
库尔勒香梨黑斑病病原鉴定
2020-11-20 04:25任美佳张华平钟聪慧张世卿黄家风
石河子大学学报(自然科学版) 2020年5期
任美佳,张华平,钟聪慧,张世卿,黄家风*
(1 石河子大学农学院/新疆绿洲农业病虫害治理与植保资源利用重点实验室,新疆 石河子 832003;2 新疆生产建设兵团第一师农业技术推广站,新疆 阿拉尔 843300)
库尔勒香梨栽培历史悠久、香甜可口、脆爽多汁,具有很高的商品价值与很大的市场竞争优势,在新疆水果产业中占据重要地位。但近几年黑斑病的发生使库尔勒香梨生产遭受严重危害,尤其是2018年梨黑斑病导致阿拉尔市库尔勒香梨大面积掉果、烂果,导致近20 hm2果园几乎绝收,对果农造成了严重的经济损失。
研究表明,梨黑斑病主要由链格孢属(Alternaria)真菌引起,目前,全世界从梨上分离并命名的链格孢有9个种[1-3],中国在梨上发现的链格孢有6个种,分别为交链格孢(A.alternata)、细极链格孢(A.tenuissima)、梨黑斑链格孢(A.gaisen)、侵染链格孢(A.infectoria)[3-4]、鸭梨侵染链格孢(A.yaliinficiens)和A.ventricosa[1-2]。链格孢属真菌种类繁多,兼具寄生性与腐生性,在作物生长期或贮藏期均能侵染植物造成危害,如A.mali在苹果上[5]、A.brassicae在生菜和胡萝卜上[6-7]、A.viniferae在葡萄上[8]、A.cucumerina在南瓜、甜瓜、西葫芦等瓜类作物上[9]引起黑斑病或叶枯病,导致品质下降、产量降低,因此链格孢属真菌是引起植物病害的重要类群之一。
链格孢属真菌分类是一个很有争议的问题,形态特征一直是链格孢属分类的主要依据,如梨黑斑链格孢(A.gaisen)相对于交链格孢(A.alternata),孢子更大,孢子链更短、几乎没有分支,因此可以依据孢子大小与产孢方式将其进行有效区分[10]。然而对于大多数小孢子种,因物种之间的高度相似性以及存在一些具有中间性状的菌株,仅凭形态学特征很难进行种类鉴定[11]。因此,越来越多的种类鉴定增加了基因进化关系作为分类依据,如基于β-微管蛋白基因(tub)、过敏原基因(Alt-a1)、组蛋白基因(His)、翻译延长因子基因(tef1)、甘油醛-3-磷酸脱氢酶基因(gpd)、内切半乳糖醛酸酶基因(endoPG)构建单基因系统发育树,通过系统进化关系对链格孢进行种类鉴定,有力促进了链格孢分类[11-15]。但是该方法依然存在局限,因为分别以不同的单基因构建系统发育树对同一个菌株进行种类鉴定时,得到的结果往往不一致[16]。近年发展的多基因联合系统发育分析法,在一定程度上克服了单基因存在的横向基因转移和不同基因间进化速率差异对系统发育树的影响,得到的结果比单基因系统发育分析更为准确,在原核和真核生物的系统发育研究中得到了广泛应用[17]。如Somma等基于翻译延长因子基因(tef)、β-微管蛋白基因(tub)、甘油醛-3-磷酸脱氢酶基因(gpd)和过敏原基因(Alt-a1)构建多基因联合系统发育树,对引起小麦黑斑病的164个链格孢菌菌株进行了鉴定,明确了菌株的种类及其进化关系[16]。
本研究为了明确新疆阿拉尔市库尔勒香梨发生的黑斑病病原,对病株中分离到的链格孢菌进行了致病性测定,在形态学基础上结合多基因联合系统发育分析,基于过敏原alt-a1基因(Alt-a1)、钙调蛋白基因(Cal)和翻译延长因子基因(tef1)构建多基因联合系统发育树,对病原菌进行了种类鉴定,为科学防治梨黑斑病提供依据。
1 材料与方法
1.1 试验材料
2018年9月于新疆生产建设兵团第一师八团梨园随机采集表现明显黑斑的库尔勒香梨病果41个,4 ℃冰箱保存用于病原菌分离。致病性鉴定所需梨果为同一地区采集和市售的健康库尔勒香梨。
1.2 梨黑斑病病原菌的分离与纯化
对具有典型黑斑症状的果实,采用常规组织分离法在病健交界处切取0.5 cm×0.5 cm组织块,依次用0.1%的升汞消毒20 s,75%乙醇消毒10 s,灭菌水清洗3次。然后将组织块用灭菌滤纸吸干置于PDA培养基上,25 ℃培养5 d;对分离得到的真菌再通过单孢分离获得纯培养菌株。将分离纯化的菌株在PDA培养基上培养5 d,根据菌落形态和分生孢子的形态将初步确定为链格孢的菌株用20%甘油保存于-80 ℃备用。
1.3 分离菌株的致病力测定
将分离纯化的链格孢菌株在PDA培养基上培养5 d,用灭菌打孔器打取直径为1 cm菌饼;再用75%乙醇对库尔勒香梨健康果实进行表面消毒;然后分别通过无伤和有伤接种对分离纯化的菌株进行致病性鉴定。无伤接种是将菌块直接贴于健康梨果表面,用封口膜固定,于25 ℃保湿培养[18]。有伤接种是先用灭菌针在健康果面刺约3 mm深的小孔,再将菌块贴于健康梨果表面,用封口膜固定,于25 ℃保湿培养。每24 h观察接种发病情况,待发病揭去菌饼。每个菌株设3个重复,以未接菌的PDA贴块作为空白对照。
1.4 梨黑斑病病原菌的形态学鉴定
将有致病力的链格孢菌株根据其在PDA上的培养特征各选取1个代表菌株(P8和P21菌株)进行形态学鉴定。将菌株在PDA培养基上培养3 d,用灭菌刀片在菌落边缘切取大小为3 mm×3 mm×2 mm的菌块,置于灭菌载玻片中央,用盖玻片压住菌块边缘,在25 ℃明暗交替条件下继续培养7 d,在光学显微镜下观察盖玻片下病原菌的产孢方式;对PDA培养基上培养7 d的菌株,用灭菌牙签挑取平板上的孢子,在光学显微镜下观察病原菌分生孢子的形态、颜色及隔膜数,至少测量100个分生孢子大小并进行统计。根据相关文献[19]进行病原菌形态学鉴定。
1.5 梨黑斑病病原菌的分子鉴定
将代表菌株P8和P21分别在PDA培养基上培养5 d,用灭菌载玻片轻轻刮取培养基表面的真菌菌丝和分生孢子,利用真菌DNA提取试剂盒(BioFlux)提取代表菌株的DNA。分别用过敏原基因(Alt-a1)的引物AltaF(5′-ATGCAGTTCACCACCATCGC-3′)和AltaR(5′-ACGAGGGTGAYGTAGGCGTC-3′)[13]、钙调蛋白基因(Cal)的引物CalF(5′-AGCAAGTCATCTCCGAGTTCAAGG-3′)和CalR(5′-CTTCTGCATCAYCTGGACG-3′)[20]、翻译延长因子基因(tef1)的引物EF-1(5′-CATCGAGAAGTTCGAGAAGG-3′)和EF-1R(5′-TACTTGAAGGAACCCTTACC-3′)[21]从各菌株基因组DNA中扩增相应的基因片段并测序。将测序结果在NCBI(National Center for Biotechnology Information)上通过BLAST比对;在GenBank中下载相关参考序列,分别按照Alt-a1和tef1的顺序、Alt-a1和Cal的顺序进行基因序列整合;以匍柄霉(StemphyliumVesicarium)相应的基因序列为外缘[16],在MEGA 7.0.20软件中按照BIC标准选择最佳模型,通过最大似然法Maximum Likelihood(ML)构建多基因联合系统发育树,对链格孢代表菌株进行进化分析。
2 结果与分析
2.1 病原菌分离与致病性测定
对库尔勒香梨上表现明显黑斑的病组织进行真菌分离,共获得36个纯培养菌株,根据菌落特征和分生孢子形态,其中34个菌株可初步确定为链格孢属(Alternaria)真菌,占分离菌株总数的94.4%。如图1所示,以P8为代表的菌株在PDA上形成的菌落外圈具白色边缘、中间棕绿色、最内圈深褐色,有角状花纹,占链格孢菌株的23%;以P21为代表的菌株菌落最外圈白色边缘、中间棕绿色、深褐色内圈与P8菌株相比明显变小,无明显角状花纹,占链格孢菌株的61.5%。对34个链格孢菌株进行无伤和有伤接种,结果均能在健康果面造成与田间相似的症状(图1):田间自然发病的病斑初期为黑褐色圆形小点,随梨果生长病斑逐渐变大,且稍有凹陷,部分病斑为同心轮纹斑。对健康梨果进行有伤接种,3 d即可在接种部位观察到黑褐色坏死斑点,随着接种时间的延长,病斑逐渐变大并略有凹陷;无伤接种5~7 d时才能在接种部位产生黑褐色坏死斑点,病斑形状与有伤接种相似。上述结果表明,从库尔勒香梨上分离获得的链格孢是导致黑斑病的致病菌。

图1 库尔勒香梨黑斑病的症状及病原菌培养性状
2.2 病原菌的形态学鉴定
从上述致病的链格孢中选取P8和P21菌株作为代表菌株进行形态学鉴定。如图2所示,P8菌株的分生孢子梗为褐色,具分枝。分生孢子短链生,孢子链一般由10个以内孢子构成;孢子链的中间孢子与顶端孢子基部以合轴式延伸产孢,形成树状短分支的分生孢子链。分生孢子为倒棍棒形、卵形或长椭圆形;淡褐色至褐色,表面光滑或具微刺;具2~5个横膈膜,0~3个纵隔膜;大小为(6.0~12.5) μm×(11.2~34.5) μm,平均大小为9.3 μm×20.3 μm;无喙或具短喙,部分喙可转变为产孢假喙,喙及假喙长0~17.7 μm。根据这些形态学特征,初步将P8菌株的种类鉴定为交链孢(Alternariaalternata)。

A至C为产孢梗及产孢方式;D至F为分生孢子。
如图3所示,P21菌株的分生孢子梗淡褐色,偶分枝。分生孢子链生,一般由10个以上孢子构成;因多数孢子只在假喙顶端次生产孢,所以分生孢子长链多不分枝。分生孢子倒棍棒形或倒梨形,淡褐色至中褐色,表面光滑或具微刺;成熟分生孢子具4~7个横隔膜,0 ~2个纵隔膜,孢子中部有2~3个主横隔膜,明显较粗,略隘缩;大小为(8.6~16.2)μm×(15.9~41.2)μm,平均值为 12.7 μm×25.9 μm,喙及假喙柱状,长0~44.1 μm。根据这些形态学特征,初步将P21菌株的种类鉴定为细极链格孢(Alternariatenuissima)。

A至B为分生孢子链;C为产孢梗;D至F为分生孢子。
2.3 病原菌的分子生物学鉴定
对P8菌株和P21菌株的基因组DNA进行过敏原基因(Alt-a1)、翻译延长因子基因(tef1)、钙调蛋白基因(Cal)的PCR扩增,P8菌株获得3个基因长度分别为472、240、787 bp,P21菌株获得3个基因长度分别为472、240、786 bp。将所有测序结果在NCBI进行比对,结果显示,2个菌株的Alt-a1基因均与交链孢(A.alternata)和A.citri具有最高序列相似性(100%);P8菌株的tef1基因与交链孢具有最高序列相似性(98.3%),P21菌株的tef1基因与交链孢、细极链格孢(A.tenuissima)和A.citri均具有100%序列相似性;P8和P21菌株的Cal基因均与A.malvae具有最高的序列相似性,分别为100%和99.6%。因此从单个基因的测序结果无法对2个菌株的种类进行确定。
基于Alt-a1和tef1基因序列构建多基因联合系统发育树(图4),P8菌株单独与交链孢的2个菌株(LH-4菌株和HMCH-9菌株)聚类在一个小的进化分支上;基于Alt-a1和Cal基因序列构建基因联合系统发育树(图5),P21菌株与细极链格孢的3个菌株(EGS38.029、EGS34.015和X1147菌株)聚类在一个进化分支上。因此系统发育分析进一步支持了上述形态学鉴定结果,明确P8菌株的种类是交链孢(A.alternata),P21菌株的种类是细极链格孢(A.tenuissima)。
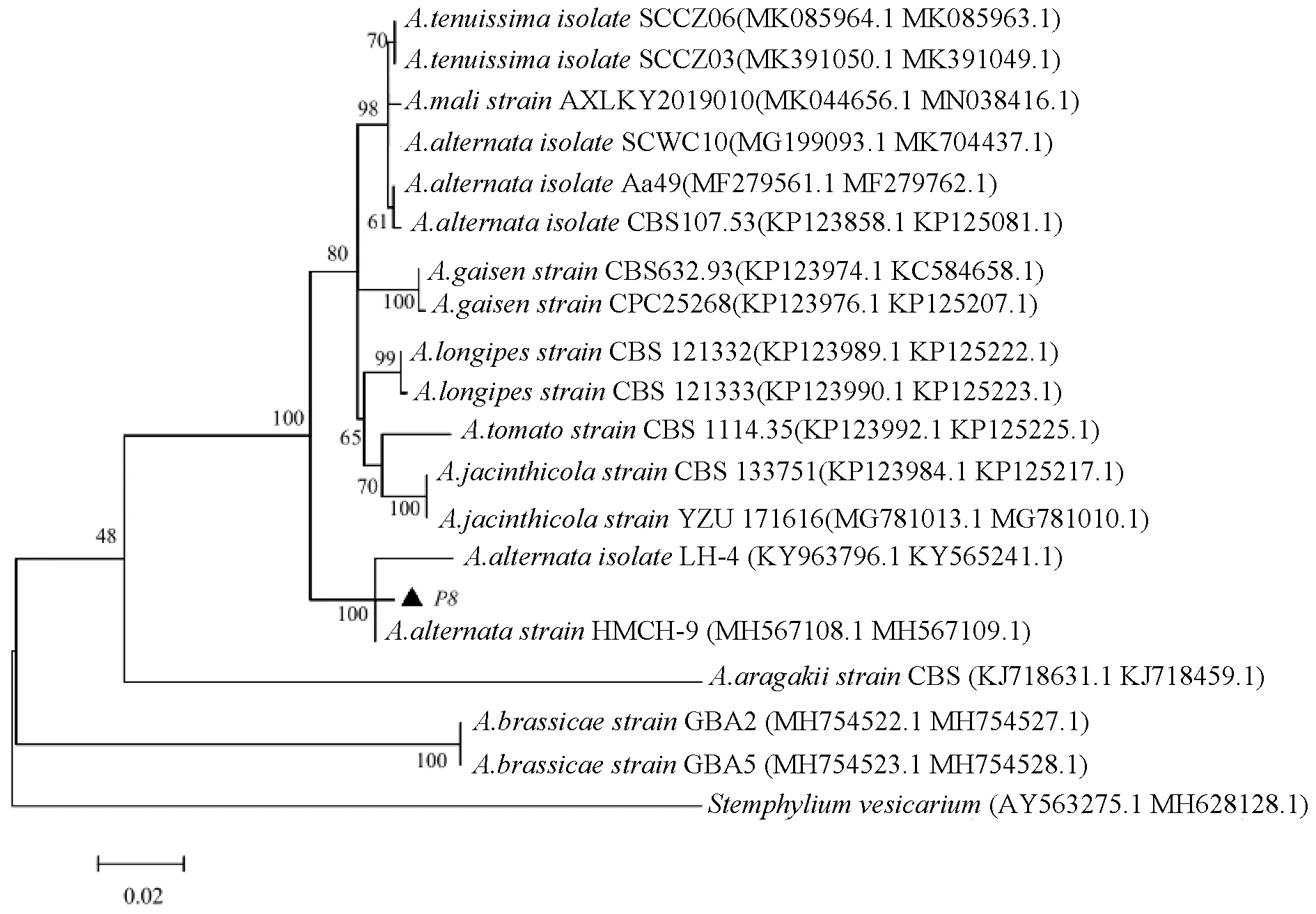
图4 基于过敏原基因和翻译延长因子基因构建的P8菌株与其他链格孢菌的系统发育树

图5 基于过敏原基因和钙调蛋白基因构建的P21菌株与其他链格孢菌的系统发育树
3 结论与讨论
(1)本研究结果表明,库尔勒香梨黑斑病的主要病原种类是交链孢(A.alternata)和细极链格孢(A.tenuissima),分离比例分别为23.0%和61.5%。这与我国其他地区报道的梨黑斑病病原种类的研究结果基本一致,严进等[4]从河北和山东鸭梨黑斑病上分离鉴定的病原种类是交链孢、细极链格孢和侵染链格孢(A.infectoria),其比例分别是41%、54.8%和1.6%;朱红艳[3]从湖北地区的梨黑斑病上分离鉴定的主要病原种类是交链孢和细极链格孢,梨黑斑链格孢(A.gaisen)所占比例较少(4.9%)。链格孢属真菌大多数种类兼性寄生于植物上,引起包括禾谷类作物、油料作物、果蔬作物及观赏植物在内的多种经济植物病害,造成田间和产后损失,是经济上非常重要的病原真菌[22],其中交链孢和细极链格孢寄主范围广泛,除引起梨树黑斑病外,在我国还侵染危害苹果、枣、蓝莓、菠萝、枸杞等果树[23-27],因此交链孢和细极链格孢是引起我国果树黑斑病的主要病原种类。
(2)尽管形态学特征在链格孢属真菌一些种的鉴定上存在难以界定的困难,但依然是种类鉴定不可替代的重要依据。本研究依据产孢方式和分生孢子形态将引起库尔勒香梨黑斑病的链格孢鉴定为交链孢和细极链格孢。在产孢方式上,交链孢以合轴式方式产孢,形成树状短分支的分生孢子链,孢子链一般由10个以内的孢子构成;细极链格孢的孢子链多为不分支的长链,一般由10个以上孢子构成。分生孢子形态差异表现在细极链格孢的成熟孢子中部有2~3个主横隔膜,明显较粗;且细极链格孢的分生孢子略大于交链格孢的分生孢子。因此形态学特征对交链孢和细极链格孢的种类鉴定非常重要。
(3)基于不同的单基因对同一个链格孢菌株进行系统发育分析时,由于所获结果往往不一致,因此越来越多的链格孢种类鉴定采用了多基因联合系统发育分析,如基于ITS,Alt-a1和gpd序列构建系统进化树,王彩霞[8]将引起葡萄叶斑病的链格孢鉴定为3个种;基于ITS、gapdh、rpb2、tef1、Alt-a1和endoPG序列,日本学者[10]将造成草莓黑斑病的链格孢鉴定为梨黑斑链格孢(A.gaisen)。本研究分别基于Alt-a1和Cal基因、Alt-a1和tef1基因构建系统进化树,将库尔勒香梨链格孢P8和P21菌株分别鉴定为交链孢和细极链格孢,并与形态学鉴定结果一致。利用基因序列对链格孢进行系统进化分析时发现,在NCBI上检索单个基因时可以获得很多相关参考基因,但对多个目标基因进行检索时,因不同的菌株可利用的参考基因不同,导致基于3~4个基因构建系统进化树时可利用的参考菌株很少,如本研究采用Alt-a1、Cal和tef13个基因构建系统进化树时,因具有相同目标基因的参考菌株很少,只能针对P8菌株基于Alt-a1和tef1基因搜索具有相应基因的参考菌株、针对P21菌株基于Alt-a1和Cal基因搜索具有相应基因的参考菌株,构建了2个系统进化树对2个菌株分别进行了系统发育分析,因此目前利用多基因进行系统发育分析还存在局限,随着今后有关链格孢的深度测序将有助该方法的广泛应用。
猜你喜欢
特产研究(2022年1期)2022-11-30
中国果业信息(2022年4期)2022-11-22
今日农业(2022年16期)2022-09-22
农民致富之友(2019年32期)2019-11-23
新疆人文地理(2018年5期)2018-11-27
南方农业·下旬(2016年9期)2017-04-27
江苏农业科学(2015年1期)2015-04-17
农产品市场周刊(2014年35期)2014-12-18
