
缠绕植物的向性缠绕方向研究综述与数学表述
2020-12-05 07:14:32王浩
生物进化 2020年3期
王 浩
缠绕植物属攀缘植物中的一类, 多数情况下指具有柔软茎、 可以攀附在其他植物茎上螺旋生长的草本植物。调查发现,不同缠绕植物的缠绕方向具有严格的种属特异性。对于某种植物而言,其自然缠绕方向是固定的,不取决于外界条件的作用, 尽管有时特定的缠绕方向不利于植物生长。缠绕植物的缠绕方向可能属于一种遗传上较为保守、起源较早的性状,这将在后面加以论述。本文将通过调查和总结多种缠绕植物的缠绕方向,进行这些植物起源地的推测,并对缠绕性状的起源做出理论上的解释。
调查发现,近90%的缠绕植物茎的向性生长方向为右旋, 这些植物不仅茎可表现出特定向性缠绕,在植物体的其他器官上,有时也可见特定的旋转方向。实验证实,多数情况下,植物的缠绕方向与外界条件无关, 外界条件包含科里奥利力、太阳辐射角等。虽然外界条件无法改变植物的缠绕方向, 但是外界条件很有可能是缠绕植物进化早期选择缠绕方向的决定性作用。调查发现,来自北半球的植物多为右旋,来自南半球的植物多为左旋。可推测,外界条件在进化早期对植物缠绕方向的分化的影响可能通过影响生长素(β-吲哚乙酸)的分布实现。随后推测, 植物茎内可能存在一种定向横向运输生长素的结构, 使得植物的缠绕方向不易受外界条件而改变。目前学界普遍赞同的假说有以下两种。
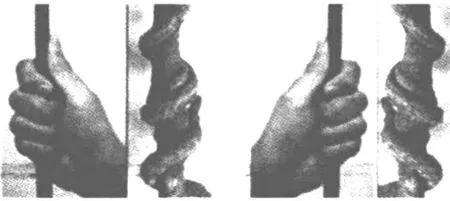
图1 缠绕植物的手性
假说一:太阳辐射角假说
以北半球为例来说明太阳辐射角对植物缠绕方向的影响。如图2,在北半球,太阳运动方向并非是自西向东, 在不同季节太阳运转方向不同,但运行轨道均偏向南侧,这导致了进化早期植物的特定的缠绕方向。如图3,植物茎在向上生长的同时,茎尖胚芽鞘部位可能受光照刺激定向运输生长素至远心端,导致远心端生长素浓度稍高于近心端, 远心端细胞拉长,由于太阳方向始终在发生变化,即向性刺激的方向始终在变化,但刺激的运动方向却始终不变,导致植物在向上生长的同时表现出逆时针生长的现象(俯视),宏观上看表现为右旋生长。
假说二:科里奥利力假说
另一理论分析了地球自转产生的向心力即科里奥利力作用对植物生长素分布继而对植物缠绕方向的影响。地球为一自转的非惯性系,只要物体运动方向不与地球自转轴平行, 则受科里奥利力作用。生长素产生于茎尖生长点处,产生后向形态学下部运输。运输过程中可能受科里奥利力影响使生长素在同一横截面处近心端和远心端浓度不同,从而远心端细胞拉长,导致旋转现象的发生。
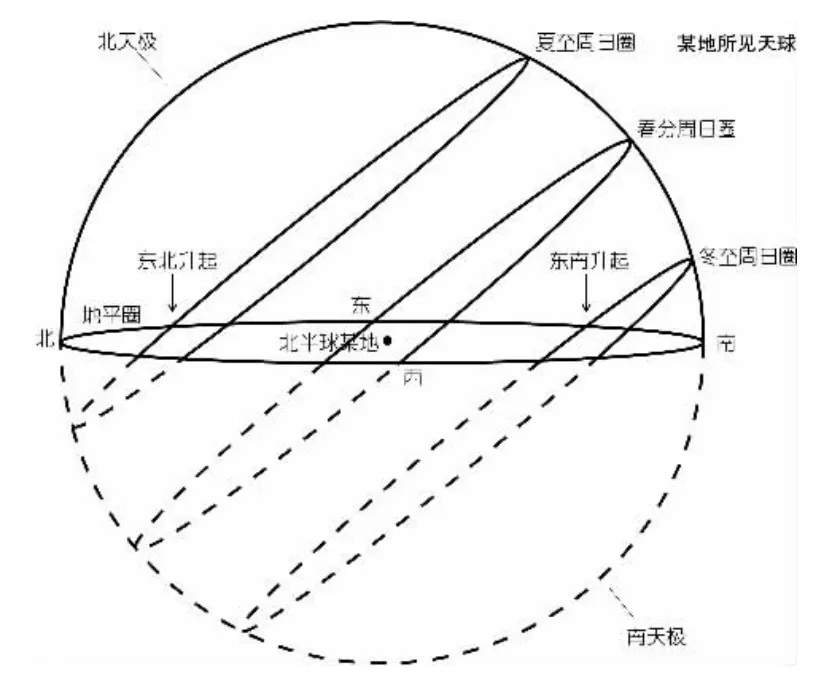
图2 北半球太阳辐射角的年变化
上述的两个理论所描述的情况发生在植物进化早期, 作者猜测在进化早期某种突变体可能产生了这种机制的更加牢固的策略, 试想在进化早期植物多为直立茎, 但某些突变体可能由于上述两个假设而形成了缠绕茎。此时南北半球的缠绕植物的缠绕方向已经基本固定。但是外界条件只能影响植物的形态发育, 却不能影响植物的基因组成。缠绕植物缠绕方向的基因水平固定可能由于在原先不固定缠绕方向的植物(即中性缠绕植物,可因外界条件改变缠绕方向)产生某种机制,或使得生长素横向运输至远心端,或产生其他未知的机制。该种突变体称为右旋/左旋缠绕植物,不能因外界条件改变缠绕方向。由于这种植物有固定的缠绕方向,因此在植物生长过程中出现的异常天气状况不会对其缠绕方向造成误导; 而为进化出这种机制的植物的缠绕方向可能在异常天气时会改变,造成植物“乱长”情况。因此突变株是更有利于生存的,而非突变株被淘汰。
以上的推论是作者根据实际调查情况推测的,据调查和植物祖源分析,左旋的植物多是南半球起源,右旋的植物多为北半球起源,且存在一些中性缠绕植物,可因重力、光照的改变而改变缠绕方向,这一现象称为卡皮罗现象。但目前就化石分析而言, 虽然可以通过化石确定植物的旋转方向, 但由于无法确定植物的缠绕方式产生机理, 因而无法找到确实的证据证明缠绕植物的起源学说。此外,目前尚未在缠绕植物的横截面上发现能够定向横向运输生长素的结构, 此处不同于传统意义上的生长素的横向运输, 传统上指生长素可在横向上因为光照方向的不同而向各个方向运输。
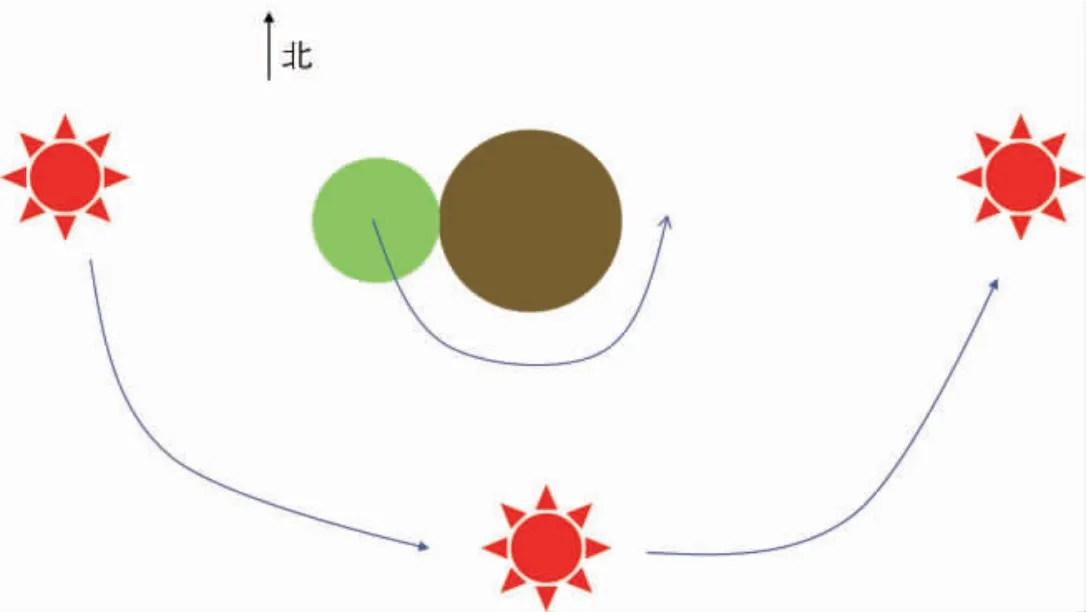
图3 太阳辐射角对植物生长方向的影响
根据假说一的推论, 右旋缠绕植物起源于北回归线以南的地方,左旋缠绕植物起源于南回归线以北的地方, 但是考证植物的起源地很困难, 尤其是诸如本理论需要大量事实证据的假说。因为在上千万年的进化过程中,植物可能经过多次引种,在不同区域进行进化,其分子生物学和形态学上可能已经发生较大的变化,目前只能根据有限的条件进行引证。
但是令人费解的是, 在陈海魁等所做的强制反旋对豇豆生长的影响实验中, 右旋生长的豇豆被强制反旋后反而生长得较好, 而根据假说, 北半球右旋的植物生长状况应当优于左旋的,由于其未实施对于其他生物的实验,本实验可能具有特殊性,在此处仅供质疑。
作者对于验证两个假说的实验概述如下,但由于目前已有充分的研究说明非中性缠绕植物的生长方向与外界条件无关, 因此此处所提到的试验方法仅可用于对中性缠绕植物的研究,可由以下两个实验证明两个假说的正确性,但不能证明其是否在进化早期有作用或证明作者提出的缠绕植物早期的进化理论。
实验一:控制太阳光照的半遮光实验
对于中性缠绕植物, 作者提出一个在特定的某个半球研究光照角度对生长方向的影响的实验。实验需要半遮光板,植物放置在半遮光板中间, 使其一半接受太阳光, 一半不接受太阳光。在北半球, 遮光处在南侧即模仿南半球环境,在北侧即模仿北半球环境。
实验二:科里奥利力实验
将植物放置于不断恒速度旋转的圆盘的非圆心处,同时在植物的正上方放置人造光源,根据圆盘转动方向的不同, 可模仿南北半球的科里奥利力。
对于验证缠绕植物起源性的猜想, 作者认为可通过对缠绕植物进行祖源分析, 推测或估计其大致产生的纬度, 此处的界定标准应该是化石上同源但未产生缠绕特征的植物生活的纬度,若假说成立,推测植物缠绕螺旋的螺距与半径的比值会随太阳在正午时的平均高度角呈现正比关系; 前者可以反映生长速度类似的植物向性生长的速度。
若将校正螺距s定义为因变量, 自变量至少有螺旋半径r、生长速度v、太阳在正午时的平均高度角θ。其中,螺旋半径越大、生长速度越慢,由数学关系得知,在相同的太阳在正午时的平均高度角下植物缠绕的螺距将会更大,且为正比或反比关系。即可能存在:

此处应为正比关系。太阳在正午时的平均高度角θ的定义见图4, 由于夏至的正午太阳高度角和冬至正午太阳高度角的均值为春分或秋分的正午太阳高度角, 因此可认为春分或秋分正午太阳高度角为年正午太阳高度角的均值。
作者认为, 植物生长的校正螺距应与正午年平均太阳高度角呈正比, 因为太阳在某一时刻的位置随纬度变化而变化, 而太阳运行的角速度不变。
而植物的发源纬度α和年平均正午太阳高度角有如下关系:

故因此, 植物缠绕生长的螺距与植物的发源纬度可能存在如下关系:

此式表明,当纬度的绝对值越大时,即发源地越接近两极时,植物的螺距就越小。注意,此处的螺距是经修正过的, 需要排除不同植物生长的速度和植物缠绕半径的问题。
作者认为, 在得出一系列植物发源纬度和植物的校正螺距后,可通过作图,判断是否可以拟合为直线,来验证目前已有的假说一。
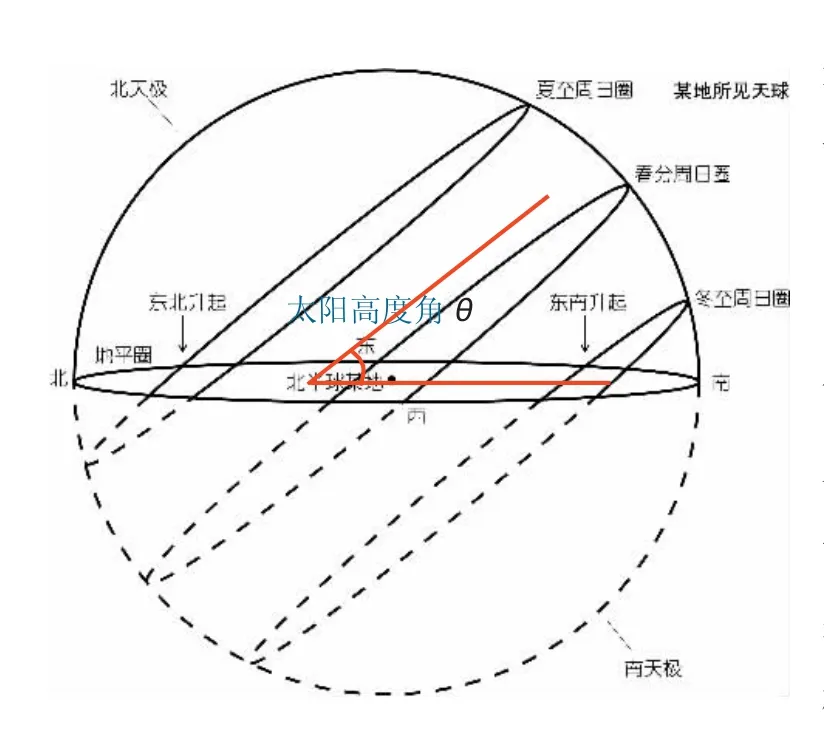
图4 正午平均太阳高度角示意图
综上, 本文对于目前的对于缠绕植物向性缠绕方向的起源假说进行总结, 并提出对于植物进化和植物起源假说的联系。作者设计了两个实验可供实验研究人员研究中性缠绕植物所用。此外,本文采用数学方法对较难量化的植物缠绕指数进行量化, 通过校正螺距排除了植物生长速度和植物缠绕半径的问题。但囿于目前对于缠绕植物缠绕机制的研究较少, 作者没有搜集到足够的数据对文中提出的模型进行拟合和修正。希望研究该方面的研究者可以对于作者提出的模型和进化假说进行修正。
猜你喜欢
军事文摘(2023年24期)2023-12-19 06:50:06
内江科技(2022年3期)2022-03-30 10:24:50
小资CHIC!ELEGANCE(2021年25期)2021-07-29 08:44:26
中国介入影像与治疗学(2020年12期)2020-12-31 06:44:04
军事文摘(2019年18期)2019-09-25 08:09:22
中国医药科学(2019年11期)2019-07-01 10:42:28
山东医药(2019年3期)2019-02-13 19:29:06
机械工程师(2018年7期)2018-07-30 09:05:26
首都食品与医药(2017年3期)2017-04-03 09:12:26
中国卫生标准管理(2015年4期)2016-01-14 05:43:52
