
新疆冬小麦品种农艺及产量性状遗传多样性分析
2020-11-27 13:23:46马艳明冯智宇张胜军倪中福
作物学报 2020年12期
马艳明 冯智宇 王 威 张胜军 郭 营 倪中福 刘 杰,*
研究简报
新疆冬小麦品种农艺及产量性状遗传多样性分析
马艳明1,2冯智宇1王 威2张胜军3郭 营4倪中福1刘 杰1,*
1中国农业大学农学院, 北京 100193;2新疆农业科学院农作物品种资源研究所, 新疆乌鲁木齐 830091;3伊犁州农业科学研究所, 新疆伊宁 835011;4山东农业大学农学院, 山东泰安 271018
产量是小麦育种最重要的目标性状之一, 而农艺性状能直观反映品种的优劣。为了研究新疆冬小麦品种资源农艺、产量性状遗传多样性演变以及对不同生态环境的适应性, 本研究以134份新疆冬小麦地方品种和54份现代育成品种为材料, 分别在新疆乌鲁木齐、新疆伊宁和山东泰安3个不同的生态试验点进行了连续2年的田间种植和农艺性状观察。结果表明, 9个农艺及产量性状的广义遗传力由大到小排序依次为: 株高>粒宽>籽粒长宽比>穗长>小穗数>千粒重>穗粒数>粒长>结实小穗数。进一步对9个农艺性状相关性进行分析, 证明各性状之间并不是相互独立的, 而是彼此相互关联。对新疆冬小麦地方品种与育成品种在不同生态环境下农艺性状的差异进行比较发现, 地方品种的株高、穗长、籽粒长宽比大于育成品种, 而穗粒数、千粒重、粒长、粒宽均小于育成品种, 育成品种农艺及产量性状的变异系数高于地方品种, 体现出近年来新疆小麦育种对株高、穗长、小穗数、穗粒数、籽粒长宽比等农艺性状的重点改良。以上研究结果可为不同地域小麦育种亲本的选择提供重要的参考依据。
新疆; 冬小麦; 地方品种; 育成品种; 农艺及产量性状; 多样性
小麦(L.)是我国第三大粮食作物, 对保障粮食供需平衡和国家粮食安全起着至关重要的作用[1]。农艺与产量性状可以直观地反映品种的优劣, 具有易观察、好测量的特点。20世纪以来, 针对农艺和产量性状的研究在小麦改良中发挥了重要作用, 即使在分子标记技术迅猛发展的今天, 对小麦种质资源和品种农艺性状的考察、分类与科学评价依然是育种工作的一项重要内容[2]。
农作物种质资源是优异基因资源的载体, 也是育种、农业生产以及基础科学研究的重要材料[3]。地方品种亦称农家品种, 是在某一地区长期种植的“土著”品种。由于各地生态条件千差万别, 造成地方品种具有不同的生态适应性, 含有丰富的遗传变异, 是重要的基因资源库。小麦地方品种适应了当地自然环境和生产条件, 是小麦新品种选育的重要基础材料。我国最早育成的一批优良小麦品种就是从农家种中筛选或混选出来的[4]。新疆地域广阔, 气候条件多样, 在长期的生产过程中, 经过人工和自然选择, 产生了大量具有鲜明地域特色, 适应寒冷、干旱、多干热风等生态环境, 且稳产性突出的地方品种。许多新疆地方品种已被用作耐逆、高产小麦育种的亲本材料。系谱分析发现, 新疆本地培育的小麦品种绝大多数都含有地方品种的血缘, 例如, 新冬1号、新冬2号、新冬3号、新冬17等分别由地方品种阿克苏红冬麦、库尔勒白冬麦、巴克甫克、热依木夏等与外国引入品种进行杂交选育而来[4]。
作物品种的生态类型必须与当地生态条件相适应。由于品种与环境之间存在复杂的互作关系, 不同品种在不同生态条件下的适应性和稳产性差异巨大。优良品种通常具有与栽培地区自然、栽培管理条件相适应的一系列优良性状, 能充分利用自然及栽培管理的有利条件, 最大程度发挥其产量潜力[5-6]。通过研究小麦品种资源在不同麦区的产量和品质表现, 可以揭示不同类型品种的生态适应性, 明确品种更替过程中重要性状的演化规律, 为小麦品种选育提供依据。本研究以134份新疆冬小麦地方品种和54份育成品种为材料, 对株高、穗长、小穗数、结实小穗数、穗粒数、千粒重、籽粒大小9个农艺和产量性状进行多年多点调查和系统分析, 比较了地方品种与育成品种主要农艺和产量性状的差异和遗传多样性, 探讨了从地方品种到育成品种演化过程中性状变异的普遍规律, 为进一步挖掘地方品种的育种价值, 指导品种遗传改良提供参考依据。
1 材料与方法
1.1 试验材料
试验选用的188个新疆冬小麦品种, 包括134份地方品种, 54份育成品种, 均为新疆农业科学院农作物品种资源研究所收集保存。分别于2016—2017和2017—2018年小麦生长季, 在新疆农业科学院安宁渠综合试验场、新疆伊犁州农业科学研究所、山东农业大学农学实验站3个试验点进行种植(表1)。试验采取完全随机区组设计, 每小区种植50份材料, 每份材料2行, 行长1.5 m, 每行点播30粒, 3次重复, 收获后参照《小麦种质资源描述规范和数据标准》[7]对株高、穗长、小穗数、结实小穗数、穗粒数、千粒重、粒长、粒宽和籽粒长宽比等9个农艺性状进行考察; 利用万深SC-G型自动种子考种分析仪测定千粒重(thousand kernels weight, TKW)、粒长(grain length, GL)、粒宽(grain width, GW)及籽粒长宽比(GL/GW)。在每个重复小区中随机取10株进行统计分析。
1.2 数据分析
利用IBM SPSS 23对188份供试材料的表型数据进行统计分析, 分别计算各性状的均值、标准差、变异系数和广义遗传力, 并对各性状进行相关性分析、地方品种与育成品种的比较分析。广义遗传力(2)的计算公式如下:
2=G/(G+E) × 100 (G代表遗传方差,E代表环境方差)。
2 结果与分析
2.1 新疆冬小麦品种农艺性状表型分析
对188份小麦品种的株高、穗长、小穗数、结实小穗数、穗粒数、千粒重、粒长、粒宽、籽粒长宽比等9个农艺性状在6种环境下的表型数据进行统计分析发现, 所有考察性状均存在明显的表型变异(表2)。9个性状在各个环境中的变异系数变化范围为5.21%~29.44%, 其中千粒重在第3个环境中的变异最大, 为29.44%, 粒长变异最小, 为5.21%。从各性状在6种环境下的平均值和变异系数来看, 株高平均值为112.94~117.14 cm, 变异系数为12.83%~19.53%; 穗长平均值为11.07~11.95 cm, 随试验点纬度降低略有增加, 变异系数为11.81%~14.01%; 小穗数平均值为19.91~20.47个, 随试验点纬度降低略有增加, 变异系数为7.10%~10.01%; 结实小穗数平均值为19.10~19.89个, 随试验点纬度降低略有增加, 变异系数为7.10%~9.45%; 穗粒数平均值为42.21~52.23个, 表现为伊宁>泰安>乌鲁木齐, 变异系数为13.29%~19.24%; 千粒重平均值为23.76~29.70 g, 随试验点纬度降低千粒重增加, 变异系数为17.13%~27.34%; 粒长平均值为6.43~7.38 mm, 随试验点纬度降低略有降低, 变异系数为5.26%~7.06%; 粒宽平均值为2.63~3.23 mm, 随试验点纬度降低略有降低, 变异系数为8.12%~11.11%; 籽粒长宽比平均值为2.24~2.51, 表现为伊宁>泰安>乌鲁木齐, 变异系数为8.13%~10.01%。各性状均值在不同环境下变化较小, 均呈现连续性分布, 且基本呈现为正态分布。9个性状的广义遗传力从大到小排列依次为: 株高(92.58%) > 粒宽(83.90%) > 籽粒长宽比(80.02%) > 穗长(79.61%) > 小穗数(75.26%) > 千粒重(71.56%) > 穗粒数(68.21%) > 粒长(64.25%) > 结实小穗数(60.44%)。株高广义遗传力最大, 结实小穗数最小, 说明不同品种间株高遗传差异明显, 而品种间结实小穗数遗传差异较小。

表1 试验点情况说明
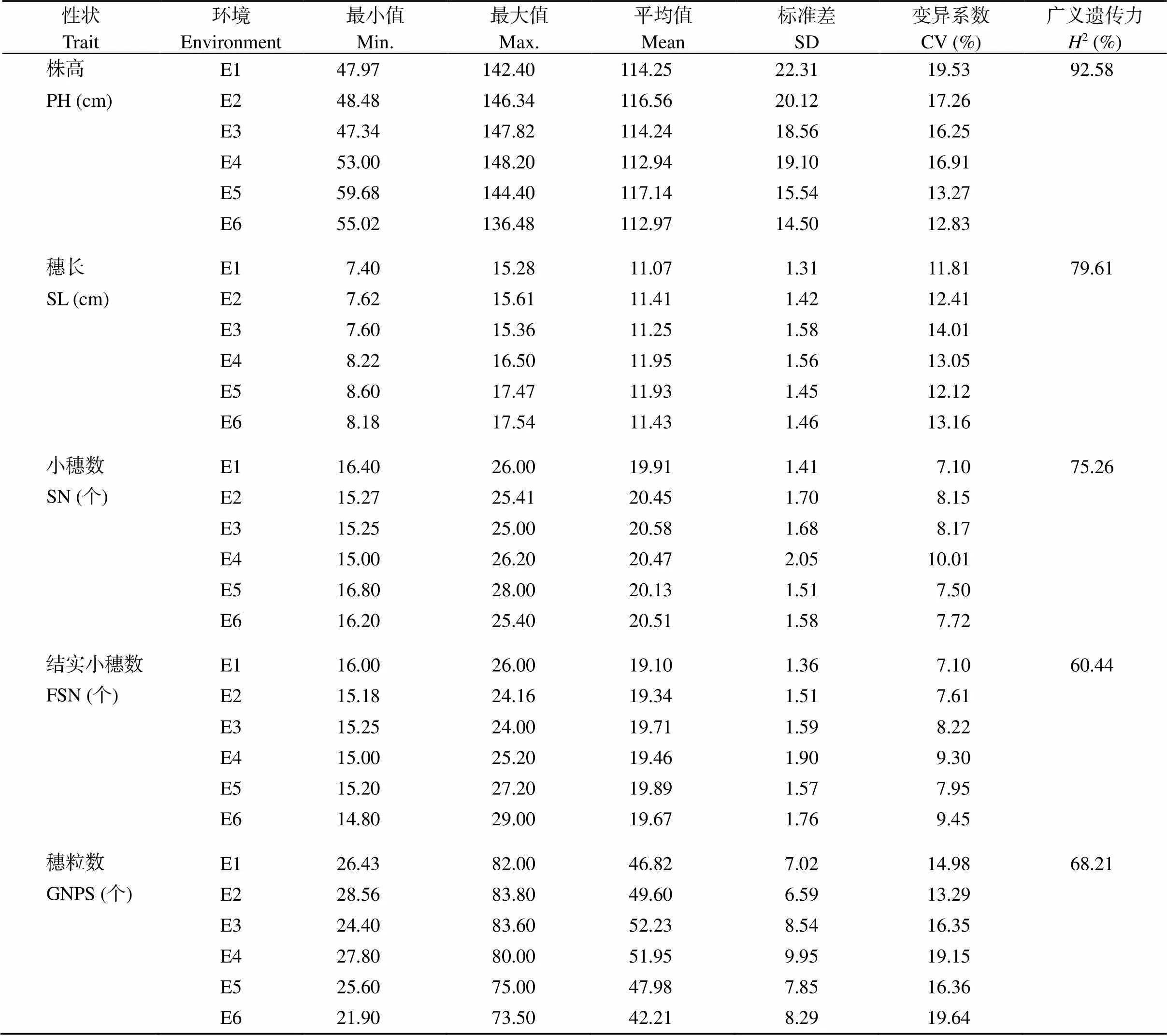
表2 188份小麦品种目标性状描述性统计分析
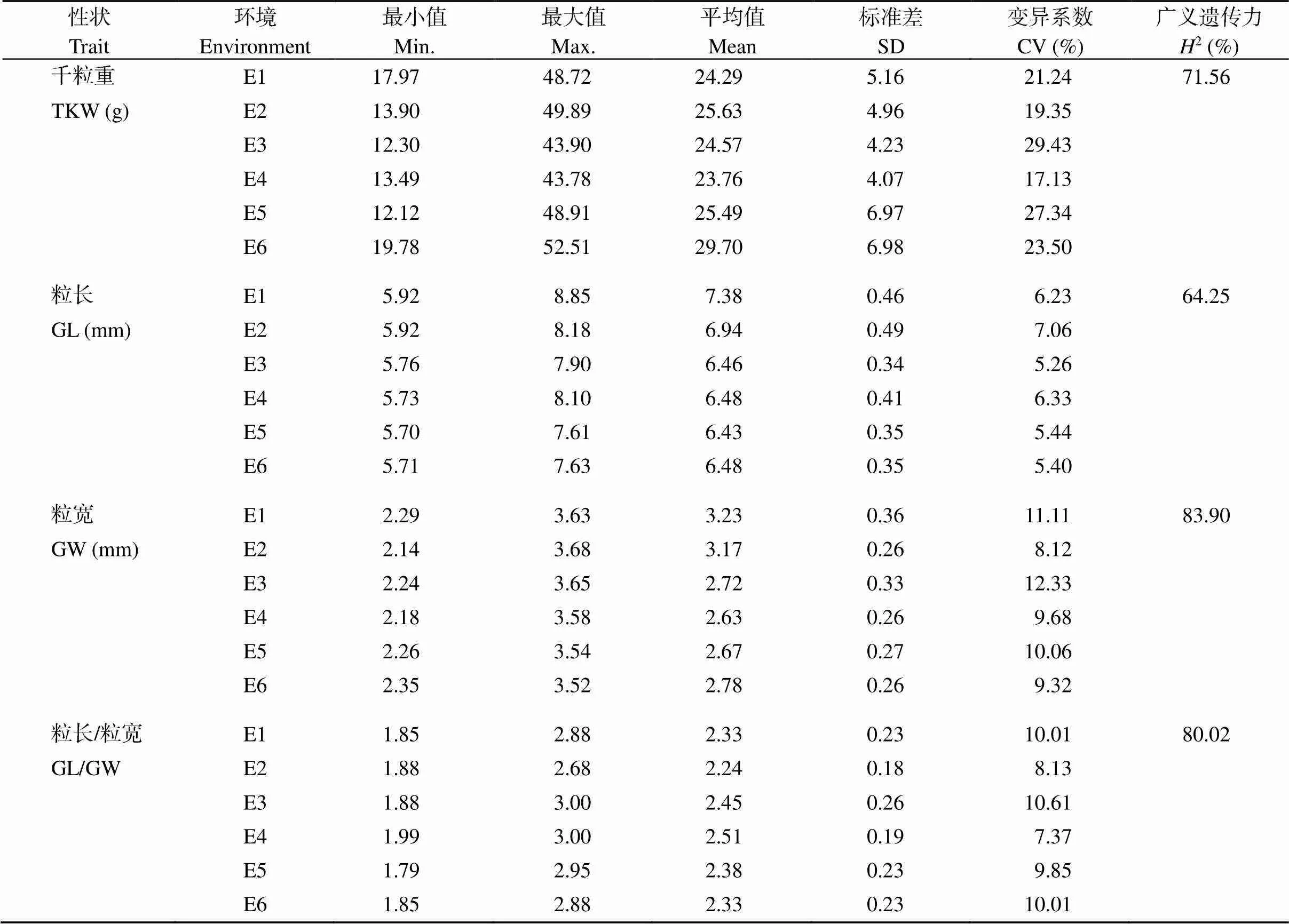
(续表2)
PH: plant height; SL: spike length; SN: spikelet number; FSN: fertile spikelet number per spike; GNPS: grain number per spike; TKW: thousand-kernel weight; GL: grain length; GW: grain width; GL/GW: grain length/width ratio. SD: standard deviation.
2.2 新疆冬小麦品种农艺产量性状相关性分析
性状相关性分析表明(表3), 9个性状之间在6种环境下存在不同程度的相关性。株高在所有环境下与穗长、小穗数均呈极显著正相关(< 0.01), 与千粒重、籽粒长宽比呈极显著负相关; 穗长与小穗数、结实小穗数在6种环境下均呈显著或极显著正相关, 与籽粒长宽比在5种环境下呈极显著负相关、1种环境下呈显著负相关(< 0.05); 小穗数与结实小穗数在除E6环境呈显著正相关外, 在其他5种环境均呈极显著正相关; 小穗粒数与结实小穗数在6种环境下呈极显著正相关, 与小穗数在除E4之外的5种环境呈极显著正相关; 千粒重与粒长在6种环境下呈极显著负相关、与籽粒长宽比呈极显著正相关; 粒长在除E3外的5种环境下与株高呈极显著正相关, 与籽粒长宽比在6种环境下均极显著负相关, 与粒宽在E1、E5和E6这3种环境下呈极显著正相关; 粒宽与籽粒长宽比在除E5、E6外的4种环境下呈极显著正相关。
上述结果表明, 考察的9个农艺性状之间并不是相互独立的, 而是彼此相互关联。在实际生产中, 只有协调好各性状之间的关系, 才能实现小麦产量的提升。
2.3 新疆冬小麦地方品种与育成品种农艺产量性状比较
将新疆冬小麦地方品种与育成品种的9个性状进行简单方差分析发现, 这9个性状的组间平方和均小于组内平方和, 说明地方品种与育成品种之间的差异不是由环境因素产生的, 而是由品种的遗传特性决定的。除结实小穗数之外, 地方品种与育成品种在株高、穗长、小穗数、穗粒数、千粒重、粒长、粒宽、籽粒长宽比这8个性状均呈现极显著的差异, 说明利用新疆冬小麦地方品种与育成品种的表型数据进行比较分析具有统计学意义(表4)。
134份地方品种9个性状的变异系数变化范围在4.63%~19.79%之间, 其中E6环境中的千粒重变异最大, 为19.79%, E3环境中粒长的变异最小, 为4.63%。将9个性状的均值范围/变异系数总结如下: 株高117.30~121.76 cm/5.49%~9.84%; 穗长11.30~12.06 cm/9.20%~12.31%; 小穗数20.01~21.24个/5.20%~7.75%; 结实小穗数19.18~20.46个/5.89%~8.41%; 穗粒数39.24~50.25个/9.48%~16.41%; 千粒重21.68~24.09 g/13.95%~19.79%; 粒长6.37~7.32 mm/ 4.63%~7.36%; 粒宽2.37~2.57 mm/5.94%~7.60%; 籽粒长宽比2.28~2.56/5.56%~8.21%。从上述数据可以看出, 新疆冬小麦地方品种的穗粒数、千粒重等性状受环境影响比较大(表5)。
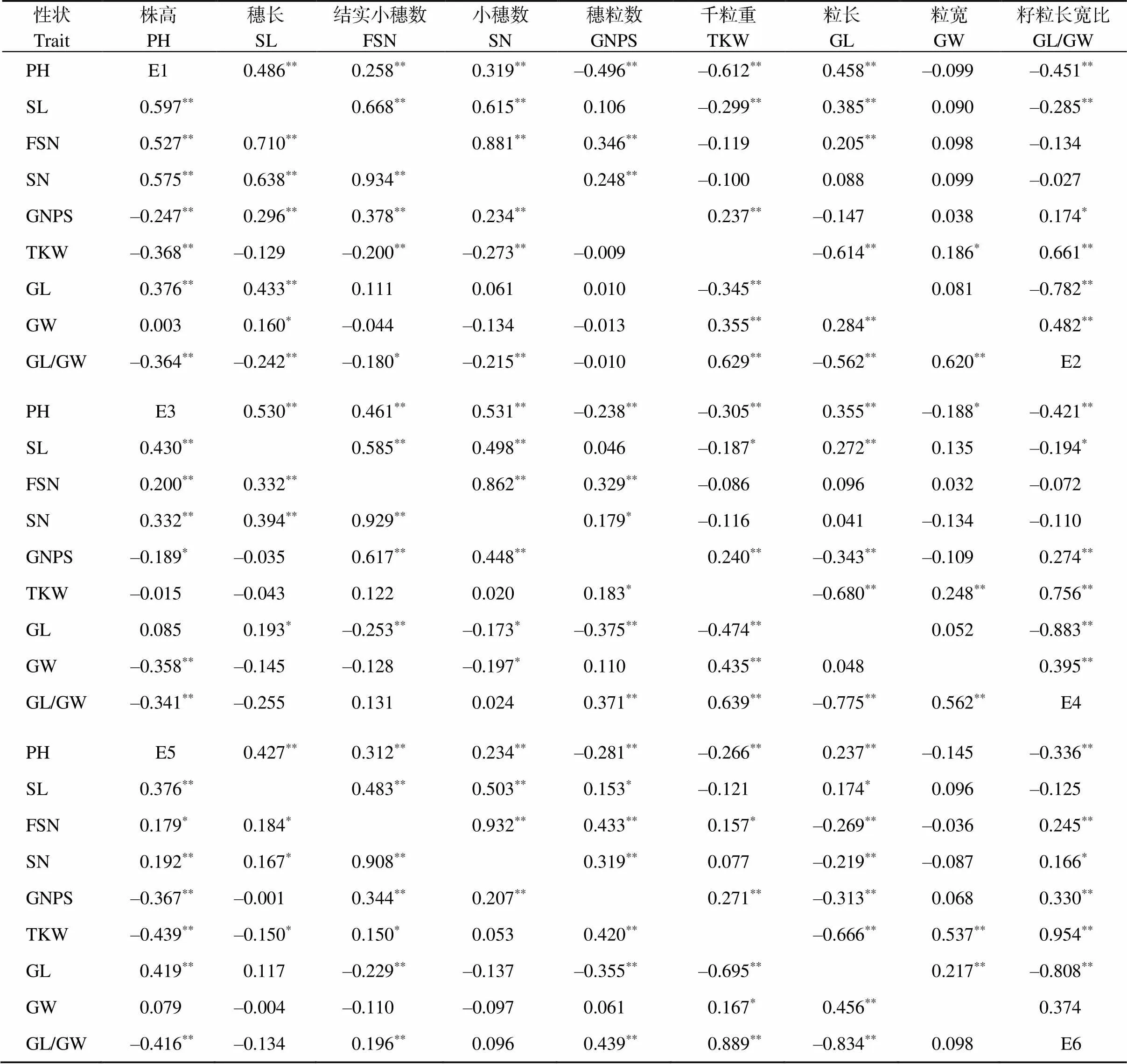
表3 不同环境下各农艺产量性状相关系数分析
*表示差异显著(< 0.05),**表示差异极显著(< 0.01)。
*and**represent the significant differences at< 0.05 and< 0.01, respectively. PH: plant height; SL: spike length; SN: spikelet number; FSN: fertile spikelet number per spike; GNPS: grain number per spike; TKW: thousand-kernel weight; GL: grain length; GW: grain width; GL/GW: grain length/width ratio.
54份现代育成品种9个性状的变异系数范围在5.05%~26.18%, 其中E2环境中的株高变异最大, E6环境中粒长的变异最小。9个性状的均值范围/变异系数分别为: 株高86.82~105.55 cm/19.28~26.18%, 穗长10.27~ 11.15 cm/13.85%~17.43%, 小穗数19.69~21.81个/6.65%~ 12.81%, 结实小穗数18.90~20.58个/8.86%~ 11.58%; 穗粒数平均值为50.15~58.33个/14.88%~20.27%; 千粒重32.02~35.56 g/8.73%~23.67%; 粒长6.45~7.55 mm/5.05%~ 5.69%; 粒宽2.93~3.61 mm/6.81%~9.30%; 籽粒长宽比2.11~2.37/5.65%~9.15%。从变异系数的范围来看, 新疆冬小麦育成品种的小穗数、穗粒数与千粒重3个性状受环境影响比较大(表5)。
将地方品种与育成品种在不同环境下的农艺及产量性状进行比较, 由性状均值差可以看出, 地方品种株高、穗长、籽粒长宽比的变异大于育成品种, 而穗粒数、千粒重、粒长、粒宽的变异均小于育成品种。由变异系数比较发现, 除粒长和粒宽外, 育成品种其余7个性状在6种不同环境下的变异系数均大于地方品种, 说明现代育成品种农艺及产量性状对环境变化表现出更高的敏感性。
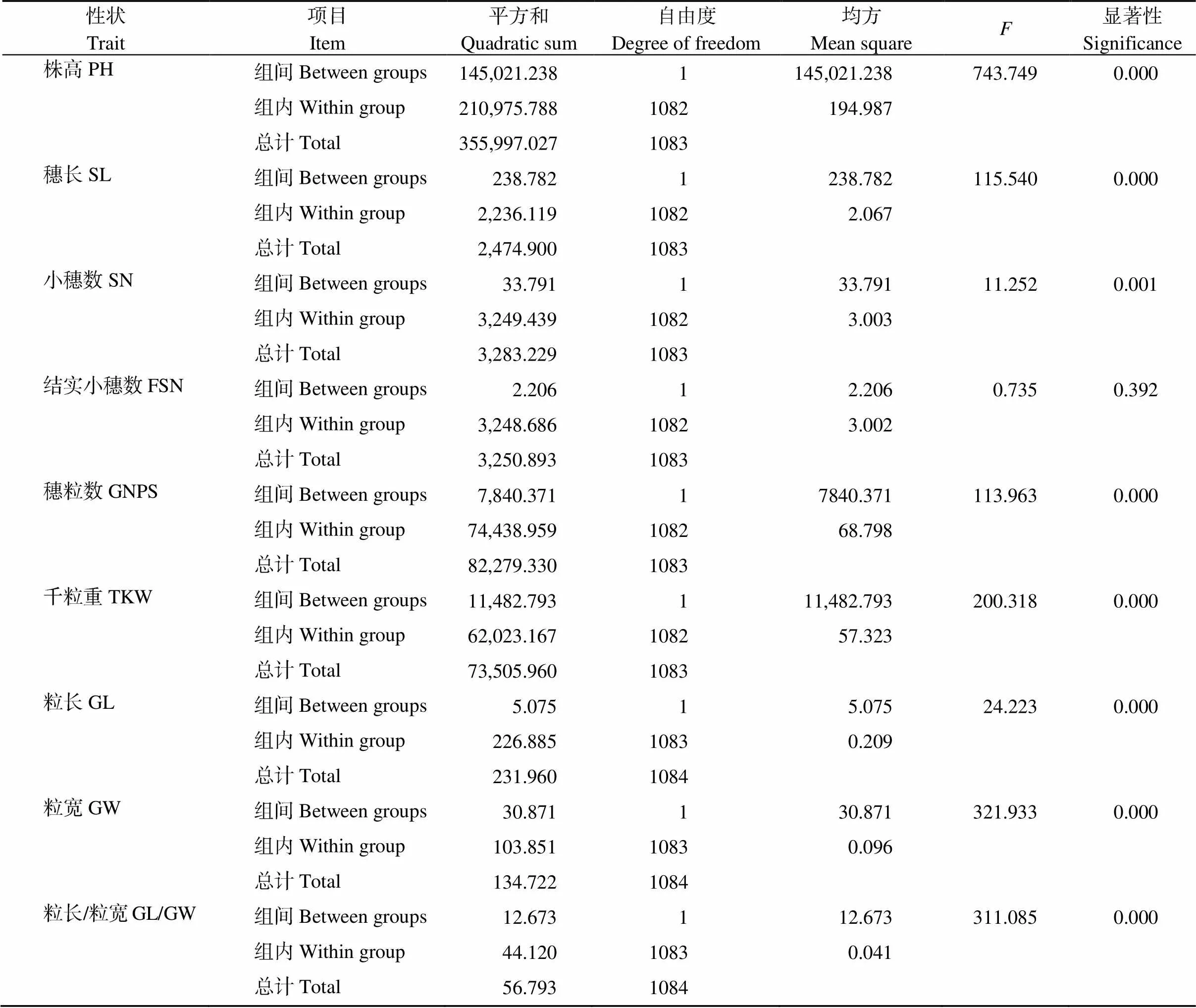
表4 地方品种与现代育成品种间表型性状的简单方差分析
PH: plant height; SL: spike length; SN: spikelet number; FSN: fertile spikelet number per spike; GNPS: grain number per spike; TKW: thousand-kernel weight; GL: grain length; GW: grain width; GL/GW: grain length/width ratio.

(续表5)
PH: plant height; SL: spike length; SN: spikelet number; FSN: fertile spikelet number per spike; GNPS: grain number per spike; TKW: thousand-kernel weight; GL: grain length; GW: grain width; GL/GW: grain length/width ratio.
分析地方品种和育成品种之间农艺性状的变异规律, 有助于深入理解某一地区小麦育种工作的偏好性和发展方向。本研究结果表明, 新疆冬小麦地方品种和育成品种之间在所有考察性状上均存在不同程度的差异, 并且这种差异还受到不同环境的影响(图1), 具体如下: 1)地方品种和育成品种的株高和穗长表型在6种环境下均表现极显著差异, 地方品种的株高、穗长均值均显著大于育成品种; 2)地方品种和育成品种间小穗数差异显著, 其中在E1、E2、E3环境下地方品种小穗数显著高于育成品种, 而E4、E5、E6环境下育成品种小穗数又显著高于地方品种; 3)结实小穗数在地方品种和育成品种间的差异较小, 仅在E2、E4和E6这3种环境下存在差异; 4)育成品种的穗粒数和千粒重均普遍高于地方品种, 且在E3、E5和E6环境下穗粒数差异达到极显著, 而在所有6种环境下千粒重差异均达到极显著水平; 5)地方品种和育成品种的粒长差异较小, 其中在E1、E3和E4环境下存在显著差异, 其他3种环境下无明显差异; 6)与粒长相比, 粒宽在地方品种和育成品种间的差异明显, 在所有环境下, 育成品种的粒宽均显著或极显著大于地方品种; 7)育成品种的籽粒长宽比均低于地方品种, 其中在E1、E3和E5环境下表现为显著差异。

图1 新疆冬小麦地方品种与现代育成品种农艺及产量性状差异
*表示差异显著(< 0.05), **表示差异极显著(< 0.01); ns表示差异不显著。
* and ** represent the significant differences at< 0.05 and< 0.01, respectively; ns: no significant difference.
3 讨论
小麦农艺和产量性状的鉴定与分析对小麦高产育种具有重要的指导意义。性状的变异越大, 表明遗传多样性越丰富。小麦地方品种是重要的种质资源, 新中国成立之初, 生产中95%以上的小麦品种均为地方品种, 是品种选育的重要亲本来源[8]。郝晨阳等[9]研究表明, 我国小麦育成品种的遗传多样性指数和品种间平均遗传距离以50年代最高, 随后逐步下降, 品种间遗传距离到90年代降至历史最低。这种变化趋势与我国小麦育种亲本遗传基础狭窄有关。地方品种通常茎秆细弱、不耐肥水、易倒伏、籽粒小、产量低, 育种家不愿意利用, 现代小麦育种亲本多集中于一些优势骨干亲本, 导致育成品种的遗传基础越来越狭窄。许多研究表明, 小麦地方品种比育成品种具有更强的地域适应性和更广的遗传多样性。刘三才等[10]对中国作物种质资源信息系统的10,000余份小麦资料进行分析发现, 我国小麦选育品种和地方品种在4个穗部性状、5种病害性状、6个农艺和品质性状方面都存在较广泛的遗传多样性差异。除粒色和株高外, 选育品种在穗部性状、农艺和品质性状的变异性呈下降的趋势, 而病害性状的变异性却表现出明显的增加。陈雪燕等[11]对陕西省1225份小麦地方品种的7个形态性状和3个农艺性状进行遗传多样性分析, 发现分别来自陕北、关中和陕南的地方品种各性状的变异大小不同, 认为陕西小麦地方品种存在较广泛的遗传多样性。本研究以134份新疆冬小麦地方品种和54份育成品种为材料, 比较了6种不同环境下主要农艺和产量性状, 发现育成品种株高、穗长、小穗数、可育小穗数、穗粒数、籽粒长宽比的变异系数均大于地方品种, 而粒长、粒宽的变异系数小于地方品种。这一结果体现了新疆地区冬小麦材料的鲜明特点, 即育成品种间产量性状差异较地方品种更为丰富。我们前期利用55K SNP芯片对新疆冬小麦材料进行基因组序列多态性分析也证明, 新疆冬小麦育成品种比地方品种在基因组水平上具有更丰富的SNP序列多态性(作物学报, 已接收), 这为本文所观察到的农艺性状表型变异结果提供了有力的基因组证据。分析原因, 我们认为这可能与新疆冬小麦品种在选育过程中大量引用了国外来源的小麦品种和种质有关, 但深入的原因还有待进一步的研究。
随着生产条件的不断改善和育种水平的不断提高, 我国北部冬麦区小麦品种的组成和主要性状发生了很大变化, 主要表现在株高、穗下节由长变短, 主穗粒数、穗粒重、千粒重由小变大, 这与多年来小麦品种选育以主攻产量的育种目标密切相关[4,8]。从本研究新疆冬小麦地方品种与育成品种各性状的差值可以看出, 地方品种的株高和穗长均大于育成品种, 而穗粒数、千粒重、粒长和粒宽则显著低于育成品种, 这与我国小麦育种对产量性状的普遍偏好性一致。值得注意的是, 与地方品种相比, 现代育成品种的籽粒长宽比显著低于地方品种, 这主要是由于育成品种籽粒粒宽显著增加的同时, 粒长未发生明显变化。这一现象说明, 在地方品种向育成品种的选育过程中, 籽粒形状表现为由原来的长粒、尖粒变成现在的短圆(椭圆)和卵圆粒, 而粒宽的增加是导致育成品种千粒重提高的主要原因。
不同类型小麦品种对不同地区的适应性首先表现在其发育特性与该地区气候条件相适应的程度。任何品种都有其自身固有的生态适应性, 其性状表现是自身遗传因子与环境条件相互作用的结果, 这就造成了不同生态条件下种植的小麦品种表现出不同的生长发育特性[12]。对材料进行多年多点试验可以更准确地反映材料自身的发育特点和遗传特性, 从而实现小麦品种特性的准确评估。目前对于小麦地方品种和育成品种的农艺性状进行遗传多样性研究的报道很多, 但大多是在某一个生态试验点进行比较, 缺乏多个不同生态环境适应性的比较分析[11,13-19]。本研究以134份新疆冬小麦地方品种和54份育成品种为材料, 通过在3个不同生态环境连续2年种植调查的9个农艺性状数据, 分析结果发现同一性状在不同生态环境的变异系数大小有差异, 但地方品种与育成品种的某些性状在不同环境下均表现稳定的差异。例如, 在6种不同环境下, 地方品种的株高、穗长、籽粒长宽比等性状均大于育成品种, 而穗粒数、千粒重和粒宽均小于育成品种, 表明这些性状在很大程度上受品种遗传特性决定, 受环境影响较小。田笑明[20]对新疆冬小麦20世纪50~80年代4个推广时期的32个代表品种的株高、穗粒数等性状进行分析, 其研究结果与本文一致。
随着全球气候变暖, 不同生态麦区的小麦农艺性状和生态适应性, 以及育种和栽培体系、栽培技术都发生了很大的变化[21]。联合国粮食与农业组织(FAO)指出, 未来农作物品种的改良必须适应气候变化, 保持生产的可持续[22]。王亚飞等[23]为了研究我国不同生态麦区小麦品种(系)农艺性状适应气候变化的调控机理, 以黄淮冬麦区北片和南片以及长江中下游冬麦区的20个大面积推广小麦品种、新审定品种和新选育品系为试验材料, 分析了在同一生态环境下主要农艺及产量性状的相关性, 认为由于气候变暖导致南方地区品种(系)在黄淮冬麦区北片有一定的适应, 实际生产中这些麦区的品种可以互相跨区域种植, 性状表现优异的品种可作为亲本材料相互杂交利用。解松峰等[24]在多年多个环境下应用主基因与多基因混合遗传模型方法对杂交组合单世代(P1、P2、RIL7:8、RIL8:9)的单株产量、千粒重、株高等农艺性状进行系统分析, 提出在选育优良品系时, 要综合考虑各重要农艺及产量性状之间, 以及与不同生态环境之间的互作关系。 与之类似, 本研究将新疆冬小麦地方品种和育成品种分别种植在3个不同的生态区域, 结果表明9个农艺及产量性状的广义遗传力由大到小排序依次为: 株高(92.58%)、粒宽(83.90%)、籽粒长宽比(80.02%)、穗长(79.61%)、小穗数(75.26%)、千粒重(71.56%)、穗粒数(68.21%)、粒长(64.25%)、结实小穗数(60.44%), 表明不同生态试验点主要产量性状之间的表现存在差异, 但总体较为稳定。这说明新疆冬小麦品种的生态适应性较广。
[1] 何中虎, 庄巧生, 程顺和, 于振文, 赵振东, 刘旭. 中国小麦产业发展与科技进步. 农学学报, 2018, 8(1): 99–106.He Z H, Zhuang Q S, Cheng S H, Yu Z W, Zhao Z D, Liu X. Wheat production and technology improvement in China., 2018, 8(1): 99–106 (in Chinese with English abstract).
[2] 易腾飞. 中国冬麦区小麦品种农艺性状与品质性状的全基因组关联分析. 河北农业大学硕士学位论文, 河北保定, 2018. Yi T F. Genome-wide Association Study of Agronomic Traits and Quality Traits of Wheat Varieties in the Winter Wheat Region of China.MS Thesis of Hebei Agricultural University, Baoding, Hebei, China, 2018 (in Chinese with English abstract).
[3] 中国农学会遗传资源学会编. 中国作物遗传资源. 北京: 中国农业出版社, 1994. pp 312–315. Society of Crop Genetic Resources Chinese Association of Agricultural Science Societies. Crop Genetic Resources in China. Beijing: China Agriculture Press, 1994. pp 312–315 (in Chinese).
[4] 庄巧生. 中国小麦品种改良及系谱分析. 中国农业出版社, 2003. pp 52, 393–419. Zhuang Q S. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003. pp 52, 393–419 (in Chinese).
[5] 金善宝. 中国小麦生态. 北京: 科学出版社, 1991. pp 9–30. Jin S B. Chinese Wheat Ecology. Beijing: Science Press, 1991. pp 9–30 (in Chinese).
[6] 金善宝. 小麦生态理论与应用. 杭州: 浙江科技出版社, 1992. pp 4–11. Jin S B. Wheat Ecological Theory and Application. Hangzhou: Zhejiang Science and Technology Press, 1992. pp 4–11 (in Chinese).
[7] 李立会, 李秀全. 小麦种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006. pp 81–83. Li L H, Li X Q. Descriptors and Data Standard for Wheat (L.). Beijing: China Agriculture Press, 2006. pp 81–83 (in Chinese).
[8] 董玉琛, 郑殿升. 中国小麦遗传资源. 北京:中国农业出版社, 2000. pp 16–30. Dong Y C, Zheng D S. Chinese Wheat Genetic Resources. Beijing: China Agriculture Press, 2000. pp 16–30 (in Chinese).
[9] 郝晨阳, 王兰芬, 张学勇, 游光霞, 董玉琛, 贾继增, 刘旭, 尚勋武, 刘三才, 曹永生. 我国育成小麦品种的遗传多样性演变. 中国科学C辑, 2005, 35: 408–415. Hao C Y, Wang L F, Zhang X Y, You G X, Dong Y C, Jia J Z, Liu X, Shang X W, Liu S C, Cao Y S. Evolution of genetic diversity of wheat varieties bred in China.(), 2005, 35: 408–415 (in Chinese with English abstract).
[10] 刘三才, 郑殿升, 曹永生, 宋春华, 陈梦英. 中国小麦选育品种与地方品种的遗传多样性. 中国农业科学, 2000, 33(4): 20–24. Liu S C, Zheng D S, Cao Y S, Song C H, Chen M Y. Genetic diversity of landrace and bred varieties of wheat in China., 2000, 33(4): 20–24 (in Chinese with English abstract).
[11] 陈雪燕, 王亚娟, 雒景吾, 吉万全. 陕西省小麦地方品种主要性状的遗传多样性研究. 麦类作物学报, 2007, 27: 456–460. Chen X Y, Wang Y J, Luo J W, Ji W Q. Genetic diversity in main characters of wheat landraces in Shaanxi province., 2007, 27: 456–460 (in Chinese with English abstract).
[12] 李煦征. 江苏省小麦品种更替过程中生态型演化与区域差异分析. 南京农业大学硕士学位论文, 江苏南京, 2007.Li X Z. Research on the Evolution of Ecotypes of Wheat Cultivars and Differences of Regions in Jiangsu. MS Thesis of Nanjing Agricultural University, Nanjing, Jiangsu, China, 2007 (in Chinese with English abstract).
[13] 马艳明, 刘志勇, 肖菁, 热依拉木, 张金波, 苗昊翠, 严勇亮. 新疆冬小麦地方品种主要性状的变异. 麦类作物学报, 2011, 31: 159–165. Ma Y M, Liu Z Y, Xiao J, Reyilamu, Zhang J B, Miao H C, Yan Y L. Analysis on main traits variation of winter wheat landrace varieties in Xinjiang., 2001, 31: 159–165 (in Chinese with English abstract).
[14] 王林海, 王晓伟, 詹克慧, 马东钦, 许兰杰. 黄淮麦区部分小麦种质资源农艺性状的聚类分析. 中国农学通报, 2008, 24(4): 186–191. Wang L H, Wang X W, Zhan K H, Ma D Q, Xu L J. Cluster analysis of some wheat germplasms in Huanghuai area based on agronomic traits., 2008, 24(4): 186–191 (in Chinese with English abstract).
[15] 李志波, 王睿辉, 张茶, 梁虹, 马峙英, 赵玉欣, 王静华. 河北省小麦品种基于农艺性状的遗传多样性分析. 植物遗传资源学报, 2009, 10: 436–442.Li Z B, Wang R H, Zhang C, Liang H, Ma Z Y, Zhao Y X, Wang J H. Genetic diversity analysis of bread wheat (L.) cultivars in Hebei province based on agronomic traits., 2009, 10: 436–442 (in Chinese with English abstract).
[16] 马艳明, 刘志勇, 热依拉木, 肖菁. 新疆冬小麦地方品种与选育品种遗传性状比较分析. 新疆农业科学, 2011, 48: 634–638. Ma Y M, Liu Z Y, Reyilamu, Xiao J. Comparative analysis of genetic characters about landrace and breeding winter wheat varieties in Xinjiang., 2011, 48: 634–638 (in Chinese with English abstract).
[17] 王小国, 梁红艳, 张薇. 新疆春小麦种质资源农艺性状和品质性状的遗传多样性分析. 新疆农业科学, 2012, 49: 796–801. Wang X G, Liang H Y, Zhang W. Analysis of genetic diversity of Xinjiang spring wheat agronomic characters and quality characters., 2012, 49: 796–801 (in Chinese with English abstract).
[18] 程西永. 不同区域小麦种质资源遗传多样性研究. 河南农业大学博士学位论文, 河南郑州, 2010. Cheng X Y. Genetic Diversity of Wheat Germplasm in Different Regions. PhD Dissertation of Henan Agricultural University, Zhengzhou, Henan, China, 2010 (in Chinese with English abstract).
[19] 曾潮武, 梁晓东, 李建疆. 新疆春小麦种质资源主要农艺性状的遗传多样性分析. 分子植物育种, 2017, 15: 3740–3750. Zeng C W, Liang X D, Li J J. Genetic diversity analysis in main characters of spring wheat germplasm in Xinjiang., 2017, 15: 3740–3750 (in Chinese with English abstract).
[20] 田笑明. 新疆冬小麦品种更替中农艺性状演变和发展方向的研究. 作物学报, 1991, 17: 297–303.Tian X M. Study on evolution and its tendency of agronomic characters of winter wheat during cultivar alternation in Xinjiang., 1991, 17: 297–303 (in Chinese with English abstract).
[21] Sun Q, Miao C, Hanel M, Borthwick A G, Duan Q, Ji D, Li H. Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming., 2019, 128: 125–136.
[22] Absentis D. Save and Grow: A Policymaker’s Guide to Sustainable Intensification of Smallholder Crop Production. Italy: FAO, 2011.
[23] 王亚飞, 李世景, 徐萍, 张正斌, 景蕊莲. 黄淮和长江中下游冬麦区小麦品种(系)农艺性状及其聚类分析. 中国生态农业学报, 2020, 28: 395–404. Wang Y F, Li S J, Xu P, Zhang Z B, Jing R L. Agronomic traits and cluster analysis of winter wheat varieties (lines) in the Huanghuai and the middle/lower reaches of the Yangtze River wheat areas., 2020, 28: 395–404 (in Chinese with English abstract).
[24] 解松峰, 吉万全, 张耀元, 张俊杰, 胡卫国, 李俊, 王长有, 张宏, 陈春环. 小麦重要产量性状的主基因+多基因混合遗传分析. 作物学报, 2020, 46: 365–384.Xie S F, Ji W Q, Zhang Y Y, Zhang J J, Hu W G, Li J, Wang C Y, Zhang H, Chen C H. Genetic effects of important yield traits analyzed by mixture model of major gene plus polygene in wheat., 2020, 46: 365–384 (in Chinese with English abstract).
Genetic diversity analysis of winter wheat landraces and modern bred varieties in Xinjiang based on agronomic traits
MA Yan-Ming1,2, FENG Zhi-Yu1, WANG Wei2, ZHANG Sheng-Jun3, GUO Ying4, NI Zhong-Fu1, and LIU Jie1,*
1College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China;2Institute of Crop Germplasm Resource, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, Xinjiang, China;3Institute of Agricultural Sciences of Ili Prefecture, Yining 835011, Xinjiang, China;4College of Agronomy, Shandong Agricultural University, Tai’an 271018, Shandong, China
Grain yield is one of the most important goals in wheat breeding, and agronomic or yield-related traits can directly reflect the characteristics of varieties. In order to determine the evolution of genetic diversity in agronomic traits of Xinjiang winter wheat varieties and their adaptabilities to different ecological environments, 134 winter wheat landraces and 54 moderns bred varieties from Xinjiang were selected for agronomic trait investigation. They were planted in three different ecological environments (Urumqi and Yining in Xinjiang province, and Tai’an in Shandong province) for two consecutive growth seasons, and nine agronomic and yield-related traits were surveyed and analyzed. The estimated broad sense heritability of nine agronomic and yield traits was in descending order: plant height > grain width > grain length/width ratio > spike length > spikelet number > thousand- kernel weight > grain number per spike > grain length > fertile spikelet number. Correlation analyses of nine agronomic and yield traits showed that these traits were correlated with each other. It was found that the plant height, spike length and grain length/width ratio of landraces were higher than that in modern bred varieties, but the grain number per spike, thousand kernel weight, grain length and grain width in landraces were less than that in modern bred varieties. However, the correlation coefficient of these nine traits was higher in modern bred varieties than that in landraces. These variations reflected the evolution of Xinjiang winter wheat varieties in agronomic traits in recent years. This study may provide important information for breeders to select the breeding parents in different winter wheat regions.
Xinjiang; winter wheat; landrace; modern bred varieties; agronomic and yield-related traits; genetic diversity
本研究由国家科技部、财政部国家农作物种质资源共享服务平台(新疆)项目(NICGR2015-029)资助。
This study was supported by the National Ministry of Science and Technology and the Ministry of Finance’s National Crop Germplasm Resources Sharing Service Platform (Xinjiang) Project (NICGR2015-029).
刘杰, E-mail: liujie13251@163.com, Tel: 010-62734072
E-mail: ymma213@sina.com
2020-04-16;
2020-07-02;
2020-08-17.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20200814.1608.002.html
10.3724/SP.J.1006.2020.01034
猜你喜欢
广东农业科学(2021年3期)2021-04-23 04:34:48
国际种业前沿动态(2020年18期)2020-12-23 06:59:17
农业科技与信息(2020年6期)2020-12-18 15:29:14
农业科技与信息(2020年8期)2020-12-18 08:18:14
植物保护(2017年1期)2017-02-13 06:44:34
广东农业科学(2016年11期)2016-03-29 07:52:11
华北农学报(2016年1期)2016-03-18 07:58:57
中学生(2015年4期)2015-08-31 02:53:50
农业科技与装备(2015年5期)2015-05-30 10:48:04
新疆农垦科技(2014年5期)2014-02-28 19:20:00
