
花青素积累与谷子低温胁迫响应的关系
2020-12-04 06:53蒋茂双王俊杰侯思宇韩渊怀李红英
核农学报 2020年11期
张 彬 李 萌 蒋茂双 王俊杰 侯思宇 韩渊怀 李红英
(山西农业大学农学院,山西 太谷 030801)
谷子[Setariaitalica(L.) P. Beauv.],禾本科狗尾草属,是我国北方重要的杂粮作物和饲草作物,至今已有8 700年的栽培历史[1]。谷子脱去颖壳后称小米,具有较高的营养价值和药用价值。谷子的抗逆性强,不仅较玉米等作物具有更强的抗旱性[2],而且部分品种具有较强的抗寒性,可在黑龙江、内蒙古等高寒地区种植[3]。
花青素属于类黄酮化合物,是植物中重要的次生代谢产物,广泛存在于植物的果实、花和叶片中,使植物呈现紫色、红色和蓝色。花青素不仅可以吸引昆虫帮助植物进行授粉,还参与调节植物对生物及非生物胁迫损伤的抵抗[4],在植物生长过程中具有重要作用。此外,花青素在抗氧化、抗癌以及抗病方面也有一定的医疗和保健作用,如降低胆固醇和血脂,防治心血管疾病,治疗视网膜疾病等,是当今科学的研究热点之一。
低温胁迫限制作物的种植范围,并严重影响作物的品质和产量。低温对植物的损害主要表现为代谢失调、膜通透性改变、呼吸作用紊乱、光合作用降低和有机大分子物质水解等[5-6]。植物花青素的积累受多种环境因素的诱导,低温是主要因素之一。低温处理可显著提高血橙[7]、苹果[8]、甘蓝[9]的花青素含量,进而提高其营养品质。在低温诱导下,茶树紫娟[10]、红叶大头芥[11]、北美短叶松[12]可通过积累花青素进而适应逆境的变化。
部分谷子品种苗期遭遇冷害时其叶片和茎秆会积累花青素而呈现紫红色表型,在拟南芥[13]、番茄[14]等研究中发现低温可以通过诱导花青素的合成来提高植物的抗逆性,但有关谷子花青素的合成代谢及环境因素对其调控机制的研究却鲜有报道。本研究以苗期绿叶谷子品种豫谷1号(YG1)、晋谷21(JG21)和紫叶谷子品种龙爪(B13)、向阳谷(B45)、绳头黄谷(B55)为材料,研究花青素的积累与谷子低温响应的相关性,以期为谷子积累花青素以适应低温环境提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为富含花青素的紫叶谷子品种龙爪(B13)、向阳谷(B45)、绳头黄谷(B55)和绿叶谷子品种豫谷1号(YG1)、晋谷21(JG21)。紫叶谷子品种由2017—2019年在山西农业大学试验田种植的420个谷子品种中筛选得到,绿叶谷子品种由山西农业大学生物工程研究所种质资源库提供。
1.2 试验设计
将试验材料种植于26℃温室,待谷子生长至五叶期,设置2个处理:常温处理为对照组,置于28℃光照16 h/20℃黑暗8 h的人工气候培养箱培养;低温处理为处理组,置于16℃光照16 h/8℃黑暗8 h的人工气候培养箱培养。处理后第8和第12天分别取处理组(记作LTD1,LTD2)和对照组(记作CKD1,CKD2)每个品种的叶片进行生理指标的测定和显微观察。
1.3 测定项目与方法
利用DFC450C光学显微镜(德国徕卡公司)观测叶片中色素的分布;采用盐酸甲醇法测定谷子叶片花青素的相对含量[15];使用LI-6400XT便携光合测定仪(美国LI-COR公司)测定谷子叶片的光合参数:气孔导度(stomatal conductanc,Gs)、胞间CO2浓度(intercellular carbon dioxide concentration,Ci)、净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、水分利用率(water use efficiency,WUE);使用SPAD-502PLUS手持叶绿素仪(石家庄泛胜科技有限公司)测定谷子叶片叶绿素相对含量;采用硫代巴比妥酸法测定丙二醛(malondialdehyde,MDA)含量[16];采用酸性茚三酮法测定脯氨酸(proline,Pro)含量[17];采用蒽酮比色法测定可溶性糖(soluble sugar,SS)含量[18]。
1.4 统计分析
采用Microsoft Excel 2010处理数据并作图,利用SPSS22.0统计软件进行相关性分析和Duncan法多重比较分析。
2 结果与分析
2.1 低温诱导下绿叶和紫叶品种谷子幼苗花青素含量的变化
通过显微镜观测,常温和低温处理下的绿叶品种幼苗均为绿色;而低温处理8 d后,紫叶品种幼苗的叶脉和叶尖开始逐渐积累花青素而呈现紫色;低温处理12 d后,紫叶品种幼苗的整个叶片均转变为紫红色。如图1所示,在显微镜下可清晰观察到低温处理的紫色品种叶片上下表皮由于积累了较多的花青素而呈紫红色;而低温处理的绿色品种叶片的叶脉和上下表皮中,未观察到花青素的存在。
苗期谷子叶片中花青素含量测定结果如图2所示,与常温对照相比,低温条件下YG1花青素含量减少,低温处理8 d的JG21叶片花青素含量显著增加。低温处理下,紫叶品种的花青素含量均显著高于常温对照,说明低温可显著诱导紫叶谷子品种叶片中花青素的积累。

图1 低温处理12 d紫叶品种(A)和绿叶品种YG1(B)的色素分布Fig.1 Pigment distribution of purple leaf variety B13(A) and green leaf variety YG1(B) after 12 days of low temperature treatment

注:不同小写字母表示差异显著(P<0.05)。下同。Note: Different lowercase letters mean significant difference at 0.05 level. The same as following.图2 低温处理对绿叶和紫叶谷子叶片花青素含量的影响Fig.2 Effect of low temperature treatment on anthocyanin contents in green and purple leaves of foxtail millet
2.2 低温诱导下绿叶和紫叶品种谷子幼苗相对叶绿素含量的变化
苗期谷子叶片中相对叶绿素含量如图3所示,绿叶品种YG1在低温处理12 d后,其相对叶绿素含量较常温对照无显著变化。与常温对照相比,低温处理的紫叶品种相对叶绿素含量总体降低,且低温诱导12 d相对的叶绿素含量显著降低。说明低温诱导对绿叶品种谷子幼苗相对叶绿素含量影响较小,但可使紫叶品种谷子幼苗的相对叶绿素含量明显降低。
2.3 低温诱导对绿叶与紫叶品种谷子幼苗光合参数的影响
由表1可知,低温处理下紫叶和绿叶品种谷子幼苗叶片的Gs均低于常温对照。低温处理第8天,除YG1外,绿叶品种JG21和紫叶品种谷子幼苗叶片的Ci均高于常温对照;5个品种低温处理第12天叶片的Ci较常温对照无明显变化规律。低温处理下,绿叶品种JG21和所有紫叶品种叶片的Pn均低于常温对照;YG1的Pn在低温处理8 d时高于常温对照,而在低温处理12 d时低于常温对照。低温处理下紫叶品种和绿叶品种谷子幼苗叶片的Tr均低于常温对照。除B55外,低温处理下绿叶品种和其他紫叶品种谷子幼苗叶片的WUE均高于常温对照。可见,低温诱导在不同程度上影响着谷子品种的光合作用。

图3 低温处理对绿叶、紫叶谷子叶片相对 叶绿素含量的影响Fig.3 Effect of low temperature treatment on relative chlorophyll contents in green and purple leaves of foxtail millet

表1 低温处理对绿叶、紫叶谷子叶片光合参数的影响Table 1 Effect of low temperature treatment on photosynthetic parameters in green and purple leaves of foxtail millet
2.4 低温诱导对绿叶和紫叶品种谷子幼苗相关生理指标的影响
2.4.1 低温对绿叶与紫叶谷子幼苗MDA含量的影响 由图4可知,低温处理的绿叶品种幼苗叶片MDA含量与常温对照相比无明显差异;低温处理12 d时紫叶品种幼苗叶片MDA含量均高于常温对照,且低温处理12 d的MDA含量显著高于低温处理8 d。结果表明,低温诱导较长时间可使紫色品种幼苗叶片MDA含量显著增加。

图4 低温处理对绿叶、紫叶谷子叶片MDA含量的影响Fig.4 Effect of low temperature treatment on MDA content in green and purple leaves of foxtail millet
2.4.2 低温对绿叶和紫叶品种谷子幼苗Pro含量的影响 低温处理12 d后紫叶和绿叶品种叶片Pro含量如图5所示。常温对照的5个品种叶片Pro含量差异较小;低温诱导后,5个品种的Pro含量均显著高于常温对照,且紫叶品种幼苗叶片Pro含量显著高于绿叶品种。说明低温可能通过诱导谷子叶片中Pro积累,从而增强其抵抗低温的能力。

图5 低温处理对绿叶、紫叶谷子叶片Pro含量的影响Fig.5 Effect of low temperature treatment on Pro in green and purple leaves of foxtail millet
2.4.3 低温对绿叶和紫叶品种谷子幼苗中SS含量的影响 由图6可知,低温处理后,5个品种叶片的SS含量均高于常温对照,且绿叶品种SS含量显著低于紫叶品种。结果表明,低温可显著诱导紫叶品种幼苗叶片中SS的积累。
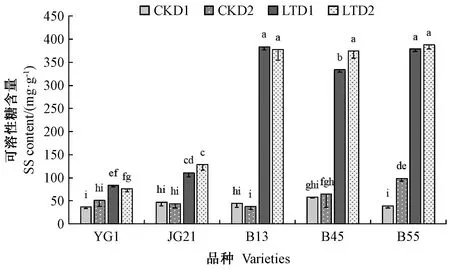
图6 低温处理对绿叶、紫叶谷子叶片SS含量的影响Fig.6 Effect of low temperature treatment on SS contents in green and purple leaves of foxtail millet
2.5 低温诱导下谷子幼苗中花青素含量与相关生理指标的相关性分析
谷子叶片中花青素含量与Pn、相对叶绿素含量、MDA含量、Pro含量以及SS含量之间的相关性分析结果表明,低温处理8 d,花青素含量与Pn呈显著负相关,与SS、Pro含量呈极显著正相关(表2)。低温处理12 d,花青素含量与Pn、相对叶绿素含量呈极显著负相关,与SS、MDA、Pro含量呈极显著正相关(表3)。
3 讨论
低温胁迫会影响谷子的形态特征和生理特性[19],严重时会影响谷子的生长发育和产量[20],但在胁迫过程中植物可以通过调节次生代谢产物的积累来抵御或适应不良环境的影响。本研究显微观察发现谷子叶片花青素主要分布在上下表皮细胞,这与Steyn等[21]对玉米叶片花青素的研究结果相似。且前期已有大量研究证明低温诱导可促进植物花青素的积累[22-24],这也与本研究低温处理下紫叶谷子幼苗花青素含量的测定结果一致。

表2 低温处理8 d花青素含量与生理指标的相关性Table 2 The correlation between anthocyanin content and physiological indexes after 8 days of low temperature treatment

表3 低温处理12 d花青素含量与生理指标的相关性Table 3 The correlation between anthocyanin content and physiological indexes after 12 days of low temperature treatment
研究表明植物组织中叶绿素含量及Pn与花青素含量呈负相关[25]。本研究发现低温处理后紫叶谷子幼苗叶片的Pn和相对叶绿素含量均降低,而花青素含量增加。这与休眠期的小苹果枝条随着气温的降低,其叶绿素含量降低,花青素含量增加的结果一致[26],表明紫叶谷子幼苗可通过调节光合作用、叶绿素及花青素含量的变化来提高其对低温环境的适应性。
MDA是细胞膜脂过氧化的产物之一,能间接体现细胞膜脂受损情况和植物的抗氧化能力。在黄瓜[27]、草莓[28]、番茄[29]等研究中证实通过抑制MDA含量的上升,可提高植株的抗寒性。但本研究中低温胁迫的紫叶品种MDA含量显著高于常温对照,可能是由于低温处理的时间不同而导致生理状况有差异,也可能由于花青素在532 nm波长处也有吸收,所以导致紫叶谷子的MDA含量升高,今后还需通过进一步试验予以验证。
Pro和SS是细胞内重要的渗透调节物质,可通过保护膜结构的完整和调节环境与原生质之间的渗透平衡提高植物抵御逆境的能力[30-31]。在玉米[32]、拟南芥[33]等研究中发现Pro和糖类大量积累可提高植株耐低温能力。这与本研究低温处理后,紫叶谷子幼苗花青素、Pro、SS含量均显著提高的结果一致,且相关分析表明花青素含量与Pro含量及SS含量呈极显著正相关,表明三者共同参与了紫叶谷子幼苗抵御低温的生理代谢过程。而花青素糖基化可以促进花青素的积累,使植物表现出较强的耐逆性[34-35],由此推测,低温胁迫可能主要通过诱导紫叶谷子品种幼苗中SS的积累进而使其花青素含量增加,从而提高谷子低温耐受能力。
4 结论
通过对绿叶和紫叶谷子品种幼苗进行低温处理,发现紫叶谷子通过降低相对叶绿素含量和Pn,增加花青素含量来提高谷子对低温环境的适应性;SS、Pro与花青素共同参与了谷子幼苗对低温胁迫的响应。本研究为了解低温胁迫花青素积累从而提高谷子的抗寒性提供了理论依据。未来可通过选育富含花青素的紫叶谷子品种,扩大谷子在北方较寒冷地区的种植面积,为功能农业的发展奠定基础。
猜你喜欢
现代园艺(2022年9期)2022-06-07
东坡赤壁诗词(2022年2期)2022-04-15
保鲜与加工(2021年1期)2021-02-06
防护林科技(2020年1期)2020-03-26
北方农业学报(2019年6期)2019-02-21
热带作物学报(2018年12期)2018-05-14
中国绿色画报(2017年11期)2018-01-04
现代家庭·生活版(2017年10期)2017-10-20
课堂内外(小学版)(2017年1期)2017-03-08
爆笑show(2015年3期)2015-05-08
