
高强度间歇运动对中年小鼠心肌自噬/线粒体自噬相关蛋白表达和线粒体呼吸功能的影响
2020-11-24 08:48:34廖波黄冲胡帅张昱贾潇彭张庆王贵平罗丽
中国运动医学杂志 2020年9期
廖波 黄冲 胡帅 张昱 贾潇彭 张庆 王贵平 罗丽
苏州大学体育学院(江苏苏州215021)
自噬作为细胞的一种分解代谢途径,能将细胞中有害或错误折叠的蛋白质以及受损的细胞器递送至溶酶体中进行降解,从而维持蛋白质稳态和细胞器结构与功能的正常[1]。作为有丝分裂后细胞,心肌细胞的分裂增殖只存在于胚胎期,在胎儿出生以后就几乎不再进行分裂增殖[2],因此,自噬在维持心肌细胞稳态中发挥重要作用。线粒体是细胞的能源中心,通过氧化磷酸化为机体生命活动提供能量。心脏作为人体的“动力引擎”,其正常功能的发挥高度依赖于线粒体呼吸。心肌衰老过程中线粒体数目减少,线粒体呼吸链上的酶活性降低,导致线粒体呼吸功能减弱。更为不利的是,功能异常的线粒体会引起活性氧生成和释放增加,后者又会进一步加剧线粒体损伤,诱导促凋亡因子释放,造成心肌细胞凋亡水平上升。因此,线粒体健康对于维持心肌正常生理功能至关重要。当线粒体发生损伤时,细胞可通过自噬作用选择性清除受损线粒体,这一过程称为线粒体自噬,也是线粒体质量控制的重要机制。线粒体自噬在心肌健康中扮演着重要角色。Parkin 和Bnip3L 是调控线粒体自噬的关键蛋白,有研究发现,在Parkin敲除小鼠的心肌梗死模型中,出现梗死区损伤线粒体数量增多,心肌细胞凋亡水平显著升高[3]。Bnip3L敲除小鼠则在12月龄时表现出明显的心脏肥大[4]。
高强度间歇运动(high-intensity interval training,HIIT)[5]可以提高心脏收缩和舒张能力,增加心输出量,降低心血管疾病患病风险[6],但其发挥心肌保护作用的内在机制仍未阐明。
衰老是心血管疾病发生发展的主要危险因素。衰老过程中自噬功能减退,且自噬衰退的程度与心脏衰老密切相关。研究表明,适宜强度的运动锻炼能通过调控自噬增进健康,延缓衰老[7],但HIIT 对中年心肌自噬/线粒体自噬的调控及其对线粒体呼吸功能的影响仍未阐明。
本研究通过建立中年小鼠HIIT运动模型,探讨HI⁃IT 对心肌自噬/线粒体自噬信号通路和线粒体呼吸功能的影响,为通过科学锻炼促进中年人群心脏健康提供更充分的科学依据。
1 研究对象与方法
1.1 实验动物与分组
12月龄健康雄性ICR 小鼠20 只,购于苏州大学实验动物中心。自由适应环境1 周后,将小鼠随机分为对照组(CON组)、高强度间歇运动组(HIIT组),每组各10 只,单笼饲养,笼内保持通风干燥,自由摄食饮水,CON组小鼠不接受任何形式的运动干预。HIIT组进行为期8 周的运动训练。光照时间为7:00~19:00,温度在23℃± 1℃,相对湿度50%~70%。
1.2 动物训练方案
HIIT 组小鼠首先在小鼠转轮式疲劳仪(图1)(YLS-10B,济南益延科技发展有限公司)中进行为期1周的适应性运动,每次运动15 min,每天1 次,共5 天。休息2天后,对HIIT组小鼠进行运动耐力测试,具体方案是:起始速度为10 转/min,每3 min 增加5 转直至力竭。力竭判定标准:小鼠跟不上设定的运动速度,滑至转轮底部,电击刺激不能促使小鼠继续运动,此时跑步机上的速度记录为小鼠的最大跑步速度(Vpeak)。

图1 小鼠转轮式疲劳仪
HIIT 组运动方案如下:先以14 转/min 的速度进行热身;随后以90%Vpeak的强度运动3min,间歇2 min,间歇期以14 转/min 的速度进行运动,共5 组;接着以12转/min的速度放松3 min。运动时间为下午14:00~16:00。每周运动5天,共8周。
1.3 实验取材
末次运动后24小时,小鼠颈椎脱臼处死后迅速剥离心脏,PBS清洗后滤纸吸干装入EP管,-80℃冰箱保存。
1.4 主要仪器及试剂
YLS-10B 小鼠转轮式疲劳仪,济南益延科技发展有限公司;电泳仪(powerPAC2000 型)、垂直电泳槽、转移电泳槽,美国Bio-Rad公司;Odyssey双色红外激光成像系统,美国LI-COR公司;BioTek酶标仪,美国博腾仪器有限公司;TY-20脱色摇床,上海西巴斯生物技术开发有限公司。
1.5 蛋白免疫印迹(western blot)实验检测心肌组织自噬/线粒体自噬相关蛋白及线粒体呼吸链复合物蛋白表达水平
微管相关蛋白1 轻链3(microtubule-associated protein 1 light chain 3,LC3)的分离胶浓度为12%,自噬相关蛋白5(autophagy-related protein 5,ATG5)、溶酶体关联膜蛋白2 (lysosomal associated membrane protein 2,LAMP2)、线粒体自噬相关蛋白Parkin、BCL2/腺病毒E1B-19kDa 相互作用蛋白3 样(BCL2/adenovi⁃rus E1B 19kDa-interacting protein 3-like,Bnip3L);线粒体标志物细胞色素C 氧化酶Ⅳ(cytochrome C oxi⁃dase subunit Ⅳ,COXⅣ)分离胶浓度为10%,线粒体呼吸链复合物蛋白Ⅰ~Ⅴ(ComplexⅠ~Ⅴ)分离胶浓度为15%。70 V 电泳至目的蛋白条带拉开,300 mA转膜2 h,用1%TBST 配制5%脱脂奶粉,将NC 膜置入牛奶中封闭1 h,一抗孵育过夜。二抗室温摇床孵育2 h,封闭、一抗以及二抗孵育后均用TBST洗膜3次,每次10 min。之后用Odyssey 双色红外激光成像系统扫膜。
抗体厂家、货号及工作浓度如下:LC3(Novus,NB100-2220,1∶500);ATG5(Cell Signaling Technolo⁃gy,12994S;1∶1000);LAMP2(sigma,016M4840V,1∶1000);Parkin(Santa Cruz,SC-32282,1∶1000);Bnip3L(Cell Signaling Technology,#12396S,1∶500);COX Ⅳ(Cell Signaling Technology,#11976,1∶1000);OXPHOS(Abcam,#110413,1∶200);GAPDH(Abcam,#ab9484,1∶10000);tubulin(Abcam,#ab6160,1∶10000)。
1.6 数据处理与统计
所有数据用平均数±标准差(±s)表示。采用Graphpad prism 进行统计分析。采用单因素方差分析进行组间比较,事后多重比较(Post Hoc Tests)采用Newman Keuls 法。P<0.05 为差异显著,P<0.01 为差异极其显著。
2 结果
2.1 8周HIIT对中年小鼠心肌自噬相关蛋白表达的影响
小鼠心肌自噬相关蛋白表达如图2所示,与CON组相比,HIIT组小鼠心肌组织LC3Ⅰ/GAPDH无明显变化,LC3Ⅱ/GAPDH 显著增加(P<0.001),LC3Ⅱ/Ⅰ显著增加(P<0.05)。如图3所示,与CON组相比,HIIT组小鼠心肌组织ATG5蛋白表达显著升高(P<0.05)。如图4所示,与CON 组相比,HIIT 组小鼠心肌组织LAMP2 蛋白表达显著升高(P<0.01)。
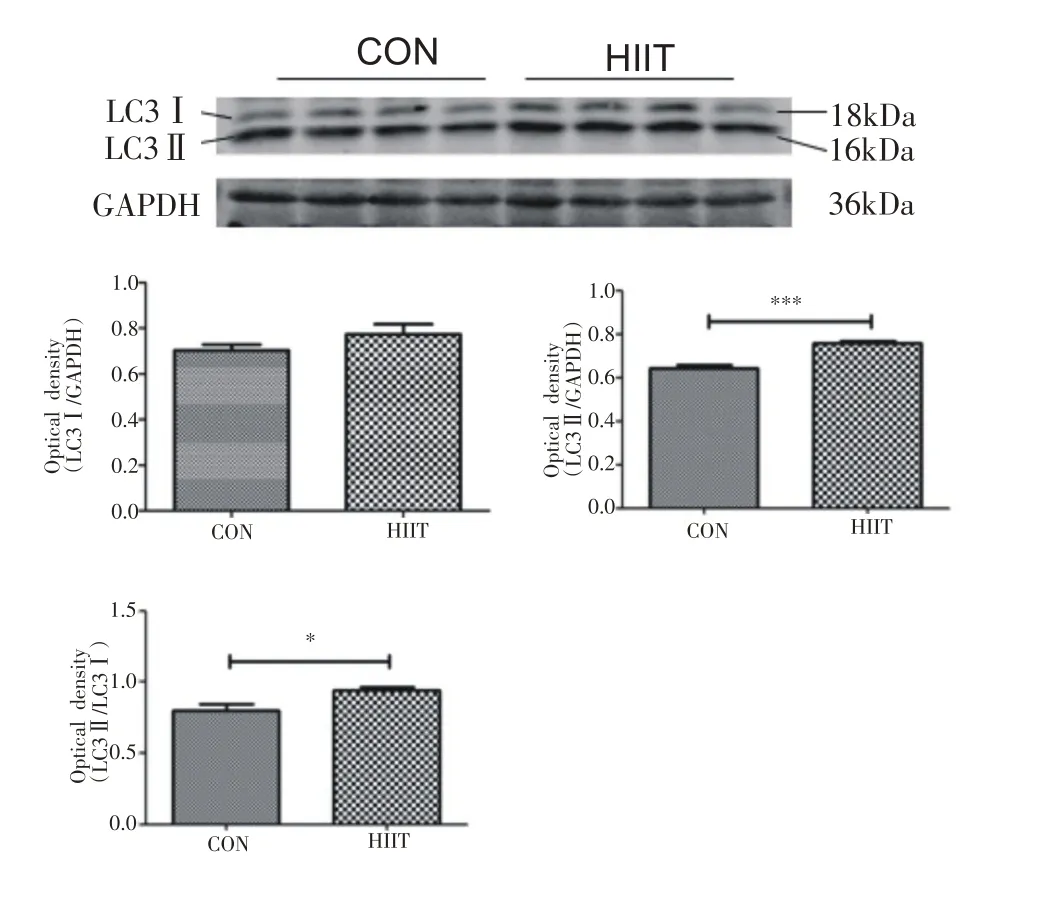
图2 8周HIIT对中年小鼠心肌LC3蛋白表达的影响

图3 8周HIIT对中年小鼠心肌ATG5蛋白表达的影响
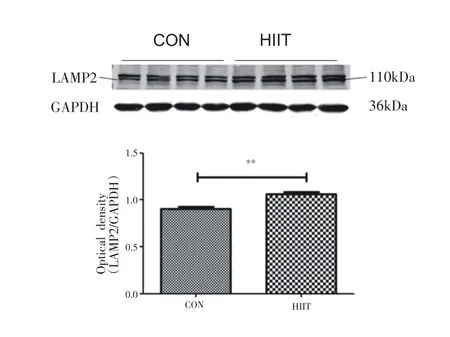
图4 8周HIIT对中年小鼠心肌LAMP2蛋白表达的影响
2.2 8周HIIT对中年小鼠心肌线粒体自噬相关蛋白表达的影响
小鼠心肌组织中线粒体自噬相关蛋白的表达如图5所示,与CON 组相比,HIIT 组小鼠心肌组织Parkin 蛋白表达水平无显著变化。如图6所示,与CON组相比,HIIT 组小鼠心肌组织Bnip3L 蛋白表达水平显著升高(P<0.05)。

图5 8周HIIT对中年小鼠心肌Parkin蛋白表达的影响

图6 8周HIIT对中年小鼠心肌Bnip3L蛋白表达的影响
2.3 8周HIIT对中年小鼠心肌线粒体含量的影响
小鼠心肌组织中COX Ⅳ蛋白表达如图7所示,与CON组相比,HIIT组小鼠COX Ⅳ的蛋白表达水平显著升高(P<0.05)。
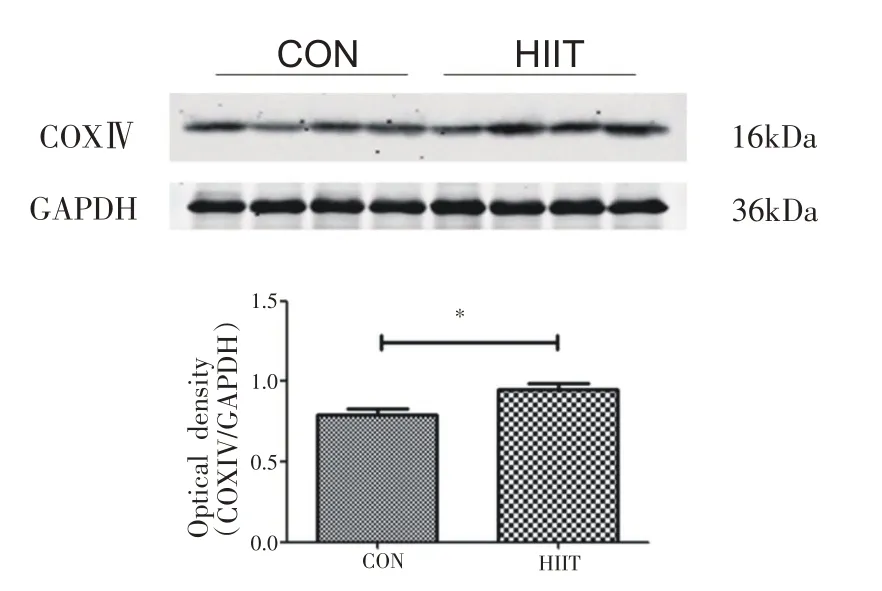
图7 8周HIIT对中年小鼠心肌COX Ⅳ蛋白表达的影响
2.4 8周HIIT对中年小鼠心肌线粒体呼吸链复合物蛋白表达的影响
小鼠心肌组织中线粒体呼吸链复合物蛋白表达如图8所示,与CON 组相比,HIIT 组小鼠心肌ComplexⅠ蛋白表达显著增加(P<0.05),ComplexⅡ~Ⅴ无显著差异。
3 讨论
3.1 8周HIIT对中年小鼠心肌自噬相关蛋白表达的影响
大量研究表明,自噬是一种进化上保守的分解代谢途径,能选择性或非选择性地清除长寿命、非必需或受损的蛋白质和细胞器以维持细胞稳态。当自噬被激活时,自噬相关蛋白ATG12 通过ATG7 作用后与ATG5发生共价结合,形成ATG5-ATG12复合物,该复合物再与ATG16、LC3Ⅱ依次相结合,进而形成ATG5-ATG12-ATG16-LC3Ⅱ终复合物,最终与自噬体膜相结合,促进自噬体膜的弯曲融合,包裹受损的蛋白质和细胞器。随后,自噬体在LAMP2的作用下与溶酶体结合形成自噬溶酶体,降解内容物。LC3 定位于前自噬体和自噬体,分为LC3Ⅰ和LC3Ⅱ,两种形式之间可以互相转化,自噬激活时LC3Ⅰ转化为LC3Ⅱ,因此LC3Ⅱ是自噬体的标志蛋白,可以反映自噬体的数量。LAMP2 是溶酶体特异性标记蛋白,可以反映溶酶体的数量。
自噬是许多生物体衰老的中心机制。在衰老过程中,心脏自噬功能进行性衰退,重要的自噬相关蛋白和自噬调节因子下调。与年轻动物相比,老年小鼠存在Beclin1 和LC3A、B 水平显著下降,这表明伴随着衰老的发生,自噬体的形成过程受到影响[8,9]。许多能够延缓衰老过程的措施,例如热量限制、运动等,其作用的发挥至少部分地与自噬激活有关。但自噬激活改善心肌功能从而延缓衰老的机制仍远未阐明。一些研究探讨了运动的心脏保护作用与自噬功能之间的关系。Ogura 等[10]报道,青年大鼠一次性恒速跑步运动后即刻,心肌LC3Ⅱ水平显著降低,1 h 后显著升高。贾绍辉等[11]发现,长时间中等强度运动可激活小鼠心肌自噬并改善自噬与细胞凋亡之间的平衡,保护线粒体功能。马晓雯等[12]报道中等强度耐力运动可上调大鼠心肌细胞自噬相关蛋白Beclin1、LC3 的表达水平。Cam⁃pos 等[13]报道,8 周有氧运动可以改善心力衰竭小鼠心肌自噬水平,增强心肌功能。与长时间中等强度运动相比,HIIT 可以更好地改善小鼠的运动能力,而这与HIIT诱导的心肌基础自噬适应有关[14]。对于自噬相关基因缺陷(ATG7敲除)小鼠来说,运动并不能改善心肌功能,这进一步证实了自噬在运动发挥心脏保护功能中的重要作用[15]。在本研究中,8周HIIT上调了中年小鼠心肌组织中自噬信号通路中的关键蛋白LC3Ⅱ、ATG5、LAMP2 的表达水平和LC3Ⅱ/Ⅰ比值,提示长期HIIT激活了中年小鼠自噬信号通路。
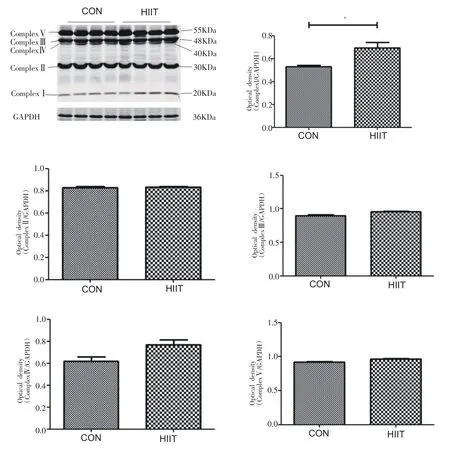
图8 8周HIIT对中年小鼠心肌线粒体呼吸链复合物蛋白表达的影响
3.2 8周HIIT对中年小鼠心肌线粒体自噬相关蛋白表达的影响
心肌细胞有很高的能量需求,主要通过氧化磷酸化产生ATP 以维持正常的收缩功能,因此富含线粒体(占细胞体积的35%)。然而,线粒体功能并不局限于提供能量,还在氧化应激、信号转导、细胞凋亡等中发挥重要作用。线粒体源性的氧化应激可造成生物大分子的不可逆损伤,这在衰老相关的疾病发生发展中具有关键作用。此外,线粒体损伤引起的氧化还原敏感介质的激活,包括核因子-κB等,参与了衰老过程中慢性炎症状态的发生发展[16]。相似的,线粒体损伤还将导致线粒体途径的细胞凋亡水平增加。
线粒体自噬是一种选择性自噬,能清除损伤或衰老的线粒体,以维持细胞内健康线粒体的数量。与神经元和骨骼肌细胞一样,作为有丝分裂后细胞,成年心肌细胞的再生能力非常有限。因此,清除受损的线粒体以确保细胞结构和功能的完整性至关重要。一项利用表达Mito-Keima 小鼠开展的研究表明,心脏是线粒体自噬功能最强健的器官之一,证实了线粒体自噬对于正常心脏功能的重要性[17]。
一些学者探讨了运动对心肌线粒体自噬功能的影响。在线粒体自噬发生的过程中,E3泛素连接酶Parkin能被选择性地募集到线粒体上,促进线粒体自噬。Bnip3L与BCL2/腺病毒E1B相互作用蛋白3(BCL2/ade⁃novirus E1B 19kDa-interacting protein 3,Bnip3)定位于线粒体膜表面,可以在LIR 结构域与LC3直接结合,将LC3 募集到损伤线粒体上促进线粒体自噬。Li 等[18]发现,45 min 急性运动可显著上调青年小鼠心肌Bnip3L 水平,激活线粒体自噬。Zhao 等[19]报道,每天15 min,连续8 周游泳运动可提高青年小鼠心肌线粒体PINK1、Parkin蛋白表达。晚期运动预处理可增强青年大鼠心肌Parkin介导的线粒体自噬,但对Bnip3蛋白表达无影响[20]。这些结果表明,急性和长期运动都可能激活心肌线粒体自噬功能,从而发挥心脏保护作用。
HIIT 在改善心脏功能,降低心血管疾病患病风险方面显示了较大的潜力,但关于HIIT 对中年心肌线粒体自噬的调控作用尚未见报道。本研究中,8 周HIIT对中年小鼠心肌组织Parkin 蛋白表达无影响,但上调了Bnip3L的表达,这提示,8周HIIT可能激活了Bnip3L依赖的线粒体自噬途径。
线粒体自噬选择性清除受损线粒体,是重要的线粒体质量控制机制之一。PINK1/Parkin 通路是目前研究较多的介导线粒体自噬的途径。线粒体损伤时膜电位下降,丝氨酸/苏氨酸激酶PINK1 在线粒体外膜累积。自磷酸化并活化的PINK1招募并磷酸化Parkin,之后Parkin 继续泛素化线粒体外膜蛋白,继而这些底物的泛素化成为自噬降解的信号。Bnip3/NIX 途径是受体介导的线粒体自噬通路。NIX,又称BNIP3L,可通过与自噬体上的LC3 或 γ-氨基丁酸受体相关蛋白(GABARAP)直接结合,启动线粒体自噬。目前研究认为,生理条件下,受体介导的线粒体自噬通路在维持线粒体健康方面发挥更大作用,但在压力或应激条件下,细胞常优先通过PINK1/Parkin 途径清除功能失调的线粒体。迄今为止,人们对不同方式运动调控心肌线粒体自噬的作用和机制仍知之甚少,需要更多的实验对这一领域深入研究。
3.3 8周HIIT对中年小鼠心肌线粒体含量和呼吸链复合物蛋白表达的影响
运动是否能通过促进线粒体生物发生,增强心肌线粒体功能?这是一个令人关注的问题。Qi 等[21]报道,4 周耐力运动提高了8~9月龄小鼠心肌中线粒体DNA 拷贝数,上调了细胞色素C 氧化酶亚单位Ⅱ、Ⅳ,细胞色素氧化酶缺陷同源物1(cytochrome oxidase de⁃ficient homolog 1,SCO1)和细胞色素氧化酶缺陷同源物2(cytochrome oxidase deficient homolog 2,SCO2)转录水平。过氧化物酶体增殖物激活受体-γ共激活因子1(peroxisome proliferator-activated receptor γ coact⁃ive-tor-1,PGC-1α)是调控线粒体生物发生的关键因子,Ko等[22]报道,抗阻训练能上调青年糖尿病小鼠心肌PGC-1α蛋白表达。
线粒体通过呼吸链上的复合物(酶)经由一系列氧化还原反应合成ATP,为细胞提供能量。线粒体呼吸链复合物包括NADH(ComplexⅠ)、琥珀酸氧化还原酶(Complex Ⅱ)、细胞色素C 氧化还原酶(Complex Ⅲ)、细胞色素C 还原酶(Complex Ⅳ)、ATP 合成酶(Com⁃plex Ⅴ)。Rita等[23]对5周龄大鼠进行为期54周的中等强度耐力训练,结果显示运动显著增强了大鼠心功能,增加了心肌线粒体蛋白丰度,增强了线粒体呼吸链复合物Complex Ⅳ和Ⅴ及ATP 合成酶功能。Baekkerud等[24]报道,8周HIIT增强了2型糖尿病小鼠心肌线粒体氧化磷酸化Complex(C)Ⅰ+Ⅱ、CⅡ和CⅣ,但对CⅠ、Ca2+反应无改善。
HIIT 是否能促进中年心肌线粒体生物发生,改善线粒体呼吸功能?本研究为这一问题的解决提供了更多证据。COXⅣ位于线粒体内膜,可反映线粒体的数量,同时也是线粒体呼吸链的终端酶,能与氧气直接作用,与线粒体氧化能力以及线粒体生物发生密切相关。本研究中,HIIT组小鼠COX Ⅳ的蛋白表达水平显著升高,提示8 周HIIT 增加了中年小鼠心肌线粒体数量。此外,8 周HIIT 上调了中年小鼠心肌线粒体Com⁃plexⅠ蛋白表达,但对线粒体呼吸链中的其他酶蛋白表达水平无显著影响。ComplexⅠ是线粒体呼吸链的关键蛋白,其活性的降低是心力衰竭的主要特征之一。这一现象是否是HIIT诱导中年小鼠心肌线粒体适应的独特效应?HIIT调控心肌线粒体呼吸链复合物表达的关键因素和具体机制如何?这些问题仍有待于进一步深入研究。
4 总结
本研究探讨了8周HIIT对中年小鼠心肌自噬/线粒体自噬相关蛋白表达和线粒体呼吸功能的影响,结果显示,8 周HIIT 引起心肌组织自噬相关蛋白LC3Ⅱ、LC3Ⅱ/LC3Ⅰ、ATG5、LAMP2 表达水平显著升高;线粒体自噬相关蛋白Bnip3L 显著增加;线粒体标志物COX Ⅳ显著增加;线粒体呼吸链复合物Complex Ⅰ表达显著升高。这些结果表明,8 周HIIT 激活了中年小鼠心肌自噬/线粒体自噬信号通路,增加了线粒体数量,改善线粒体呼吸功能。
猜你喜欢
海洋通报(2021年1期)2021-07-23 01:55:14
原子与分子物理学报(2021年2期)2021-03-29 07:30:46
生物学通报(2021年4期)2021-03-16 05:41:26
中国临床医学影像杂志(2019年1期)2019-04-25 06:49:44
中成药(2018年7期)2018-08-04 06:04:18
中成药(2018年3期)2018-05-07 13:34:18
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:34
山东医药(2015年14期)2016-01-12 00:39:45
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
长江大学学报(自科版)(2014年27期)2014-02-27 07:08:27
