of winter wheat

图6 土壤缺钾对冬小麦籽粒硝酸还原酶(NR)活性的影响Fig.6 Effect of soil potassium deficiency on NR activity in grains of winter wheat

4 结 论
土壤缺钾条件下,耐低钾型和敏感型冬小麦品种旗叶、籽粒钾含量和旗叶叶绿素含量均有所降低,且敏感型品种总体差异显著;旗叶碳含量、蔗糖合成酶和蔗糖磷酸合成酶活性降低,各糖分含量不同程度升高;籽粒氮、粗蛋白质、醇溶蛋白、麦谷蛋白含量和硝酸还原酶活性降低,而清蛋白、球蛋白含量变化因品种而异。土壤缺钾导致光合作用减弱,减少同化产物,抑制了旗叶蔗糖合成酶和蔗糖磷酸合成酶活性,造成碳含量降低和各糖分含量积累;同时抑制了硝酸还原酶活性,使得籽粒氮、粗蛋白质含量降低,影响蛋白组分含量,导致冬小麦碳氮代谢失衡。
猜你喜欢
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
作物学报(2022年9期)2022-07-18 00:58:42
农业工程学报(2022年7期)2022-07-09 06:16:32
江苏农业科学(2022年4期)2022-03-11 21:10:40
干旱地区农业研究(2021年6期)2021-12-08 07:52:22
今日农业(2021年17期)2021-11-26 23:38:44
浙江农业科学(2019年11期)2019-11-09 01:37:48
今日农业(2019年11期)2019-08-15 00:56:32
农业科技与装备(2016年9期)2017-03-10 15:04:56
