
贵州省西北部马尾松人工林种群数量特征与动态
2020-12-08 00:28:22陈群利薛晓辉李望军
中南林业科技大学学报 2020年11期
何 斌,李 青,陈群利,薛晓辉,李望军
(贵州工程应用技术学院 a.贵州省普通高等学校生物资源开发与生态修复特色重点实验室;b.生态工程学院,贵州 毕节 551700)
植物种群是特定时间内占据一定空间的同种植物个体的总和,由物种生物学特性和生境条件共同决定,植物的不同生活史阶段会影响种群的数量动态变化[1]。种群结构和动态是种群生态学研究的热点问题之一[2],种群动态是研究特定生境条件下种群的变化规律,种群结构是研究特定时间内种群的空间配置状况[3]。研究植物种群结构与动态不仅可以反映种群的生存状态,揭示植物与环境之间的适合度,而且对保护和利用植物资源具有重要意义[4-5]。种群统计是研究种群数量动态的基本方法,可以筛选出影响植物生长发育的环境因子,预测种群未来的发展趋势[6-7],为种群的管理和利用提供理论指导[8]。目前,常用的种群统计方法有静态生命表、生存分析和时间序列预测等,已被广泛应用到灌木、乔木和濒危、稀有树种的研究中。通过这些方法研究植物种群结构与数量动态有助于人们更好地认识种群的生态特性、形成机制、更新策略与演替规律,对种群及群落的更新复壮、生态系统的恢复与发展、预测种群的演化趋势具有重要的理论意义和应用价值[8-10]。不少学者运用种群统计学方法开展了种群数量特征与动态方面的研究,如朱强等[11]对鄂西南川陕鹅耳枥种群研究表明,其属于增长型种群,存活曲线趋于Deevey-Ⅲ型;谢春平等[12]对五指山地区海南苏铁种群研究表明,其存活曲线趋近于Deevey-Ⅱ型,属于稳定型种群;刘海燕等[13]对稀有濒危植物长柱红山茶种群研究表明,其生存曲线基本属于Deevey-I 型,属于衰退型种群等等。可见,不同种群的数量特征不尽相同。
马尾松P.massoniana分布范围极广,适应能力强,耐干旱瘠薄,是中国长江流域主要的荒山造林和生态建设树种之一,也是该区域植被演替的先锋植物。中国以贵州为中心的西南地区是世界上面积最大、最集中连片的喀斯特区域,其独特的地质背景和不合理的土地利用,导致了土壤质量退化、生态环境质量恶化等严重的石漠化现象,严重制约了地方经济和社会的可持续发展,甚至威胁到了长江、珠江流域的生态安全。近年来,党中央和地方政府高度重视喀斯特地区植被的恢复和重建。马尾松作为先锋树种是喀斯特地区植被恢复和生态建设的关键,掌握该区域马尾松的生存和发展状况十分必要。迄今为止,国内外有关马尾松种群动态的研究很多,董鸣[14]对缙云山马尾松种群数量动态进行了定量分析,方炜等[15]研究了华南地区的马尾松种群动态,李根前等[16]对陕南的马尾松种群结构进行了初步研究,熊能等[17]定量分析了千岛湖姥山马尾松种群结构,万志兵等[18]研究了黄山市稽灵山次生马尾松种数量动态,吴语嫣等[1]定量分析了长汀水土流失区侵蚀劣地马尾松种群动态,但尚未见到有关西南喀斯特地区马尾松种群数量的相关研究。因此,分析喀斯特地区马尾松种群数量特征及动态变化,了解其现存状态并提出相应的经营管理措施迫在眉睫。
鉴于此,以分布于贵州省西北部的马尾松为研究对象,用“空间代替时间”的方法,从种群的年龄结构、静态生态表、生存分析以及时间序列预测等方面定量分析马尾松种群的生存现状和数量动态变化,阐明喀斯特地区马尾松种群结构和动态变化规律,以期为马尾松种群的有效管理和利用提出合理建议,进而为喀斯特地区植被的恢复和重建提供理论指导和决策依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于贵州省西北部的毕节市黔西县境内(26°45′~27°21′N,105°47′~106°26′E),为典型的喀斯特地区,平均海拔1 250 m,具有低纬度、高海拔的特点。研究区属于中亚热带湿润季风气候,夏无酷暑、冬无严寒,雨量充沛,年平均气温13.8 ℃,年平均降水量1 050 mm,年平均日照1 348.9 h。地貌复杂多样,以低中山、丘陵为主;土壤类型多样,主要有黄壤、石灰土、紫色土、水稻土和潮土。森林覆盖率52.4%,主要有马尾松、云南松、杉类、栎类等树种。
1.2 研究方法
1.2.1 样地设置与调查
在对马尾松人工林全面踏查的基础上,于2018年7月在马尾松人工林的集中分布区黔西县中坪镇,采用20 m×30 m 的典型样地调查取样,共设置9 块样地,每块样地划分为24 个5 m×5 m的小样方,对样方内的乔木每木检尺,测量胸径、树高、活枝下高和冠幅等,同时记录海拔、坡度、坡向等基本信息。
1.2.2 种群结构划分
根据相关研究,相似环境条件下同种植物的龄级和径级具有一致的反应规律[19]。因此,采用“径级代替龄级”的方法分析种群结构。根据马尾松的生活史特点和样地调查情况,按照上限排外法,每4 cm 划分为1 个等级,将胸径分为12 个等级,其中Ⅰ级为BDH<2.5 cm、Ⅱ级为2.5 cm≤BDH<5 cm 和Ⅻ级为DBH≥41 cm;从高度0 m开始,每增加3 m 划分为1 个等级,将马尾松高度划分为10 个等级,其中Ⅹ级为H≥27 m。用经过树冠中心的2 个垂直直径的乘积来表示冠幅,将马尾松冠幅划分为9 个等级:Ⅰ级(C<1 m2)、Ⅱ级(1 m2≤C<5 m2)、Ⅲ级(5 m2≤C<10 m2)、Ⅳ级(10 m2≤C<15 m2)、Ⅴ级(15 m2≤C<20 m2)、Ⅵ级(20 m2≤C<25 m2)、Ⅶ级(25 m2≤C<30 m2)、Ⅷ级(30 m2≤C<35 m2)和Ⅸ级(C≥35 m2)。统计各等级的马尾松株数,绘制马尾松种群径级、高度级和冠幅级的结构图。
1.2.3 种群数量动态量化
参考陈晓德[20]和姜在民等[2]的种群动态量化分析方法,定量描述马尾松种群动态,具体计算公式如下:

式中:Vn代表种群从n到n+1 级的个体数量变化;VPi代表整个种群年龄结构的数量变化动态指数;V′Pi代表种群在外部干扰下年龄结构的数量变化动态指数;Sn、Sn+1分别为第n和n+1 级种群个体数;k为种群大小级数量。当Vn、VPi、V′Pi分别为正、0、负值时,反映种群个体数量的增长、稳定和衰退变化趋势。
1.2.4 静态生命表和生存分析
根据静态生命表的编制原理,统计不同龄级马尾松的个体数量,编制马尾松种群静态生命表,具体参数参考黄冬柳等[21]的方法。为了避免编制过程中死亡率出现负值的情况,采用方程拟合法对各龄级的实际个体数进行匀滑处理。同时,引进4 个生存函数分析马尾松种群的生存规律,具体计算公式如下:

式中:Si为存活率;hi为龄级宽度。
1.2.5 时间序列分析
采用一次移动平均法[1-2]模拟和预测未来马尾松种群的数量动态,具体公示如下:

式中:n表示预测的龄级时间,t为龄级,Xk为k龄级内的个体数量,表示未来经过n个龄级后t龄级的个体数量。预测了马尾松未来经过2、4、6、8 和10 龄级后各龄级的个体数量。
2 结果与分析
2.1 种群结构特征
已有研究表明,可以用种群的径级结构代替年龄结构来反映种群的生存状态[22]。本研究中,马尾松种群径级结构呈不规则的倒“J”型(图1),Ⅰ径级的个体数相对较少,占总数的3.29%;Ⅱ~Ⅳ径级的个体数最多,占总数的55.42%;Ⅲ径级植株数达到峰值,占总数的22.09%;Ⅴ~Ⅸ径级的个体数次之,占总数的35.94%;Ⅸ径级以上的老树最少,仅占总数的5.35%;中老径级个体数呈逐渐下降趋势,由Ⅴ径级个体数占比的12.76%逐渐降低到Ⅻ径级个体数占比的1.51%。说明该地区马尾松中幼龄级个体数充足,具有较强的自我更新能力,属于增长型种群。
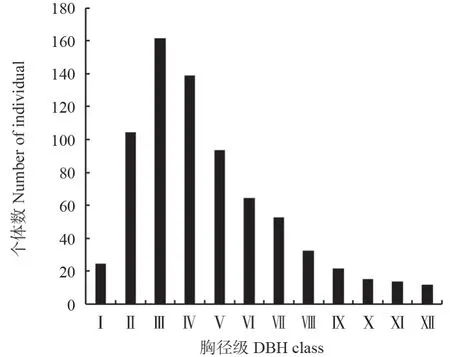
图1 马尾松种群径级结构Fig.1 Diameter class structure of P.massoniana population
植物种群高度能够客观地反映不同种群个体在垂直结构中的地位,揭示种群的演替趋势[23]。图2为马尾松种群的高度结构,从图2中可以看出,Ⅰ级个体数最少,仅占总数的1.10%,随后Ⅱ~Ⅶ级个体数逐渐上升,占比从5.91%升高到18.68%,此后,Ⅷ~Ⅹ级个体数又逐渐下降。总体上看,马尾松种群高度结构呈不规则的单峰曲线,Ⅵ~Ⅷ级个体数占总数的比例最大,达到了50.14%(图2)。
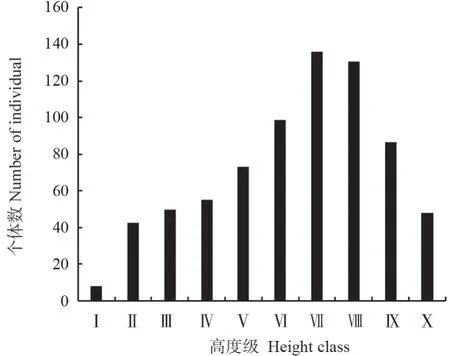
图 2 马尾松种群高度结构Fig.2 Height class structure of P.massoniana population
冠幅是反映植株树冠的重要参数,一定程度上说明了树木占据空间的能力和竞争力[24-25]。从马尾松种群的冠幅结构(图3)可以看出,Ⅱ级冠幅的马尾松个体数最多,占总数的34.71%;随着冠幅的增加马尾松个体数迅速下降,Ⅷ级冠幅的个体数最少,仅占总数的2.89%;Ⅸ级冠幅个体数又略有上升。总体上,马尾松种群冠幅结构呈非典型的倒“J”型,Ⅱ~Ⅲ级冠幅的马尾松个体数最多,占总数的55.42%,即冠幅主要集中在1~10 m2。
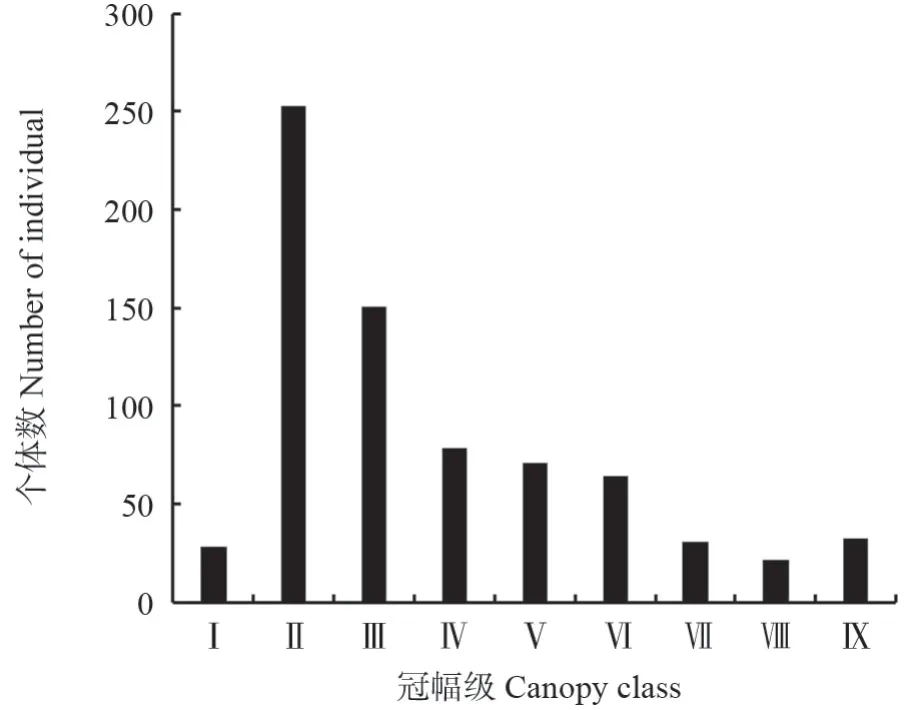
图3 马尾松种群冠幅结构Fig.3 Crown width class structure of P.massoniana population
种群动态量化方法可以对种群结构特征进行更客观的评价,表1为马尾松种群个体数量动态变化。由表1可以看出,马尾松种群相邻各龄级间个体数量均有明显波动,且存在差异;马尾松种群V1~V2均小于0,说明Ⅰ~Ⅱ龄级呈衰退的结构动态关系;V3~V11均大于0,说明Ⅲ~Ⅺ龄级呈增长的结构动态关系。当不考虑种群受外界环境干扰时,马尾松种群结构数量动态变化指数VPi=13.21>0;当考虑种群受外部随机干扰时,马尾松种群结构数量动态变化指数V′Pi=0.10>0,但趋近于零,说明马尾松种群为增长型种群。随机干扰风险极大值P极大=0.76,说明马尾松种群对外界随机干扰不敏感。
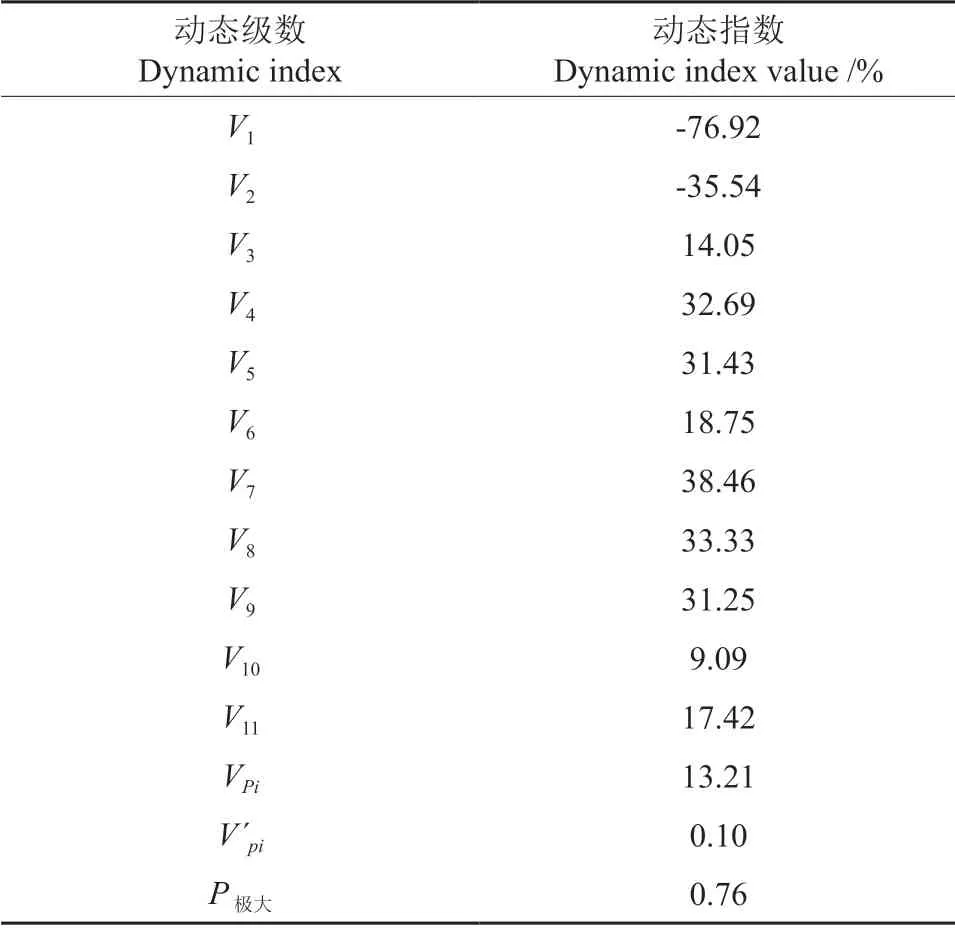
表1 马尾松种群动态变化指数Table 1 Dynamic indices of P.massoniana population
2.2 静态生命表及存活曲线
由表2可知,随着林龄的增加,马尾松种群的死亡率和消失率尽管有小幅波动,但整体上表现为先降低后升高的趋势。由于环境的筛选作用对Ⅰ龄级个体的存活影响比较大,Ⅰ~Ⅳ龄级的死亡率和消失率呈降低趋势;随着林龄的增长,植株个体的生长发育需要更大的空间和更多的养分,种间和种内竞争越来越激烈,Ⅳ~Ⅹ龄级的死亡率和消失率呈升高的趋势。从生命期望值(ex)来看,随着林龄的增加,马尾松种群的生命期望值总体上呈下降趋势,这可能与马尾松的生物学特性有关。Ⅱ~Ⅳ龄级生命期望值较高,说明该阶段群落内竞争比较小,生存环境较好,马尾松种群的生理活动比较旺盛;Ⅹ~Ⅻ龄级生命期望值较低,说明该阶段马尾松种群已经进入了生理衰退期。
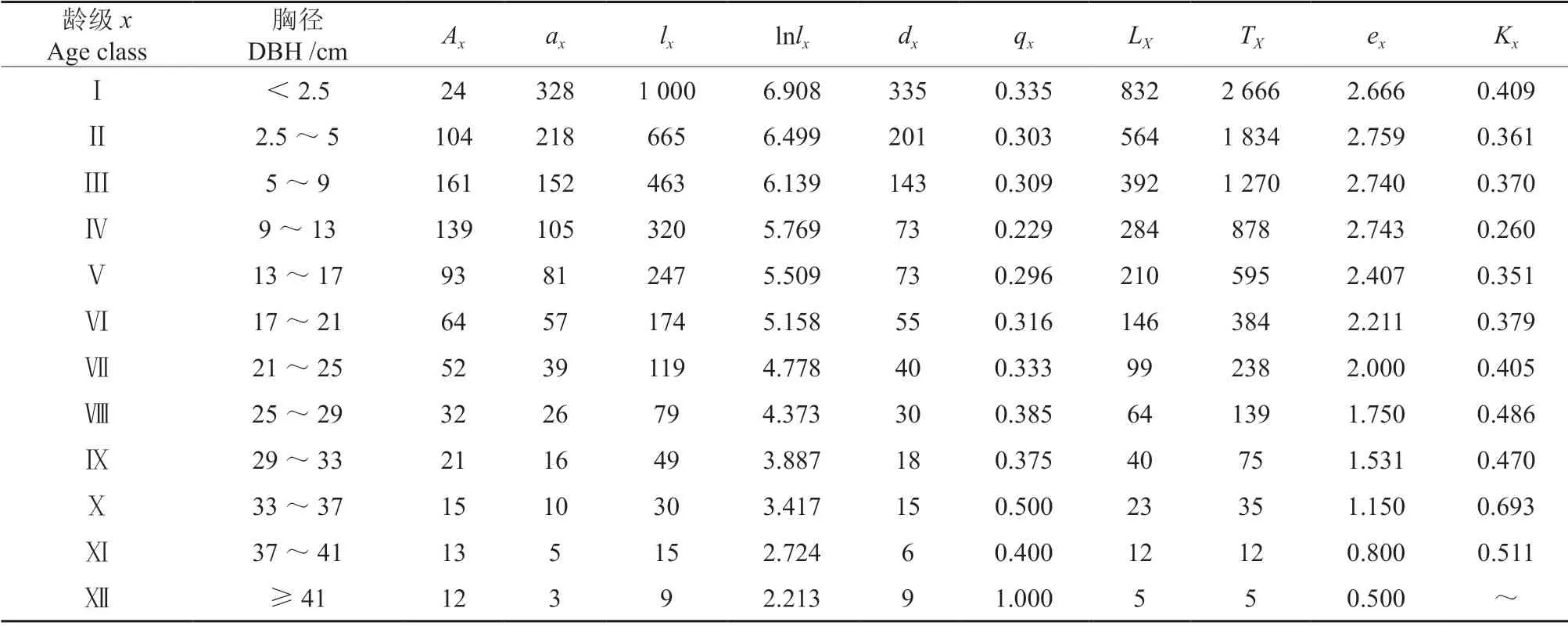
表2 马尾松种群静态生命表Table 2 Static life table of P.massoniana population
图4为马尾松种群存活曲线,从图4中可以看出,马尾松种群的存活率整体呈下降趋势。根据Deevey 对存活曲线的划分,马尾松种群的存活曲线介于Deevey-Ⅱ型和Deevey-Ⅲ型之间之间。应用Hett 和Loucks 的数学模型检验马尾松种群的存活曲线,建立了如下模型:Nx=8.412 1e-0.095x(R2=0.927 9),Nx=8.815 2x-0.399(R2=0.718 6)。由于幂函数模型的R2值(0.718 6)小于指数函数模型的R2值(0.927 9),因此,马尾松种群的存活曲线更趋于Deevey-Ⅱ型。根据戈峰[26]的研究,Deevey-Ⅱ型可分为3 个亚型,马尾松种群存活曲线的直线拟合模型为:y=-0.412 1x+7.459 8(R2=0.986 6)。综上分析,马尾松种群存活曲线属于Deevey-Ⅱ型中的B1亚型。
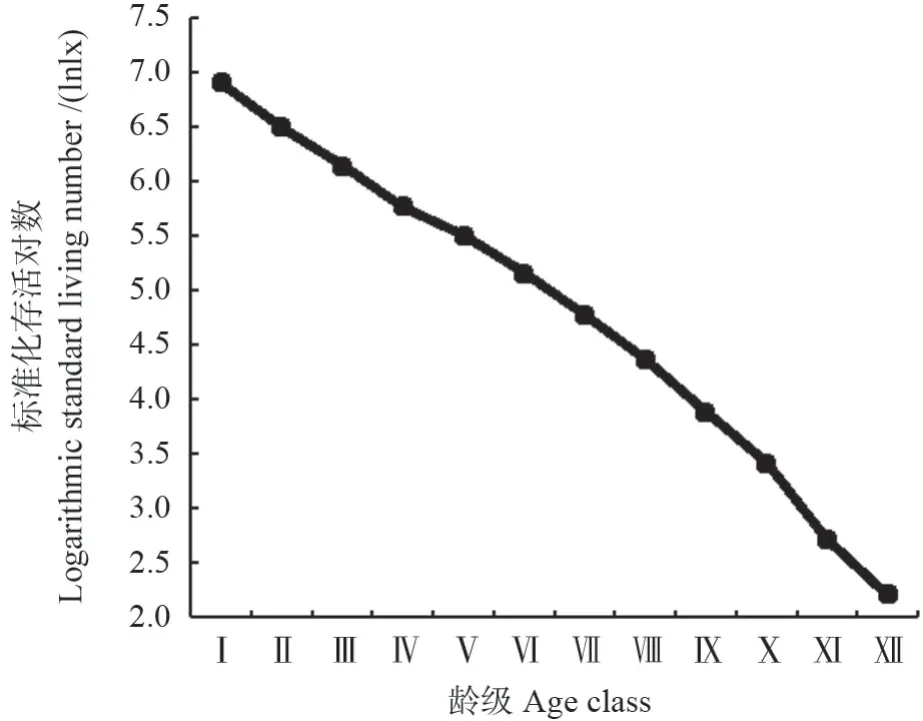
图4 马尾松种群存活曲线Fig.4 Survival curve of P.massoniana population
图5为马尾松种群的死亡率和消失率曲线。从图5中可已看出,尽管死亡率与消失率的变化幅度有差别,但是总的变化趋势基本一致。死亡率和消失率的第一个峰值出现在Ⅰ龄,分别为0.335 和0.409,这可能是由于幼龄阶段喀斯特地区土壤薄、养分贫瘠等严酷的生境条件以及种内、种间竞争导致的幼树个体大量死亡;此后,死亡率和消失率波动下降,在Ⅳ龄级达到最低点,分别为0.229 和0.260,说明马尾松经历了幼林期的环境筛选和竞争后,适应能力增强;随后死亡率和消失率缓慢上升,在Ⅹ龄级时达到最大值,分别为0.500 和0.693,说明此时基本上达到了生理寿命。

图5 马尾松种群死亡率和消失率曲线Fig.5 Mortality and vanish rate of P.massoniana population
2.3 生存分析
从马尾松种群生存分析函数曲线(图6和图7)可知,随着林龄的增加,马尾松生存率和累计死亡率呈单调下降和上升趋势,在Ⅱ龄级前达到平衡,两者存在互补现象。马尾松生存率和累计死亡率在Ⅰ~Ⅲ龄级升降幅度较大,生存率急速下降,累计死亡率急速上升;在Ⅹ龄级以后,生存率和累计死亡率变化趋于平缓,此时生存率为1.5%,累计死亡率为98.5%,种群表现出稳定状态,说明马尾松在该龄级以后逐渐进入生理衰退期。由图7可知,马尾松的死亡密度曲线Ⅳ龄级前急剧下降,此后缓慢下降,总体表现为前期>中期>后期。危险率在Ⅰ龄级出现第一次峰值,为16.1%,随后急剧下降,在Ⅳ龄级达到最低值,为6.5%,Ⅳ龄级到Ⅹ龄级,危险率缓慢波动上升,在Ⅹ龄级出现峰值(16.7%)后下降,随后危险率急速上升,最后达到50%。综合4 个生存函数的分析结果,马尾松种群具有前期锐减、中期稳定、后期衰退的动态变化特征。
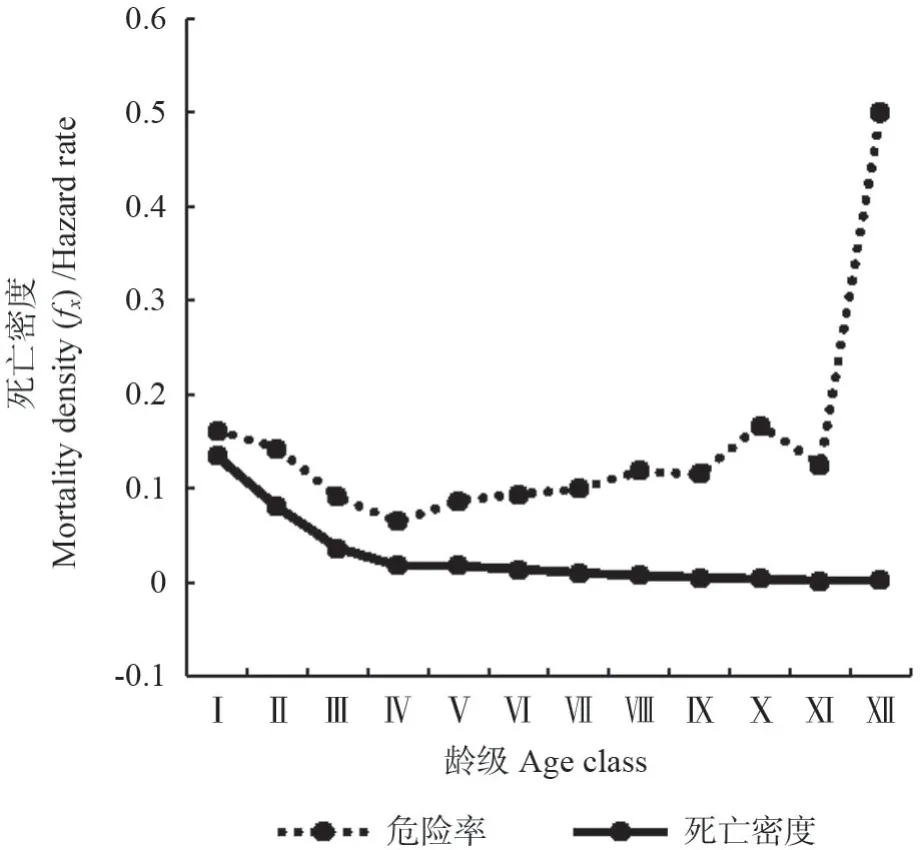
图6 马尾松种群死亡密度和危险率曲线Fig.6 Mortality density and hazard rate of P.massoniana population
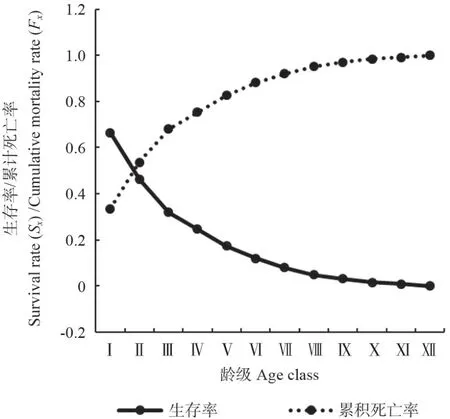
图7 马尾松种群生存率和累计死亡率曲线Fig.7 Survival rate and cumulative mortality rate of P.massoniana population
2.4 时间序列分析
根据时间序列预测模型,应用一次移动平均法预测马尾松种群未来经过2、4、6、8 和10 个龄级时间后数量变化(表3)。由表3中可知,马尾松种群经过2、4、6 龄级后,Ⅶ~Ⅻ龄级个体数表现出不同程度的增加趋势;经过8 和10 龄级后,Ⅺ~Ⅻ龄级个体数也表现出增加趋势,说明随着时间的推移,马尾松种群幼龄个体在一定程度上可以补充中老龄级个体,保证马尾松种群的增长。但是,马尾松种群经过2 龄级后,Ⅱ~Ⅲ龄级个体数分别从104 株和161 株下降到64 株和133 株,分别降低了38.46%和17.39%,说明由于马尾松幼苗的不足,幼树数量将会有较大幅度的减少。综合其总体发展趋势来看,可以推测,如果不采取适当的抚育措施,随着时间的推移,马尾松种群由于幼树更新不良,未来将难以维持,趋于衰退。
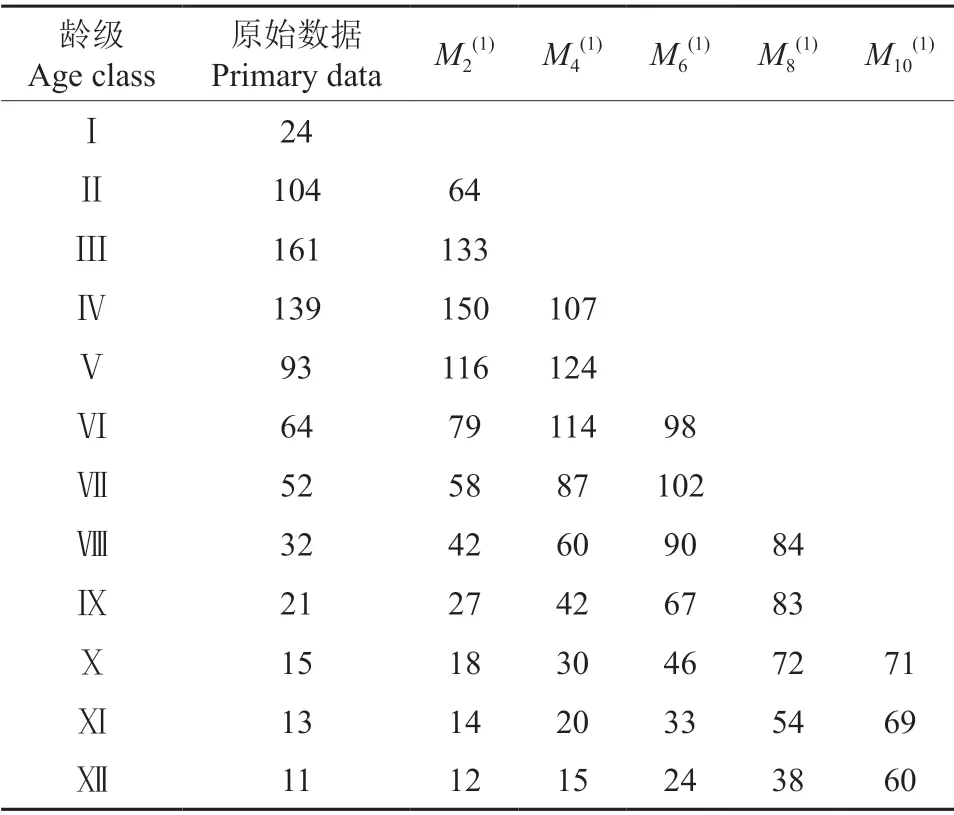
表3 马尾松种群的时间序列分析†Table 3 Time sequence analysis of P.massoniana population
3 结论与讨论
3.1 马尾松种群的结构特征
植物种群结构是种群发育过程中内部个体的表达,受生物因素和非生物因素的共同影响,反映了植物对环境的适应性和立地条件的优劣[2]。植物种群结构分析有助于推测种群的动态变化过程及其潜在的生态学机制,揭示种群生存现状和更新策略[27]。本研究调查发现,样地内马尾松种群的径级结构呈不规则的倒“J”型,属于增长型种群,有向稳定型转化的趋势,说明马尾松种群适应当地的环境条件,并能最大化的实现自身生存扩展。但是,Ⅰ、Ⅱ龄级个体数少于Ⅲ龄级个体数,说明马尾松种群幼龄个体无法充分补充中老龄个体,种群的长期稳定性难以维持,这与一些濒危植物种群特征相似[8]。马尾松天然更新能力强,能在养分贫瘠的土壤中萌发生长,且马尾松喜光,样地内低的覆盖度有利于马尾松种子发芽,从而使得幼苗个体数量相对丰富;但是由于幼苗个体竞争力较弱,强烈的环境筛选作用导致马尾松幼苗死亡率高,仅有部分幼苗能进入到下一阶段,种群更新过程被延缓。中龄林阶段,马尾松种群又会因为种内竞争的自疏作用和外部的人为干扰等影响种群的正常增长。吴语嫣等[1]对长汀水土流失区侵蚀劣地马尾松种群动态分析后认为马尾松种群为基部极宽顶部狭窄的金字塔形结构,属增长型种群;程寅瑞等[28]对南京南郊马尾松种群动态分析后认为马尾松种群年龄结构呈明显的纺锤形,属典型的衰退型种群。造成这种差异的原因可能是调查研究区域的生境条件、龄级划分标准和研究方法不同造成的。
3.2 马尾松种群的动态变化趋势
植物种群动态是个体生存能力与外界环境相互作用的结果[29]。静态生命表是反映植物种群发展趋势的重要指标,可以揭示种群与环境的竞争关系[30]。从静态生命表可以看出,马尾松种群经历了3 次死亡高峰。第一次死亡高峰发生在Ⅰ龄级,这一时期幼苗较多,抵抗外界环境的能力较弱,同时需要争夺水分和养分来满足生存,幼苗以高死亡率为代价来应对强烈的环境筛选和种内竞争[31];第二次死亡高峰发生在Ⅹ龄级,经过环境筛选后存活的马尾松,生存能力增强,死亡率下降,种群进入了平稳发展时期,但是,随着马尾松的生长,有限的生存空间不能满足马尾松个体日益增长的需求,种群内部竞争加剧,密度制约效应开始发挥作用,种群第二次出现死亡高峰;第三次死亡高峰发生在Ⅻ龄级,此时死亡率达到了100%,但是从整个种群的繁衍来看,老龄树的死亡为马尾松的中幼龄树占领有限的空间资源提供了契机。本研究认为马尾松种群属于Deevey-Ⅱ型,这与吴语嫣等[1]的研究结果相同,但程寅瑞等[28]认为南京南郊马尾松种群属于Deevey-I 型。费世民等[32]研究认为同一树种的存活曲线在不同研究区域可能不同,造成这种现象的主要因素可能是环境因子。生存函数是任意时刻的函数,更能直观地反映种群的结构和动态变化,有很高的实际应用价值[33]。生存分析表明,马尾松种群生存率和累计死亡率单调下降和上升,二者互补;整体表现为前期锐减、中期稳定、后期衰退的动态特征,这和径级结构、静态生命表的分析结果一致。从种群数量动态指数可以看出,从Ⅲ龄级开始各动态指数均大于0,说明马尾松种群能够很好地适应当地的生存环境。由于VPi=13.21>0,说明马尾松种群为增长型种群;V′Pi=0.10>0,但趋近于零,有趋向于稳定的种群特征,说明马尾松种群有从增长向稳定的变化趋势;P极大=0.76,表明马尾松种群不易受到环境干扰,对外界干扰不敏感。时间序列分析是预测种群未来发展趋势的重要手段[34]。由时间序列分析可知,在未来2、4、6、8 和10 个龄级时间后马尾松老龄株树逐渐增多,幼龄株树降低,会影响该种群未来的生存和发展。
3.3 马尾松种群的抚育与经营对策
植物种群数量的减少主要是由以下两方面原因:一是幼苗供给不足,二是成活个体的死亡[35]。以上分析结果表明,马尾松种群呈不规则的倒“J”型结构,植株个体主要集中在Ⅱ~Ⅳ龄级,只有少部分个体能够成功进入下一龄级;中龄林阶段,植株个体占据的生存空间和营养资源可以满足其生长发育需求,种群数量趋于稳定;老龄林阶段,马尾松进入生理死亡年龄,种群逐渐衰退。因此,要保证马尾松种群的可持续发展必须针对不同阶段的生长发育特点采取不同的抚育措施。幼龄林阶段,对密度较大的马尾松林进行间伐、清理林下活地被物等措施,为马尾松幼苗的生长发育创造有利的生境,提高存活率;中龄林阶段,通过疏伐、施肥等措施,降低种内个体间对养分和生存空间的竞争,促使其茁壮生长,提高种子质量;老林龄阶段,及时将枯死木、病倒木移出林地,避免病虫害的发生。同时,要加强宣传教育活动,减少当地群众对马尾松的砍伐。
由于受实验条件、样地数量等客观因素的限制,本研究只依据径级结构对贵州省西北部马尾松人工林的种群数量和动态进行了初步研究,而对于种群在空间上的分布格局以及种群与生境、人为干扰之间的相互关系均未涉及,这些内容都有待将来进一步深入研究。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
丽水学院学报(2022年2期)2022-04-19 01:22:10
广西林业科学(2021年6期)2022-01-21 07:39:10
林业科技情报(2021年3期)2021-09-01 02:01:26
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
中南林业调查规划(2017年1期)2017-12-19 00:58:07
