
外源调节物质对铬(Cr6+)胁迫下小麦种子萌发的缓解效应
2020-12-17 12:13孙淑珍田中伟郑巧梅孙传蛟孙建云
麦类作物学报 2020年9期
孙淑珍,田中伟,郑巧梅,孙传蛟,孙建云
(1.南京农业大学农学院,农业农村部作物生理生态与生产管理重点实验室, 江苏南京 210095; 2.南京农业大学生命科学院,江苏南京 210095)
铬是水环境和土壤环境中最常见的重金属污染物之一。目前,我国约有0.2亿hm2耕地受铬等重金属的污染[1]。Cr6+是自然环境中最常见且稳定的形式,有很强的移动性和不可降解性,极易对植物造成伤害,并可通过食物链不断富集,进而危害人体健康,因此,Cr6+对植物的影响逐渐成为人们的研究热点[2]。
小麦是我国主要粮食作物之一,其种子萌发质量的高低是影响产量的关键因素。有研究表明,种子萌发时对Cr6+胁迫十分敏感,主要表现为种子内α-淀粉酶构象受损,活性降低[3],糖和蛋白质代谢发生紊乱[4],大量活性氧生成,破坏膜脂并造成氧化损伤[5],导致种子萌发率明显下降。因此,保证小麦种子在Cr6+污染环境中正常萌发、出苗,是维持其产量平稳的前提。添加适合的外源调节物质能够提高重金属胁迫下种子的萌发率,降低重金属在其植株可食部位的积累[6]。前人已有报道证实,葡萄籽提取物(grape seed extract,GSE)[7]、氯化血红素(hematin chloride,Hemin)[8]和茉莉酸甲酯(methyl jasmonate,MeJA)[9]可有效缓解动植物重金属胁迫,但它们对Cr6+胁迫下植物生长的影响及其相应机制仍不明确。且已有研究只限于外施一种调节物质,而未比较分析多种调节物质对同一植物在Cr6+胁迫下的缓解效应及其相关机制上的差异。本研究利用水培试验在小麦种子萌发期分别添加GSE、Hemin和MeJA,探讨这3种物质对Cr6+毒害下小麦生理反应的影响,以期明确它们在缓解Cr6+毒害、提高发芽率和出苗率等方面的潜在作用。
1 材料与方法
1.1 试验设计
试验为三因素(小麦品种、Cr6+浓度、调节物质种类)设计。根据Xu等[10]的小麦耐Cr6+性筛选结果,选取豫麦51(敏感型)和扬麦16(耐铬型)为供试材料。小麦种子用20%的双氧水消毒10 min后,迅速用蒸馏水清洗数次,均匀摆放至铺有双层滤纸的培养皿内。Cr6+处理浓度设0和100 μmol·L-1两个水平,Cr6+以重铬酸钾(K2Cr2O7)的形式溶于蒸馏水或对应浓度调节物质中。调节物质设100 μmol·L-1GSE、50 μmol·L-1Hemin、 1 μmol·L-1MeJA三个处理,以上处理浓度均由预试验筛选得出,以蒸馏水为对照(CK)。每个处理设置10个重复(每皿50粒,一个培养皿为一个重复),小麦种子置于 24 ℃/16 ℃(12 h/12 h)光照培养箱培养。
1.2 测定项目与方法
种子萌发3 d时,取种子鲜样测定α-淀粉酶活性。处理7 d时,统计发芽率并取鲜样和干样,同时在每个处理和每个培养皿里随机取20颗发芽的小麦种子,测定根系和芽长。

1.3 数据处理
数据采用Excel和SPSS 10.0进行统计分析。
2 结果分析
2.1 外源调节物质对Cr6+胁迫下小麦种子萌发的影响
由表1可知,在无Cr6+胁迫的情况下,添加调节物质,两品种的各指标均无明显差异。Cr6+胁迫显著抑制了小麦种子的萌发以及萌发后幼苗的生长。主要表现为,与无Cr6+胁迫处理下的对照相比,豫麦51种子萌发时的发芽率、根系干物质积累量、地上部干物质积累量、根长和株高分别降低了25.9%、37.0%、25.0%、35.8%和 29.6%;扬麦16分别下降了13.9%、27.8%、 19.0%、23.5%和21.8%。与Cr6+胁迫处理下的对照相比,分别添加GSE、Hemin和MeJA后,豫麦51和扬麦16的上述指标均显著上升,其中添加GSE后,豫麦51和扬麦16的发芽率分别上升了25.4%和10.7%,Hemin处理后分别上升了24.9%和7.3%,MeJA处理后分别上升了 15.8%和6.3%。上述结果表明,外源调节物质可减轻Cr6+胁迫对小麦种子萌发的毒害作用,促进Cr6+胁迫下小麦种子的萌发与生长。

表1 外源调节物质对Cr6+胁迫下小麦种子萌发和幼苗生长的影响Table 1 Effect of exogenous regulators on seed germination and seedling growth of wheat under Cr6+ stress
2.2 外源调节物质对Cr6+胁迫下小麦种子α-淀粉酶和渗透调节物质的影响
由表2可知,Cr6+胁迫显著降低了小麦种子的α-淀粉酶活性,与无Cr6+胁迫处理下的对照相比,豫麦51降低了25.7%,扬麦16降低了 20.0%。与Cr6+胁迫处理下的对照相比,添加GSE、Hemin和MeJA均显著提高了小麦种子α-淀粉酶的活性,其中豫麦51分别提高了25.5%、 23.6%和14.5%,扬麦16分别提高了20.0%、11.7%和5.0%。表明外源调节物质可通过提高α-淀粉酶活性,诱导小麦种子在Cr6+胁迫下快速打破休眠。
在无Cr6+胁迫的情况下,添加调节物质后,两个小麦品种根系中可溶性糖含量均显著提高。Cr6+胁迫显著提高了两品种根系中可溶性糖含量,且扬麦16远高于豫麦51。而与Cr6+胁迫处理下的对照相比,分别添加GSE、Hemin和MeJA后,豫麦51可溶性糖含量分别提高了9.6%、26.0%和 27.2%;扬麦16添加Hemin后可溶性糖含量提高了13.1%,而添加GSE和MeJA分别下降了22.7%和14.9%。Cr6+胁迫处理下,两小麦品种根系中可溶性蛋白含量显著增加,添加调节物质后其含量进一步增加 。与Cr6+胁迫处理下的对照相比,添加GSE、Hemin和MeJA后,豫麦51根系中可溶性蛋白含量分别提高了 10.5%、2.5%和8.9%,扬麦16分别提高了 19.5%、14.7%和10.3%。表明外源调节物质在一定程度可增强小麦根系的渗透调节调节能力。
2.3 外源调节物质对Cr6+胁迫下小麦根系MDA及活性氧(reactive oxygen species,ROS)含量的影响
由表3可知,在无Cr6+胁迫的情况下,添加调节物质对小麦根系MDA的含量无明显影响。Cr6+胁迫显著提高了两小麦品种根系MDA的含量,其中豫麦51根系MDA含量的升高幅度显著大于扬麦16。与Cr6+胁迫处理下的对照相比,添加调节物质后,两小麦品种根系MDA的含量均显著降低。在外源添加GSE后,小麦根系MDA含量下降幅度最大,其次是Hemin,最后为MeJA。

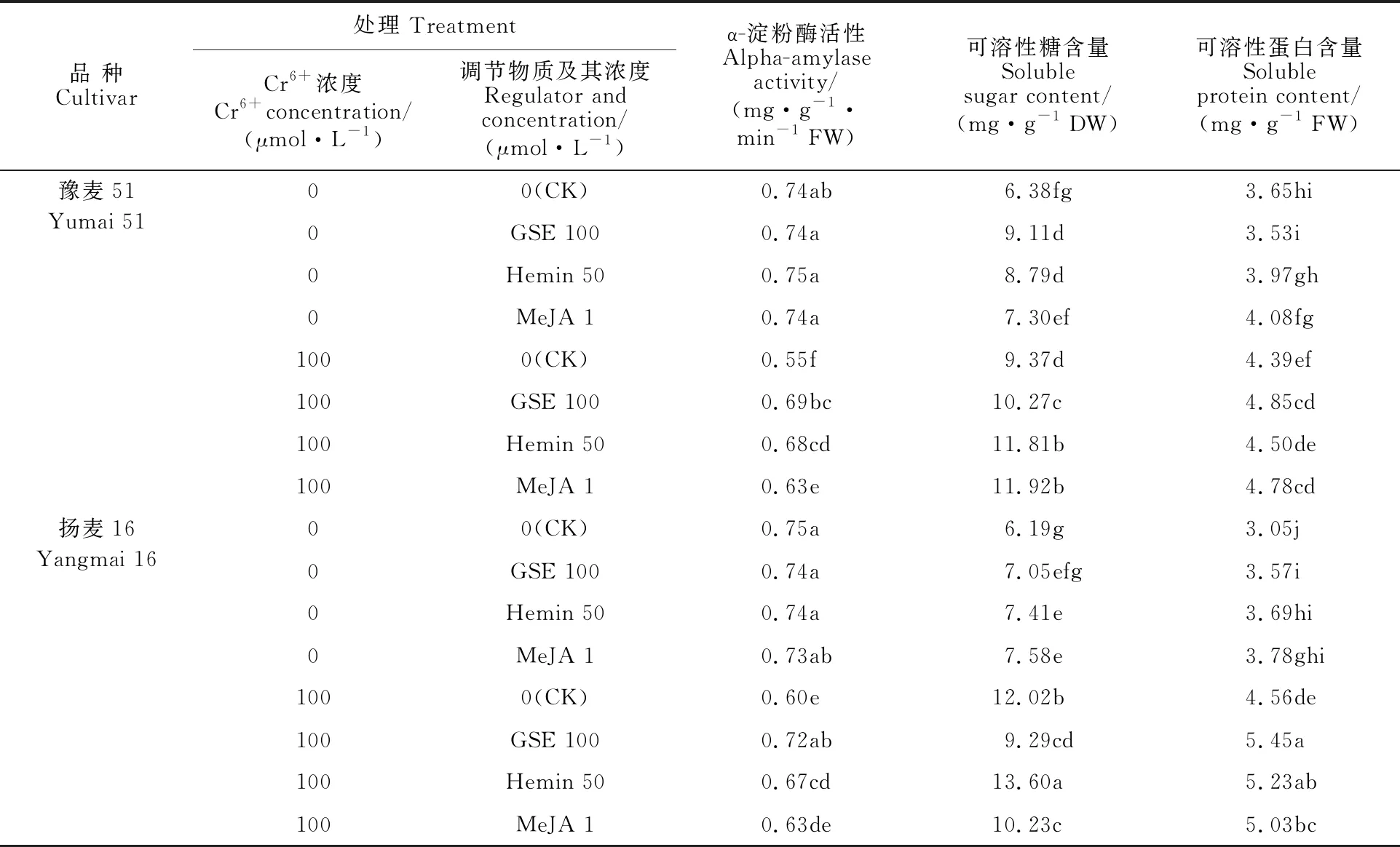
表2 外源调节物质对Cr6+胁迫下小麦种子α-淀粉酶活性和渗透调节物质含量的影响Table 2 Effect of exogenous regulators on alpha-amylase activity and osmotic adjustment substance content of wheat seed under Cr6+ stress

表3 外源调节物质对Cr6+胁迫下小麦根系和H2O2含量的影响Table 3 Effect of exogenous regulators on and H2O2 content of wheat root under Cr6+ stress
2.4 外源调节物质对Cr6+胁迫下小麦根系抗氧化酶活性的影响
由表4可知,与无Cr6+胁迫处理下的对照相比,Cr6+胁迫显著影响了两个小麦品种根系被测抗氧化酶的活性,其中豫麦51根系的SOD和CAT活性显著降低,APX和GR活性显著增加;扬麦16根系的SOD活性也呈降低趋势,但其下降幅度低于豫麦51,根系其他抗氧化酶活性均显著增加。与Cr6+胁迫处理下的对照相比,添加GSE、Hemin和MeJA后,两个小麦品种根系的SOD、CAT、APX和GR活性均显著上升(扬麦16添加Hemin后的GR活性除外)。表明外源调节物质能够通过提高小麦根系抗氧化酶的活性,清除由Cr6+产生的过量ROS ,维持细胞内部ROS稳态,进而提高小麦种子对Cr6+胁迫的适应性。

表4 外源调节物质对Cr6+胁迫下小麦根系抗氧化酶活性的影响Table 4 Effect of exogenous regulators on antioxidase activity of wheat root under Cr6+ stress
3 讨 论
Cr6+对土壤和水系的长期污染严重抑制了植物种子的萌发。本研究中,Cr6+胁迫对小麦种子萌发期的毒害症状主要表现为发芽率降低、干物质积累量减少等,这与Velez等[19]对鹰嘴豆的研究结论一致。Cr6+胁迫对两小麦品种根系的毒害作用远大于对芽,可能是由于Cr6+直接接触小麦根系,大量的Cr6+与细胞结合滞留于根部,导致ROS的大量生成,从而引发了细胞代谢受阻和细胞凋亡[20]。本研究发现,与Cr6+胁迫处理下的对照相比,外源同时添加调节物质后,两小麦品种发芽率、干物质积累量、根长和株高均显著增加,增加幅度依次为GSE>Hemin>MeJA。α-淀粉酶活性的高低影响种子萌发的速度。本研究发现,Cr6+胁迫显著降低了两小麦品种种子内α-淀粉酶的活性,进而使种子内淀粉水解速度变慢。原因可能是,α-淀粉酶是Ca2+依赖性酶[21],高浓度Cr6+进入种子内部与Ca2+竞争淀粉酶活性位点,致使α-淀粉酶的构象发生变化,限制小麦种子萌发。与Cr6+胁迫处理下的对照相比,添加3种调节物质后,小麦种子的α-淀粉酶活性均有不同程度的上升,其中添加GSE的酶活性与无Cr6+胁迫处理下的对照差异最小。这可能是外源调节物质降低了Cr6+胁迫下小麦根系的氧化应激,保护膜结构,从而维持小麦生理生化稳态。
本研究发现,在无Cr6+胁迫的情况下,添加调节物质后,两小麦品种根系内可溶性糖和可溶性蛋白含量均增加(豫麦51添加GSE后的可溶性蛋白含量除外)。这表明外源调节物质有利于小麦胚乳内淀粉和蛋白质的水解,为种子萌发供应了丰富的物质合成碳架。Cr6+胁迫下小麦根系的渗透调节物质显著增加,表明小麦种子萌发对Cr6+胁迫具有自我调节能力。添加外源调节物质后,小麦根系的可溶性糖和可溶性蛋白含量进一步增加,这表明外源调节物质有利于小麦进一步提高这种自我调节能力,有助于缓解Cr6+胁迫下由于膜结构破坏而引起的渗透压不平衡,使小麦根系保持相对较低的水势,能从胁迫环境中吸收更多水分和养分,进而促进萌发后小麦幼苗的生长,这与Handa等[6]对芥菜的研究结果一致。而略有不同的是,在Cr6+胁迫下,外源添加GSE和MeJA后,扬麦16根系可溶性糖含量下降,推测是由于扬麦16的耐Cr6+性较豫麦51强,在外源添加GSE和MeJA后,渗透胁迫降低,造成根系可溶性糖含量减少,也有可能是不同缓解物质对不同品种小麦的缓解效应不同导致。
4 结 论
Cr6+胁迫通过抑制淀粉酶活性,打破植株渗透平衡和加剧氧化损伤,从而抑制了小麦种子的萌发。添加调节物质可以有效增强小麦的耐Cr6+性,主要表现在:提高小麦种子萌发期α-淀粉酶活性,增加种子萌发过程中的能量物质供应;调节小麦幼苗根系可溶性糖以及可溶性蛋白的含量,保证渗透稳态,促进小麦萌发过程中的各种代谢;提高根系抗氧化酶活性,降低氧化应激,最终缓解Cr6+胁迫。外源调节物质对Cr6+胁迫的缓解效果为GSE>Hemin>MeJA。
猜你喜欢
农业科技通讯(2022年11期)2022-11-30
现代农业科技(2022年20期)2022-11-09
舰船科学技术(2022年11期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
麦类作物学报(2021年1期)2021-04-12
临床检验杂志(电子版)(2020年3期)2020-04-11
CHINESE JOURNAL OF AERONAUTICS(2019年10期)2019-12-19
科技与创新(2017年13期)2017-07-19
人人健康(2017年11期)2017-06-02
食品工业科技(2014年23期)2014-03-11
