
不同品种油茶根际丛枝菌根真菌群落结构特征
2020-12-08 08:03林宇岚李正昀张林平谭明曦
林业科学研究 2020年5期
林宇岚,李正昀,吴 斐,裴 蕴,张 扬,张林平,杨 滢,谭明曦
(江西农业大学 鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室,江西 南昌 330045)
油茶(Camellia oleifera Abel.)又称茶子树,属于山茶科、山茶属植物,是我国南方丘陵地区重要的木本油料树种[1]。油茶籽所榨茶油中含有大量不饱和脂肪酸,具有降低血脂、降低胆固醇、保护心血管系统等功效[2]。此外,油茶也是防风固沙和保持水土的重要树种[3]。截止2017 年底,我国油茶种植面积已扩大到443 万多hm2[4]。然而,在油茶林经营过程中,大量不合理施肥已造成生态环境质量下降,引起的土壤重金属含量过高、土壤环境酸化板结等问题日益突出[5]。因此,提高化肥利用效率,降低施用量,恢复退化土壤已成为我国油茶产业可持续发展突出问题。
丛枝菌根(Arbuscular mycorrhizas,AM)真菌,作为一种广泛存在于陆地生态系统的共生菌,能够改善植物对磷、氮等养分的吸收,提高植物对生物和非生物逆境的抗性[6]。随着人们对作物生产中高产、资源高效意识的不断提高,AM 真菌在集约化作物生产体系中的作用日益受到关注[7]。AM 真菌的生理生态功能与其群落结构密切相关,不同的AM 真菌群落通常具有不同的功能[8-9]。植物种类是影响AM 真菌群落结构特征的重要因素[10]。大量研究发现,在物种水平上,同一生境下共存的不同植物,其共生的AM 真菌种类明显不同[11]。少量有关同一植物不同品种AM 真菌群落组成的研究表明,AM 真菌的群落组成受到寄主植物基因型的影响[11]。郭邵霞和刘润进[8]研究发现,牡丹基因型能改变AM 真菌的侵染率和种属组成。Mao 等人[12]发现,不同品种小麦AM 真菌侵染率相同但群落组成不同。近年来,人们不断地筛选和培育出具有优良性状的作物新品种,并应用于生产,因此,开展同一植物不同栽培品种AM 真菌群落组成的研究具有一定的理论研究和实际应用意义。
目前,关于油茶根际AM 真菌种类已有相关报道[5],然而,对不同品种油茶根际AM 真菌的多样性是否存在差异未见报道。为此,本研究以不同品种油茶根际土壤为研究对象, 采用Illumina MiSeq 高通量测序技术研究油茶不同品种AM 真菌多样性,并分析AM 真菌群落特征与土壤理化性质间的关系,为油茶专用菌肥研发和集约化可持续经营提供科学理论依据。
1 材料与方法
1.1 样地和根际土壤采集
油茶根际土壤取自江西省宜春市袁州区江西星火农林公司油茶种植基地(27°43′ N,114°11′E),该基地属典型亚热带季风气候,年降水量1 595.8 mm,年平均气温16.4 ℃,土壤类型为红壤。选取的长林3(CL3)、长林4(CL4)、长林53( CL53)、 赣无84-8( GW84-8)、 赣兴48(GX48)5 个品种均为5 年生油茶,每个品种生态环境条件一致,且经营管理措施相同,种植密度为2 m×3 m。每个品种油茶林中随机划定5 个样地,每个样地随机选取长势相似的油茶5 株。去除土壤表面枯落叶层,在距植物主干50 cm 的东南西北4 个方位,采集0~20 cm 的油茶根系,在无菌自封袋中轻轻抖动根系,收集根系上的土壤,作为根际土,将同一样地5 株植物根际土壤混合均匀,5 个样地作为重复,共计25 个土壤样品,将土样编号装入自封袋运回实验室并分为两份,1 份在-80 ℃条件下冷冻保存,用于DNA 提取及高通量测序;另一份土样避光风干保存,用于测定土壤理化性状。
1.2 土壤理化分析
AM 真菌侵染率测定采用台盼蓝( Trypan blue)染色法[13]和显微十字交叉法[14]进行观察统计;土壤理化性质测定方法具体参照鲁如坤主编《土壤农业化学分析方法》[15],土壤含水率测定采用烘干法;土壤pH 采用电位法;土壤全氮、全磷采用HClO4-H2SO4消化法;铵态氮采用靛酚蓝比色法测定;硝态氮用镀铜镉还原-重氮化偶合比色法测定;有效磷测定采用氟化铵-盐酸浸提法;有机质采用重铬酸钾容量法(外加热法)测定。
1.3 高通量基因组测序方法
油茶根际土壤总DNA 的提取使用土壤基因组DNA 抽 提 专 用 试 剂 盒 ( 购 自MP biomedicals 公司),利用琼脂糖电泳检测DNA 的完整性。采用AMV4.5NF( 5 ′-AAGCTCGTAGTTGAATTTCG-3 ′)/ AMDGR( 5 ′-CCCAACTATCCCTATTAATC AT-3′)引物对AM 真菌rDNA 进行PCR 扩增,利用illumina 高通量测序仪(Illumina MiSeq)测序平台进行测序,下机数据通过QIIME(v1.8.0)剔除问题序列。
1.4 数据分析
采用SPSS 20.0 软件进行数据分析[16],用邓肯检验(Duncan’s test)进行分析和差异显著性检验。采用R 软件,用非度量多维尺度分析(nonmetric multidimensional scaling, NMDS) 检验AM真菌属水平群落组成结构[17];用Mothur[18]软件对属水平的各个分类单元在品种之间的序列量差异进行两两比较检验。使用QIIME[19]软件计算Alpha多样性指数[ 主要包括辛普森指数(simpson)、Chao 1指数、ACE 指数和香农指数(shannon)],并进行多元方差分析和置换检验(Adonis/ Permutational multivariate analysis of variance, PERMANOVA)。利用R 软件的冗余分析(Redundancy analysis, RDA)描述根际AM 真菌丰富度最高的5 个属和土壤因子间的关系。采用Origin Pro 8.5 软件进行绘图(http://www.originlab.com/)。
2 结果与分析
2.1 AM 真菌侵染率
AM 真菌对5 个品种油茶的主要侵染类型均为菌丝侵染,不同品种油茶AM 真菌的侵染率存在差异(图1)。GX48 的总侵染率最高,GW84-8的总侵染率最低。5 个品种的菌丝侵染率变化规律与总侵染率基本一致。CL4 和CL53 的泡囊侵染率高于其他品种。CL4 的丛枝侵染率高于其他品种。
2.2 AM 真菌群落组成
对5 个不同品种油茶根际土壤进行高通量测序,共获得1 376 048 条高质量序列,所有序列共归为2 538 个OTUs,其中CL4、GW84-8、GW48、CL3、CL53 中包含的OTU 数目分别是830、931、1 163、1 103、1 012 个。5 个品种共有OTUs 为176 个,其中不同品种特有OTU 数目分别是237、196、259、306、201 个。总体来看,不同品种油茶根际土壤共有的微生物数量较少。

图1 油茶不同品种根系AM 真菌侵染率Fig. 1 Infection rate of AM fungi in roots of different cultivars of Camellia oleifera
对OTUs 从门到属依次进行分类,总共获得AM 真菌群落1 门、 1 纲、 4 目、 10 科、 12 属(表1),即为球囊菌门Glomeromycota 和球囊菌纲Glomeromycetes, 4 个目分别为球囊霉目Glomerales( 占总数的62.40%)、 类球囊霉目Paraglomerales( 25.32%) 、 原 囊 霉 目Archaeosporales( 4.28%) 、 多 样 孢 囊 霉 目Diversisporales(0.98%);在科水平,5 个油茶品种根际中共鉴定出10 个科, 其中球囊霉科Glomeraceae(占总数的41.12%)、类球囊霉科Paraglomeraceae( 25.32%) 、 近 明 球 囊 霉 科Claroideoglomeraceae(21.28%)等相对丰度较高,而原囊霉科Archaeosporaceae(2.22%)、双型囊霉 科Ambisporaceae( 2.05%) 、 无 梗 囊 霉 科Acaulosporaceae( 0.57%) 、 巨 孢 囊 霉 科Gigasporaceae( 0.33%) 、 多 样 孢 囊 霉 科Diversisporaceae( 0.07%) 、 和 平 囊 霉 科Pacisporaceae( 0.02%) 、 地 管 囊 霉 科Geosiphonaceae(0.01%)等相对丰度较低。
2.3 油茶不同品种AM 真菌属水平分类学组成及丰度
不同品种油茶根际AM 真菌群落组成不同,其中从CL4、CL53 根际土壤中均检测到12 个属,而GW84-8、CL3、GX48 中分别检测到11、10、9 个属(图2)。球囊霉属Glomus、类球囊霉属Paraglomus、巨孢囊霉属Gigaspora 和近明球囊霉属Claroideoglomus 的相对丰度在5 个油茶品种间具有显著差异(图2)。与其他品种相比,GW84-8 中球囊霉属Glomus 相对丰度(71.49%)最高,而类球囊霉属Paraglomus(6.70%)和巨孢囊霉属Gigaspora(0.02%)相对丰度最低;CL53 中近明球囊霉属Claroideoglomus相对丰度(34.12%)最高,而巨孢囊霉属Gigaspora相对丰度(0.18%)最低。

表1 油茶根际土壤AM 真菌群落系统分类 Table 1 Taxonomy of AM fungal communities in rhizosphere soil of Camellia oleifera
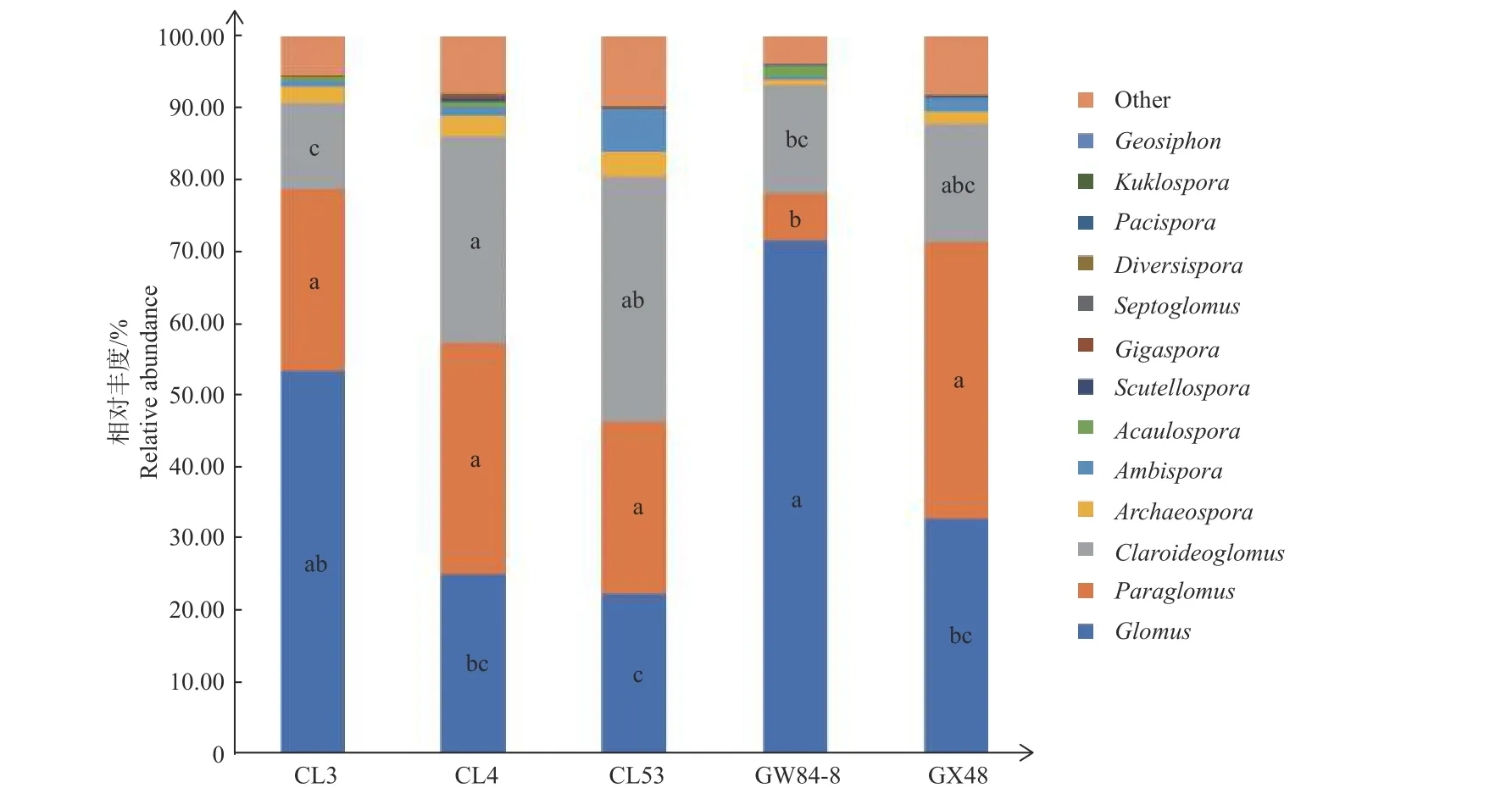
图2 油茶不同品种根际土壤AM 真菌属水平分类单位组成及丰度Fig. 2 Taxonomic composition and distribution abundance of the AM fungal genera in rhizosphere soil of different cultivars of Camellia oleifera
2.4 AM 真菌群落多样性分析
不同品种油茶AM 真菌α 多样性分析结果如表2 所示,5 个品种油茶根际土壤AM 真菌的辛普森指数、Chao 1 指数、ACE 指数和香农指数均无显著差异,说明品种对油茶根际AM 真菌多样性影响不大。
2.5 AM 真菌群落结构的相似性分析
不同品种油茶根际土壤AM 真菌β 多样性分析结果如图3 所示,第一轴从左至右的梯度基本代表品种从CL53 到CL3 排列,代表梯度比较短,第二轴从上至下的梯度代表品种从CL53 到CL4 排列,代表梯度较短。从品种分布来看,GW84-8 和GX48 分布较集中,而CL3、CL4 和CL53 分布较发散,表明CL3、CL4 和CL53 群落结构差异较大,而GW84-8 和GX48 差异较小。Adonis 分析结果表明,油茶5 个品种的AM 真菌属水平上的群落结构存在显著差异(P<0.05)。
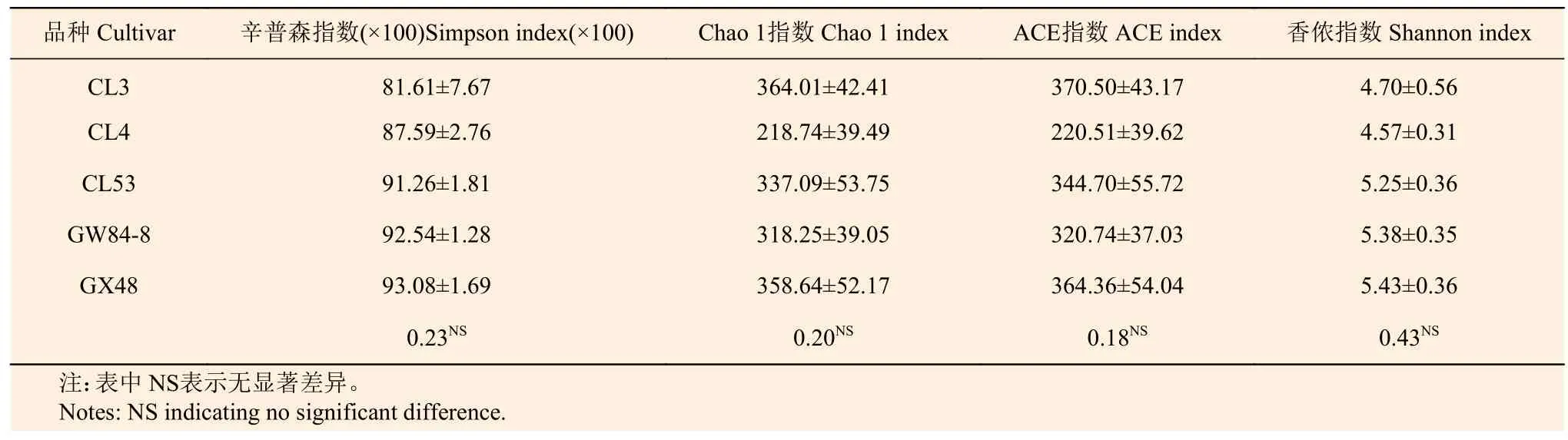
表2 油茶不同品种根际土壤AM 真菌α 多样性指数 Table 2 Alpha diversity index of AM fungi in the rhizosphere soil of different cultivars of Camellia oleifera
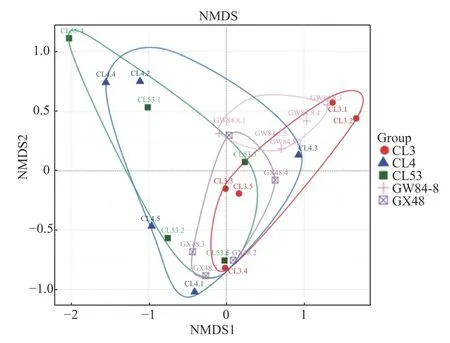
图3 油茶不同品种根际土壤AM 真菌属水平群落结构NMDS 分析Fig. 3 NMDS analysis of community structure of AM fungi in rhizosphere soil of different cultivars of Camellia oleifera
2.6 AM 真菌群落组成与土壤因子的RDA 分析
RDA 分析表明,土壤因子对AM 真菌群落变异的解释量超过30.13%,第一排序轴解释群落变化的16.91%, 而第二排序轴解释群落变化的13.22%(图4)。硝态氮对AM 真菌群落结构有显著影响(P<0.05)。其中,类球囊霉属Paraglomus、巨孢囊霉属Gigaspora、原囊霉属Archaeospora、多样孢囊霉属Diversispora、 无梗囊霉属Acaulospora、隔球囊霉属Septoglomus和地管囊霉属Geosiphon丰度与硝态氮正相关,而双型囊霉属Ambispora丰度与硝态氮负相关。
3 讨论

图4 油茶不同品种根际土壤AM 真菌属水平群落及土壤理化因子的RDA 分析Fig. 4 RDA analysis of genus-level communities and soil properties of AM fungi in rhizosphere soil of different cultivars of Camellia oleifera
地球陆地上绝大多数植物都能与AM 真菌形成菌根,AM 真菌的生长发育和完成生活史离不开寄主植物[20]。本研究结果表明5 个不同品种油茶均能与AM 真菌形成共生体,AM 真菌总侵染率介于20.61%~43.07% 之间,平均侵染率为34.10%,属于中度侵染。各油茶品种间的AM 总侵染率、菌丝侵染率、泡囊侵染率及丛枝侵染率均有显著差异, 表明在本试验系统中, 油茶品种是影响AM 真菌侵染率的重要因素,这与前人的一些研究结论一致,如Steinkellner 等人[21]对10 个品种番茄的AM 侵染率进行研究,发现番茄品种显著影响AM 真菌的侵染率;郭绍霞等[8]研究表明牡丹品种会影响到AM 真菌侵染率和种属构成。同一植物不同品种的生理生化特性不同,根系生理代谢和分泌物组成含量的差异可能影响AM 真菌对植物根系的识别和侵染强度[12]。
运用高通量测序技术对5 个不同品种油茶根际土壤AM 真菌进行检测,结果表明油茶根际土壤共有AM 真菌12 个属,相对丰度前3 的属分别为球囊霉属Glomus( 41.053%)、 类球囊霉属Paraglomus( 25.323%) 、 近 明 球 囊 霉 属Claroideoglomus(21.276%),其中Glomus 属是油茶林AM 真菌中分布范围最广的属,这与邓小军等[5]、苏琍英等[22]研究结论一致;而刘辉等[23]在茶树中共分离鉴定出8 属36 种AM 真菌,其中Acaulospora、Glomus、Funneliformis 为 丰 度 较 高的前三个属,上述结果表明AM 真菌对寄主植物有一定的偏好性。本研究中,5 个品种油茶根际分别鉴定出9~12 个属AM 真菌,且AM 真菌的辛普森指数、Chao 1 指数、ACE 指数和香农指数均无显著差异,表明品种对油茶根际土壤AM 真菌丰富度和多样性的影响不明显; 由于Glomus、Paraglomus、Gigaspora 和Claroideoglomus 等属在不同品种中的相对丰度存在差异,因此造成了不同品种油茶的AM 真菌群落结构的变化,与郭邵霞等[8]、Mao 等人[12]研究结果一致。植物种类是影响AM 真菌群落结构的重要的因子[24],不同植物和同一植物不同品种遗传物质并不相同,可能由于根系形态结构、分泌物等差异,能促进或抑制某些AM 真菌的生长发育,进而影响到植物AM 真菌群落结构特征。
在陆地生态系统中,土壤类型、土壤肥力、土壤pH 值等土壤因子都会对AM 真菌群落组成和分布有重要影响[25-28]。Matevž等人[29]认为有效磷是影响葡萄园AM 真菌群落变化的主要原因之一;Guo 等人[30]认为土壤pH 是影响AM 真菌多样性的最主要土壤因子,可能是由于其能够直接影响AM真菌生理状态,改变其生态位,并通过调节土壤养分的生物有效性等间接影响AM 真菌群落;而邓小军等[5]在研究中发现油茶AM 真菌丰度与全氮、有机质显著正相关。在本研究中,我们发现硝态氮是影响油茶根际AM 真菌群落组成与分布的最主要因素,这可能是因为硝态氮在土壤中移动性较强,能够通过质流被植物吸收,并改变土壤养分平衡和时空特性,从而间接地影响AM 真菌群落[31]。AM 真菌的群落组成是其生理生态功能的基础[9],这些AM 真菌群落结构的差异是否影响其功能还有待进一步研究。
4 结论
AM 真菌是陆地生态系统中重要的微生物之一,本研究研究了5 个不同品种油茶根际AM 真菌群落的多样性及其群落结构特征,结果表明,油茶根际土壤中AM 真菌资源较为丰富,5 个不同品种油茶根际土壤AM 真菌多样性无显著差异,而油茶品种对AM 真菌群落结构有显著影响,且硝态氮是影响油茶根际AM 真菌群落组成和分布的主要土壤因子。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
中国土壤与肥料(2021年5期)2021-12-02
中国瓜菜(2020年8期)2020-09-26
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
中国西部(2017年4期)2017-04-26
湖南农业(2017年1期)2017-03-20
中国蔬菜(2016年8期)2017-01-15
