
小麦光温敏雄性不育系穗发芽抗性鉴定及相关分子标记验证
2020-12-17 12:13侯起岭赵昌平杨卫兵高建刚陈现朝杨吉芳白秀成张立平张风廷
麦类作物学报 2020年9期
侯起岭,赵昌平,杨卫兵,高建刚,陈现朝, 杨吉芳,白秀成,张立平,张风廷,孙 辉
(北京杂交小麦工程技术研究中心/杂交小麦分子遗传北京市重点实验室,北京 100097)
小麦穗发芽(pre-harvest sprouting,PHS)是指小麦在成熟期遇到连续阴雨天气或处在潮湿环境下,籽粒在穗部发芽的现象。小麦穗发芽是一种世界性灾害,不仅导致籽粒产量降低,而且降低小麦加工品质和种子质量[1]。小麦光温敏雄性不育系在可育条件下繁殖,在不育条件下制种,成熟期如遇连续阴雨天气,将严重降低穗发芽率较高的不育系繁殖和杂交小麦制种的种子质量。因此,抗穗发芽小麦不育系种质的挖掘和抗穗发芽品种选育是杂交小麦育种的重要课题之一。
小麦穗发芽受多种因素的影响,包括籽粒休眠特性、穗部性状、淀粉酶活性、植物内源激素和外部环境(如温度、湿度)等,其中籽粒休眠特性是主要因素[2-3]。小麦穗发芽的鉴定受外部环境因素影响较多,鉴定难度较大,目前主要鉴定方法有整穗发芽法、籽粒发芽法、大田发芽法、酶反应生化标记选择法等。整穗发芽法、籽粒发芽法是常用方法;整穗发芽法能直观反映品种的总体穗发芽抗性,籽粒发芽法主要反映籽粒的休眠特性,但不能反映品种穗发芽的综合抗性。分子标记辅助选择法能够从早代准确选择目标,开发、验证并利用有效的分子标记是鉴定小麦穗发芽抗性的有效方法之一[4]。
小麦穗发芽的抗性机制较为复杂,是由主效基因和微效基因(QTLs)控制的遗传性状,根据穗发芽抗性相关的QTL位点开发有效的分子标记是分子标记辅助育种的关键。目前,已发现了一系列与穗发芽抗性相关的QTLs,主要集中在3AS和4AL染色体上[5]。Yang等[6]发现,Vp1基因的等位变异基因Vp1B与穗发芽抗性相关;标记Vp1B3在抗穗发芽品种中可扩增出845 bp或569 bp片段,在感穗发芽品种中可扩增出652 bp片段。Mares等[7]在小麦籽粒颜色合成相关基因(R基因)的作用机制研究中发现,位于4A染色体上的一个SSR标记Xwmc468与小麦种子的休眠特性相关。刘光辉等[4]利用发芽指数和整穗发芽率作为小麦穗发芽抗性鉴定指标,发现标记Xwmc468与穗发芽抗性相关。小麦TaSdr基因与水稻种子休眠相关基因OsSdr4是同源基因,对小麦种子休眠及穗发芽抗性起重要调控作用;基因序列分析表明,TaSdr-B1在其起始密码子上游存在一个SNP(A/G),A类型为TaSdr-B1a,G类型为TaSdr-B1b,通过RIL群体(Yangxiaomai/Zhongyou 9507)的验证发现,TaSdr-B1b基因型的GI(germination index)值显著高于TaSdr-B1a基因型[8]。研究发现,TaMFT-A1基因位于小麦3AS上,可以调控小麦籽粒的休眠,抑制籽粒的萌发,与抗休眠QTL位点QTsg.osu-3A紧密关联[9]。TaMFT-A1基因在春小麦品种中国春(Chinese Spring,CS)中与种子休眠调控有关,其Jagger等位变异类型在冬小麦和春小麦品种间存在差异[10]。Liu等[11]从白粒小麦Rio Blanco中克隆了一个位于小麦3AS染色体上的控制穗发芽抗性基因TaPHS1,该基因是TaMFT的同源基因,对小麦穗发芽抗性起正向调控作用。
小麦穗发芽抗性易受环境因素影响,很多已经开发的分子标记只在特定环境和遗传背景下有效,很难在小麦育种中被广泛应用,已开发的标记对于具有不同遗传背景小麦的适用性和有效性还需要进一步验证。鉴于此,本研究利用STS标记Vp1B3,SSR标记Xwmc468和基于小麦穗发芽抗性基因开发的KASP标记TaSdr-B1、TaMFT-721J、TaPHS1-646和TaPHS1-646,结合整穗发芽法对88份小麦光温敏雄性不育系进行穗发芽抗性鉴定,以期筛选出抗穗发芽的不育系品系,为育种提供优质亲本,同时筛选出能有效评价小麦光温敏不育系穗发芽抗性的分子标记,为杂交小麦育种提供参考。
1 材料与方法
1.1 材 料
供试材料为北京杂交小麦工程技术研究中心选育的88份不育系,于2018年秋季在北京顺义试验基地(40°08′N,116°39′E)种植,试验采取随机区组排列,每份材料4行,行长1.5 m,株距 0.03 m,行距0.25 m,3次重复,常规田间管理。
1.2 方 法
1.2.1 整穗发芽率测定
参照Yang等[6]的方法并稍作调整,将穗发芽抗性水平划分为4类,分别为高抗、中抗、中感和高感,对应的发芽率依次为0~10%、10%~30%、30%~60%和60%~100%。在小麦蜡熟期,每个品系的每个重复选取成熟一致的主茎穗10个,室温下自然风干后置于-20 ℃冰箱存放,以维持种子休眠特性。所有供试麦穗都用0.5%的NaClO表面消毒20 min,在蒸馏水中浸泡洗净,放在不同的塑料自封袋中,每个重复做好标记,置于25 ℃恒温培养箱中发芽。发芽期间保持培养箱湿度,每天查看穗子发芽状况。7 d后取出立即放置于60 ℃烘箱中烘干24 h,然后手工脱粒,记录总穗粒数及发芽粒数,计算整穗发芽率(spike germination rate,SGR)。整穗发芽率=10个穗子总发芽籽粒数/10个穗子总籽粒数×100%。三次重复。
1.2.2 基因组DNA提取
每份材料取6~8粒种子置于铺有滤纸的培养皿中培养,萌发后取适量叶片装入2.0 mL的离心管中,加入钢珠,液氮中研磨后用CTAB法[13]提取基因组DNA。
1.2.3 分子标记检测
分子标记引物由北京六合华大基因科技有限公司合成,引物序列见表1。标记Vp1B3和Xwmc468的PCR扩增体系为20 μL,包括2×Mix混合液10 μL,模板DNA 2 μL(200 ng·μL-1),上、下游引物各1 μL(2 μmol·μL-1),ddH2O 5.0 μL;反应程序为:95 ℃预变性5 min;95 ℃变性1 min,58 ℃退火1 min,72 ℃延伸1 min ,35个循环;72 ℃延伸10 min。Vp1B3的反应产物用1.5%琼脂糖凝胶电泳进行检测,Xwmc468的反应产物用6%聚丙烯酰胺凝胶电泳进行检测。
KASP标记检测采用QuantStudioTM7 Flex实时定量PCR进行。PCR反应体系为:模板DNA 1.5 μL(50 ng·μL-1),2×Master mix 0.75 μL,引物0.0417 μL,ddH2O补充至3.0 μL。PCR反应程序为:I: 94 ℃ 15 min,1次循环;II:94 ℃变性20 s,61 ~55 ℃,60 s 10次循环(每次循环降低0.6 ℃);III:94 ℃ 20 s,57 ℃ 60 s,26次循环。以ddH2O为空白对照。

表1 抗穗发芽相关分子标记Table 1 Markers associated with PHS tolerance applied in this study
1.3 数据分析
用Excel 2010进行数据统计,采用SPSS 22.0进行方差及相关性分析。
2 结果与分析
2.1 供试不育系材料的整穗发芽率
88份供试不育系材料的整穗发芽率为 3.75%~93.23%,平均整穗发芽率为45.49%,变异系数为0.59。由表2可知,各抗性等级间小麦整穗发芽率(SGR)差异明显,SGR小于10%的材料有10份,平均值为 6.51%,占比为11.36%,抗性等级为高抗;SGR在10%~30%之间的材料有19份,平均值为 20.23%,占比为21.59%,抗性等级为中抗;SGR在30%~60%之间的材料有25份,平均值为39.22%,占比为28.41%,抗性等级为中感;SGR在60%~100%之间的材料有34份,平均值为75.69%,占比为38.64%,抗性等级为高感。

表2 供试材料整穗发芽分析Table 2 Analysis of sprouting of the tested materials

表3 88份小麦光温敏雄性不育系的穗发芽抗性及6个标记的扩增产物分析Table 3 PHS tolerance and amplified products of six markers in 88 wheat sterile lines
(续表3 Continued table 3)
2.2 不同穗发芽抗性标记的有效性分析
2.2.1 STS标记Vp1B3鉴定结果
由表3可知,在88份小麦光温敏雄性不育系材料中,共扩增出片段长度为569 bp、652 bp和845 bp三种Vp1B3类型的条带,分别命名为Vp1B3a、Vp1B3b和Vp1B3c,部分结果如图1所示。供试材料中,扩增出Vp1B3a类型片段的有24份,占供试材料数的27.27%,平均SGR为34.73%,变化范围为3.75%~84.75%;扩增出Vp1B3b类型片段的有59份,占供试材料数的67.05%,平均SGR为51.77%,变化范围为 3.87%~ 93.23%;扩增出Vp1B3c类型片段的有5份,占供试材料数的5.68%,平均SGR为23.12%,变化范围为7.63%~42.00%。相关性分析表明,Vp1B3a类型片段与SGR值呈显著负相关关系;Vp1B3b类型片段与SGR值呈显著正相关关系;Vp1B3c类型片段与SGR值相关性不显著(表4)。扩增出Vp1B3a和Vp1B3c类型片段的材料的平均SGR明显低于Vp1B3b类型片段的材料,说明标记Vp1B3可用于供试材料的穗发芽抗性鉴定。

M:D2000 marker; 1:BS104; 2:BS212; 3:BS252; 4:BS206; 5:BS283; 6:BS237.
2.2.2 SSR标记Xwmc468鉴定结果
用标记Xwmc468共扩增出A、B、C和D共4种类型的片段,部分结果如图2所示。扩增出A类型片段的材料有9份,占总材料的10.23%,平均SGR为53.30%,变化范围为4.73%~ 93.23%;扩增出B类型片段的材料有65份,占供试材料的73.86%,平均SGR为46.91%,变化范围为3.87%~91.68%;扩增出C类型片段的材料有6份,占供试材料的6.82%,平均SGR为47.63%,变化范围为8.39%~81.18%;扩增出D类型片段的材料有8份,占供试材料的 9.09%,平均SGR为23.57%,变化范围为 3.18%~34.38%。扩增出D类型片段材料的SGR平均值明显低于其他类型片段的材料。相关性分析表明A、B和C类型片段与SGR值的相关性均不显著;D类型片段与SGR值呈显著负相关(表4)。由此可见,标记Xwmc468可用于供试材料的穗发芽抗性鉴定。
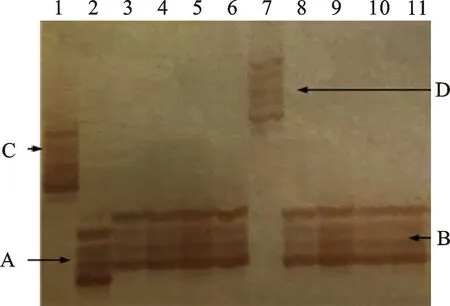
1:BS265; 2:BS104; 3:BS177; 4:BS293; 5:BS283; 6:BS182; 7:BS129; 8:BS270; 9:BS485; 10:BS166; 11:BS130.
2.2.3TaSdr-B1鉴定结果
用标记TaSdr-B1在供试材料中共检测出TaSdr-B1a和TaSdr-B1b两种等位变异类型(图3)。其中TaSdr-B1a等位变异类型有26份,占供试材料的29.55%,SGR平均值为45.06%;TaSdr-B1b等位变异类型有54份,占供试材料的61.36%,SGR平均值为46.33%(表3)。TaSdr-B1a和TaSdr-B1b两种等位变异类型品系间的SGR值无显著差异,二者与SGR的相关性均不显著(表4)。
2.2.4TaPHS1-646鉴定结果
用标记TaPHS1-646在88份供试材料中共检测出Rio Blanco和NW97S186两种等位变异类型(图3)。其中Rio Blanco等位变异类型有78份,占供试材料的88.64%,SGR平均值为 42.99%;NW97S186等位变异类型有7份,占供试材料的7.95%,SGR平均值为76.20%(表3)。Rio Blanco和NW97S186两种等位变异类型品系间的SGR值差异显著;Rio Blanco等位变异类型与SGR值呈显著负相关关系,NW97S186等位变异类型与SGR值呈显著正相关关系(表4)。表明标记TaPHS1-646可用于供试材料的穗发芽抗性鉴定。
2.2.5TaPHS1-666鉴定结果
用标记TaPHS1-666在供试材料中共检测出Rio Blanco和NW97S186两种等位变异类型(图3)。其中Rio Blanco等位变异类型有64份,占供试材料的72.73%,SGR平均值为44.10%;NW97S186等位变异类型有8份,占供试材料的9.09%,SGR平均值为70.19%(表3)。RioBlanco和NW97S186两种等位变异类型品系间的SGR值差异显著,Rio Blanco等位变异类型与SGR值相关不显著,NW97S186等位变异类型与SGR值呈显著正相关(表4)。
2.2.6TaMFT-721J鉴定结果
用标记TaMFT-721J在供试材料中共检测出Jagger和CS两种等位变异类型(图3)。其中Jagger等位变异类型有42份,占供试材料的 47.73%,SGR平均值为45.79%;CS等位变异类型有36份,占供试材料的40.90%,SGR平均值为46.56%(表3)。Jagger和CS两种等位变异类型品系间的SGR值差异不显著,二者与SGR的相关性均不显著(表4)。

红色点:HEX基因型;蓝色点:FAM基因型;黑色点:空白对照。图A为 TaSdr-B1标记检测结果;图B为 TaPHS1-646标记检测结果;图C为 TaPHS1-666标记检测结果;图D为 TaMFT-721J标记检测结果。
3 讨 论
穗发芽抗性是受多基因调控、多因素影响的复杂性状,抗性机理十分复杂。目前已开发出很多与种子休眠或穗发芽相关的的分子标记,但这些标记对不同基因型小麦品种的有效性还需进一步验证。本研究采用整穗发芽的鉴定方法,同时选择6个被认为与穗发芽相关的分子标记对88份小麦光温敏雄性不育系的穗发芽抗性进行鉴定。结果显示,整穗发芽率低于10%的有10份,占供试材料的11.36%,表现为高抗;在10%~30%之间的有19份,占比为21.59%,表现为中抗;这说明小麦光温敏雄性不育系中存在丰富的抗穗发芽材料。其中有4个标记的部分基因型与穗发芽抗性显著相关,与前人研究结果一致[3-4,21]。本研究所用的6个分子标记中,Vp1B3等位变异Vp1B3a和Vp1B3b与SGR显著相关(P<0.01);Xwmc468的D片段类型与SGR显著相关(P<0.05);TaSdr-B1和TaMFT-721J与SGR相关不显著;TaPHS1-646与SGR值呈显著相关(P<0.05);TaPHS1-666的NW97S186单倍型与SGR显著相关(P<0.01)。因此,标记Vp1B3、Xwmc468和TaPHS1-646可用于小麦光温敏雄性不育系穗发芽抗性的分子鉴定;KASP标记TaPHS1-666可作为小麦光温敏雄性不育系穗发芽抗性鉴定的参考标记。
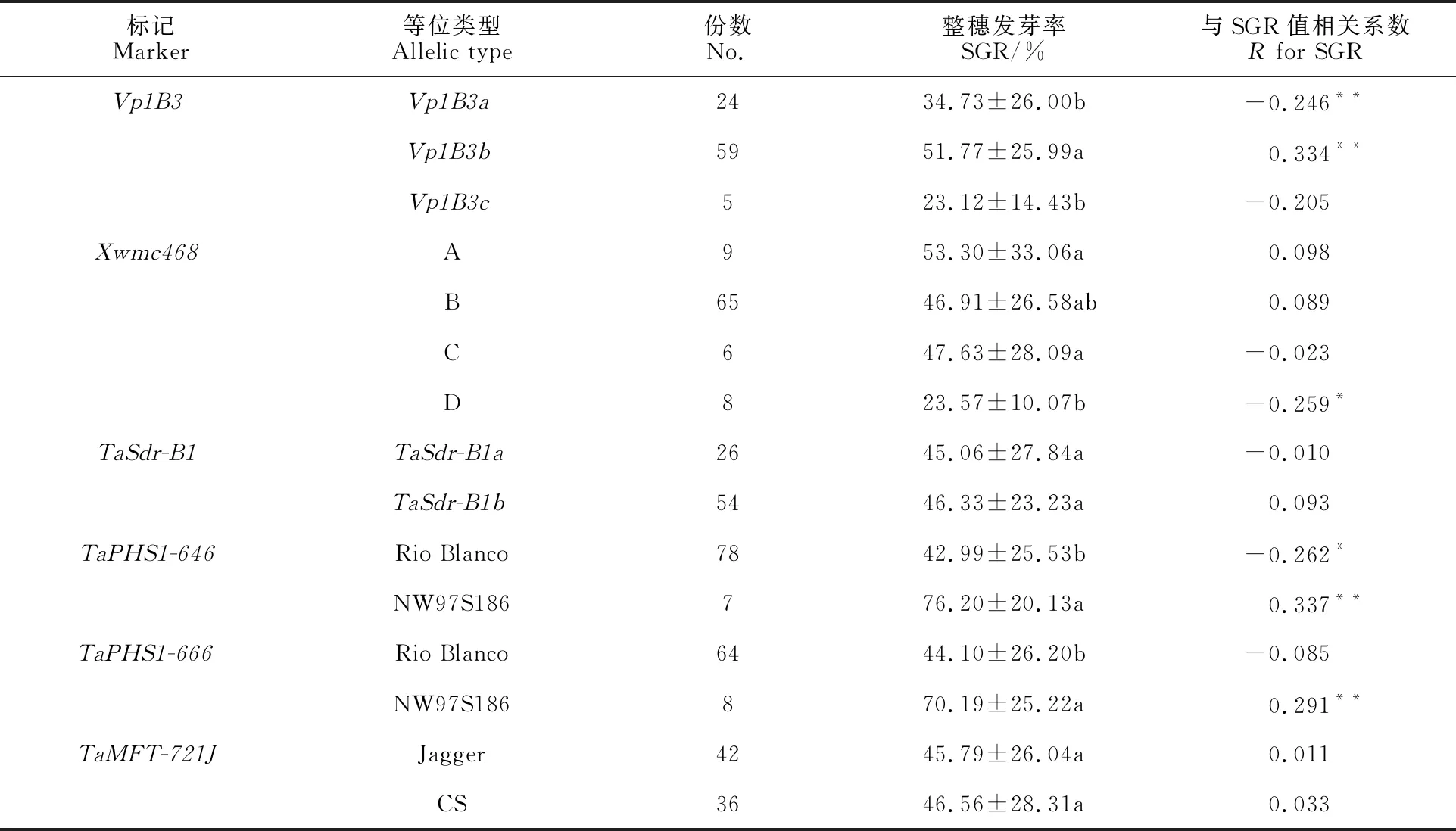
表4 分子标记等位类型与SGR的相关性Table 4 Correlation between alleles and SGR
本研究发现,标记Vp1B3可用于小麦不育系穗发芽抗性鉴定,与前期研究人员的研究结果基本一致[3-4,6,12,14-17,19-25];而杨 燕等[22]发现标记Vp1B3不适合红粒小麦抗性筛选,孙果忠等[26]认为,标记Vp1B3难以用于品种的穗发芽抗性筛选,具体原因有待更多相关研究。本研究发现,标记Xwmc468可用于小麦不育系穗发芽抗性鉴定,与刘光辉等[4],马 丽等[16],杨 燕等[22]研究结果一致。Zhang[8]等发现,TaSdr-B1b基因型的GI值显著高于TaSdr-B1a基因型;Nakamura等[9]发现,TaMFT基因与籽粒休眠呈正相关,而本研究发现,TaSdr基因的两个等位变异的SGR差异不显著,TaMFT-721J与SGR相关不显著,与张维军等[21]的研究结果一致。TaPHS1基因是在白粒小麦中发现的[11]。邹景伟等[18]利用基于TaPHS1基因开发的KASP标记对120份小麦进行了抗穗发芽基因检测,可显著提高小麦育种效率;张维军等[21]发现,分子标记Phs646、Phs666可用于宁夏引黄灌区小麦穗发芽抗性的分子鉴定;而本研究发现,标记TaPHS1-646与SGR显著相关,TaPHS1-666的NW97S186单倍型与SGR显著相关,与前人研究结果不完全一致的原因可能是穗发芽抗性与温度、湿度、激素和穗部性状等因素有关,另外标记对品种和生态区可能具有特异型。
本研究发现,整穗发芽率与抗穗发芽分子标记不完全对应,如BS184的SGR值为4.81%,BS286的SGR值为28.13%,分别属于高抗和中抗穗发芽品系,但6个分子标记均未检测到抗穗发芽基因型,推测这些材料有可能携带有其他未被发现的抗性基因;BS141的SGR值为 84.75%,BS169的SGR值为68.60%,BS111的SGR值为67.15%,都属于高感穗发芽品系,但分子标记TaPHS1-646、TaPHS1-666和Vp1B3的检测结果均为抗穗发芽基因型; BS291的SGR值为34.38%,属于中感穗发芽品系,但分子标记TaPHS1-646、Vp1B3和Xwmc468检测结果均为抗穗发芽基因型。这些结果说明小麦穗发芽抗性是受多基因控制的复杂性状,受自身休眠特性、籽粒颜色、穗部性状和外源激素等因素影响;现有的分子标记都是在特定的生态区和不同材料构建群体开发出来的,对品种具有特异性。因此,小麦穗发芽抗性鉴定要综合利用表型鉴定和分子鉴定,对不同区域不同类型的材料进行分子鉴定时要进行有效性检验,鉴定结果会相对准确。
综合整穗发芽抗性鉴定和分子标记检测结果,筛选出BS212、BS143、BS206、BS129、BS117和BS237共计6份具有较低SGR值的不育系种质资源。在小麦光温敏雄性不育系选育和强优势组合配制中可以对这6份不育系材料加以利用。
猜你喜欢
当代水产(2022年7期)2022-09-20
作物学报(2022年6期)2022-04-08
河南农业·综合版(2022年2期)2022-03-18
河南农业(2022年2期)2022-03-14
中国饲料(2021年17期)2021-11-02
国际医学放射学杂志(2021年5期)2021-10-22
农村实用技术(2020年9期)2020-12-16
猪业科学(2020年6期)2020-12-11
第一财经(2017年36期)2017-09-25
Asian Journal of Urology(2015年3期)2015-12-16
