
黄土高原雨养区苜蓿地土壤微生物功能多样性研究
2020-11-19 23:19:22程文文罗珠珠牛伊宁
河南农业科学 2020年11期
程文文,罗珠珠,牛伊宁,蔡 霞
(1.甘肃农业大学 资源与环境学院,甘肃 兰州 730070;2.甘肃省干旱生境作物学省部共建国家重点实验室,甘肃 兰州 730070)
微生物是土壤生态系统的重要组分[1],在参与土壤养分循环[2]、改善土壤结构[3]、有机质转换[4]以及维持生态系统的稳定方面具有重要的作用。研究表明,土壤微生物参与土壤反应的绝大部分进程[5],微生物的分布和活动是土壤环境与微生物群落相互影响与适应的结果[6]。微生物在一定程度上能反映土壤环境,更早地指示生态系统功能的变化,可以作为评价土壤环境质量的重要参数[7]。因此,微生物在农业土壤生态系统中的作用愈发突出。土壤微生物多样性的研究方法很多,GARLAND等[8]首次应用Biolog生态微平板描述微生物的群落功能特征,该方法通过检测土壤微生物对单一碳源的利用方式的差异,进而表征环境中各个类型的土壤微生物代谢等方面的功能多样性,用来预估评价土壤质量与不同耕作栽培措施之间的响应[9]。
黄土高原半干旱区是我国重要的雨养农业区,干旱少雨、水土流失严重是该区农业可持续发展的主要限制因素[10]。除了特殊的地形地势、黄土抗蚀能力差、植被覆盖度低、降雨分配不均等自然原因外[11],传统农业对土地的不合理利用是导致严重水土流失的人为原因之一[12]。黄土高原丘陵沟壑区长期以小麦(TriticumaestivumL.)为当地重要的粮食作物,以小麦单作为主的传统农业生产系统极易引起水土流失,导致土壤质量退化。紫花苜蓿(Medicagosativa)根系周围环境土壤中的微生物具有较好的根瘤固氮作用,同时由于苜蓿本身的生长特性,收割之后地上部分还拥有大量可以供给作物生长需要的残茬,地下部分大量的须根给土壤留下的腐殖质可增加土壤有机质[13],改善土壤团粒结构[14]。苜蓿在黄土高原半干旱区的可利用价值受到广泛关注,但是持续种植多年后苜蓿产量会下降,制约农业的绿色环保与可持续发展[15]。
目前,苜蓿连续种植多年导致的产草量下降和土壤干燥化现象已经引起重视,国内学者围绕苜蓿在不同生态区种植与发展对土壤水分变化特征的影响进行了深入研究[16-18],长期种植苜蓿对土壤理化性质的变化方面亦有报道[19-22]。但是,关于苜蓿土壤微生物研究相对较少。因此,依托黄土高原雨养农业区的长期定位试验,分析探讨土壤微生物多样性与苜蓿种植年限之间的关系,研究结果可用来评价苜蓿土壤肥力状况,做到健康管理土壤微生物,亦可以指导黄土高原雨养农业系统苜蓿人工草地的可持续利用,促进当地农业可持续发展。
1 材料和方法
1.1 研究区概况
试验依托设于位于温带干旱少雨地区的甘肃省定西市甘肃农业大学长期定位试验(2002—2016年)。年均太阳辐射592.9 kJ/cm2,年平均日照时长2 476.6 h,年均降水390.9 mm,地面降水年蒸发量1 531 mm,日平均气温6.4 ℃,满足作物生长的有效积温2 239.1 ℃,全年无霜期140 d。
1.2 研究方法
1.2.1 试验设计 选取种植年限分别为6、13、15 a的紫花苜蓿人工草地和农田为研究对象(处理),不同种植年限苜蓿生长期间均未施肥、灌水,且各小区面积均为21.0 m2(3.0 m ×7.0 m),不同种植年限的苜蓿取3次重复,得出各处理的平行值。
1.2.2 土壤样品的采集与测定 于苜蓿盛花期,用5点取样法在每个测定的试验小区作物行间采集0~30 cm和30~60 cm土壤深度样品,并剔除土样中的植物根系、落叶、石块等杂物,随后将土样混合均匀,过筛(0.25 mm)。将用于测定土壤微生物量碳(MBC)含量和微生物功能多样性的土样放置在4 ℃冰箱中避光保存;同时预留配套的土样,风干研磨之后,用于测定土壤有机碳(SOC)含量。
SOC含量采用外加热重铬酸钾法测定[23],MBC含量采用氯仿熏蒸浸提法测定[24]。使用Biolog生态微平板(Biolog eco plate TM)测定土壤微生物功能多样性等相关指标[25]。
1.2.3 计算方法 平均颜色变化率(AWCD)值可以解释土壤微生物对碳源利用程度的差异,是衡量微生物代谢活性的指标。计算公式如下:
AWCD=∑(Ci-R)/n
(1)
式中:Ci为各反应孔在590、750 nm下的光密度值之差;R为生态微平板对照孔的光密度值;在计算中,Ci-R为负值时,该孔的结果记为0,即Ci-R的值均大于或等于0[26];n=31。
土壤微生物群落功能多样性可以借助Shannon-Wiene物种多样性指数(H)、均匀度指数(U)和碳源利用丰富度指数(S)来表征,计算公式如下[27-28]:
H=-∑Pi×ln(Pi)
(2)
(3)
式中:Pi为第i孔的相对吸光值与平板所有反应孔相对吸光值总和的比率,即Pi=(Ci-R)/∑(Ci-R);ni为第i孔的相对吸光值;S采用反应孔(吸光值>0.2,则代表该孔的碳源被利用,该孔为反应孔)的数目表示[29-30]。
将Biolog生态微平板中95种碳源分为6类,反映土壤微生物群落利用各类碳源的特征,其中,各类碳源占碳源总数的比例即碳源利用率。
1.2.4 主成分分析(PCA) 载荷因子反映了微生物对土壤碳源利用情况和所提取主成分之间的相关性。载荷因子是正的,说明两者正相关,反之亦然。碳源的初始载荷因子绝对值越大,说明该碳源对主成分的影响越大。本研究选取荷载因子绝对值大于0.5的碳源作为与主成分1(PC1)和主成分2(PC2)显著相关的碳源[31]。
1.3 数据分析
采用Excel 2003进行数据统计,用Origin 8.0制图,用SPSS 21.0软件进行方差显著性分析、主成分分析。利用主成分分析衡量微生物对不同碳源的利用情况[32]。
2 结果与分析
2.1 不同年份苜蓿种植和农田土壤SOC、MBC含量
由图1可知,随着土壤深度的增加,SOC与MBC含量均呈现下降趋势。SOC含量在0~30 cm土壤深度内表现为13 a>6 a>农田>15 a,13 a与6 a之间SOC含量差异不显著,但是与15 a之间差异显著;SOC含量在30~60 cm土壤深度下呈现出6 a>农田>15 a>13 a,6 a与13 a以及15 a之间差异显著,13 a与15 a之间差异不显著,6 a SOC含量比13 a提高48.08%,比15 a提高45.84%。在土壤深度0~60 cm内,MBC含量均呈现出6 a>13 a>15 a>农田,6 a与13、15 a之间差异显著,13 a与15 a之间差异不显著,6 a比13 a提高37.47%~38.16%,比15 a提高47.81%~54.40%。
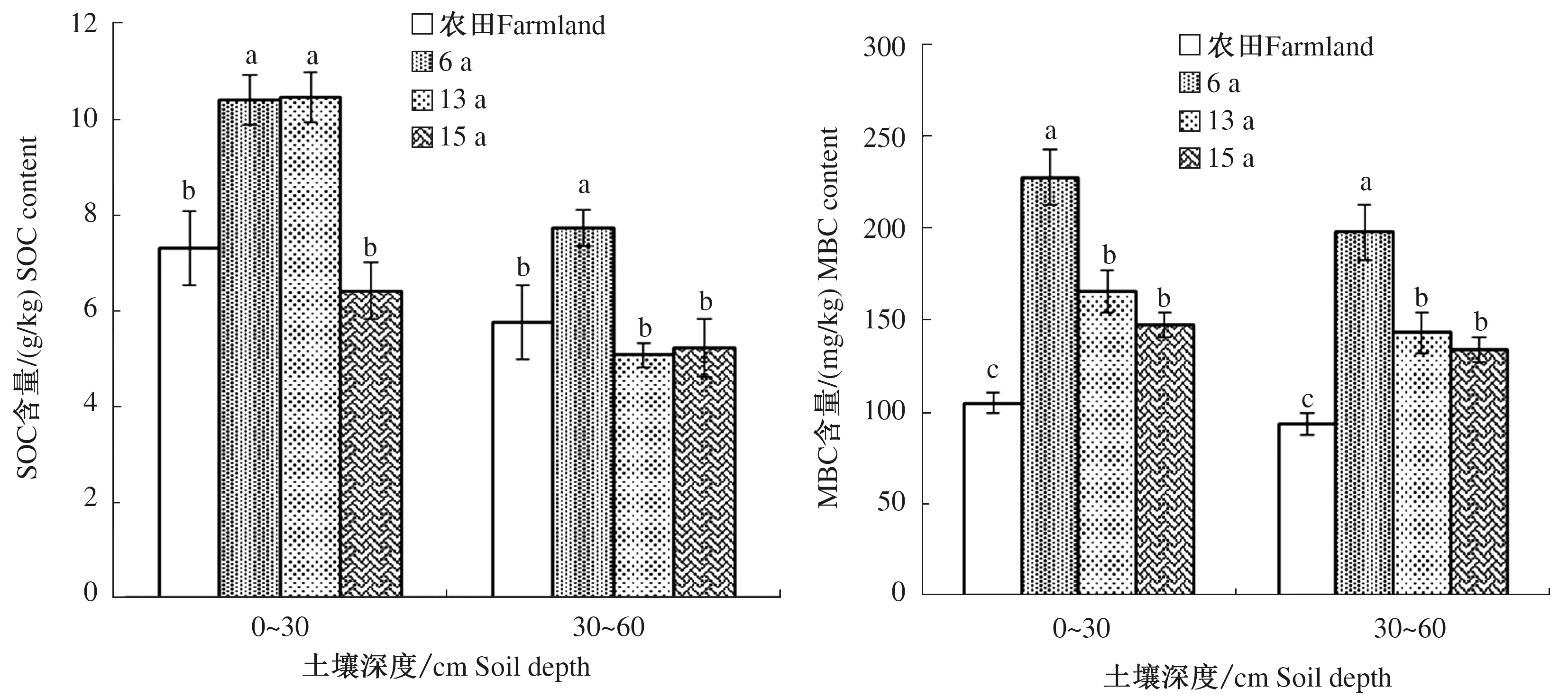
不同小写字母分别表示相同土壤深度下不同处理间差异显著(P<0.05),下同
2.2 不同年份苜蓿种植和农田土壤微生物AWCD值
由图2可知,AWCD值随着培养时间的延长而增加。各处理的AWCD值在0~24 h内几乎无变化,表明此时间段内微生物基本未利用碳源,未开始自身的代谢活动。在24 h之后,微生物开始逐步利用碳源,AWCD值迅速提高,微生物进入迅速增长时期,进入一种理想化的增长模式,即为生态学上所提及的对数增长期,呈“J”形增长曲线(24~144 h)。当144 h之后,曲线开始渐渐的趋于平滑。不论何种处理,随着土壤深度的加深,AWCD值降低。同时发现,土壤深度0~30 cm时,培养时间120~192 h时,AWCD值表现为6 a最高,农田和13 a居中,15 a较低,体现了较强的碳源的利用能力;土壤深度30~60 cm时,AWCD值整体表现为6 a最高,13、15 a居中,农田最低。
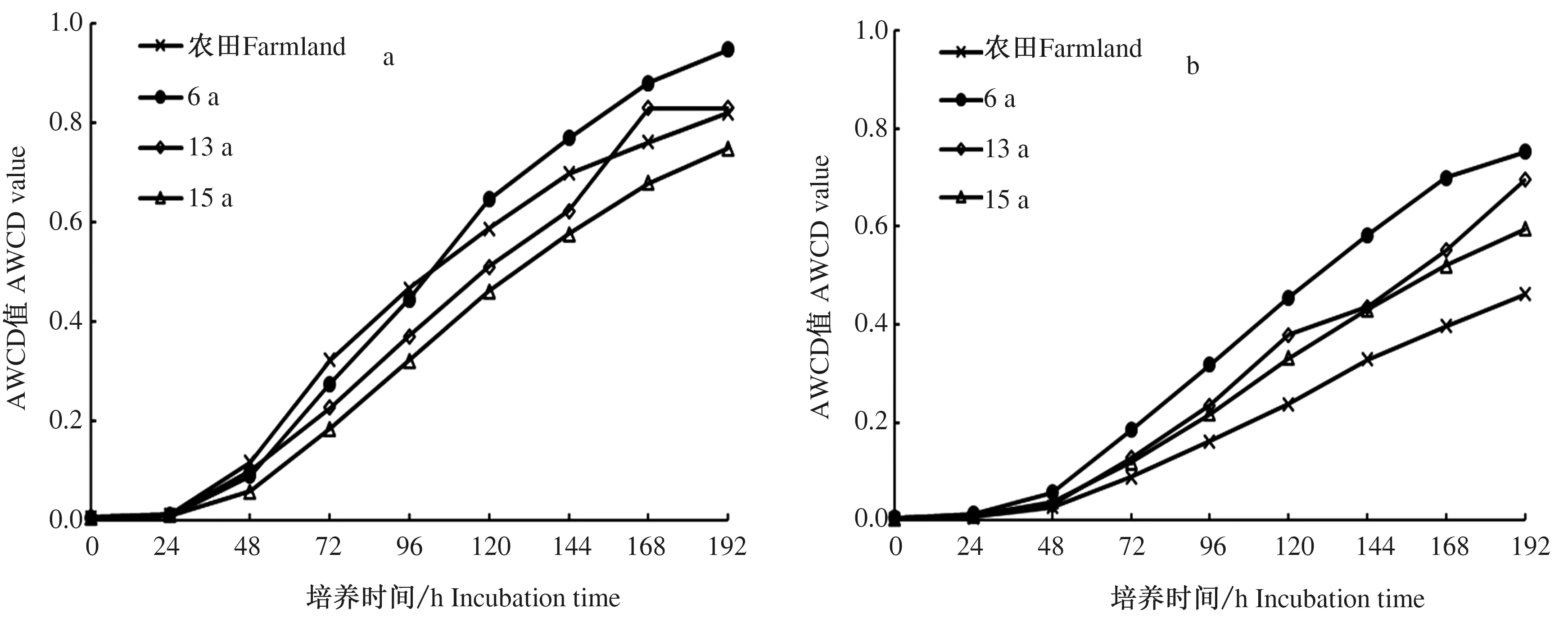
a:土壤深度0~30 cm;b:土壤深度30~60 cm,下同a:Soil depth af 0—30 cm; b:Soil depth of 30—60 cm,same below
2.3 不同年份苜蓿种植和农田土壤微生物碳源利用率
由图3可知,苜蓿种植土壤和农田土壤微生物对碳源利用情况有所不同,土壤深度0~30 cm和30~60 cm,农田土壤微生物碳水化合物类(50.68%、51.53%)、羧酸类(21.01%、21.89%)利用率较高;苜蓿种植土壤碳水化合物类和氨基酸类利用率较高,其中,土壤深度0~30 cm,碳水化合物类和氨基酸类利用率分别为40.50%~45.54%、20.66%~23.61%,土壤深度30~60 cm,碳水化合物类和氨基酸类利用率分别为38.90%~41.45%、19.66%~21.31%。种植苜蓿降低了土壤微生物对碳水化合物类和羧酸类的利用率,增加了氨基酸类、聚合物类、酚酸类和胺类的碳源利用率。同时发现,农田土壤微生物所利用碳源的一半以上来源于碳水化合物类,土壤深度30~60 cm酚酸类和胺类(0.44%、0.92%)利用率较少,这可能是由于农田土壤微生物类群相对单一所致。
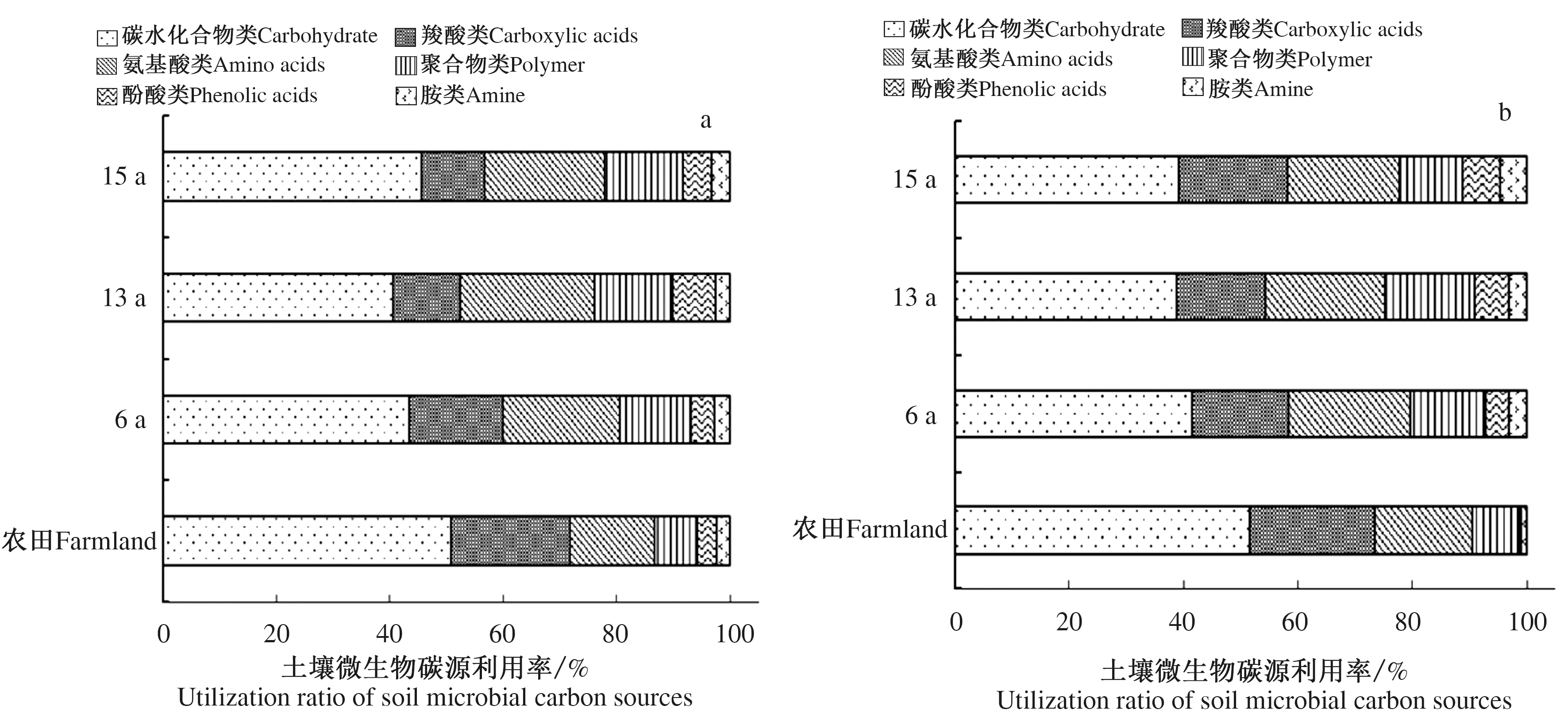
图3 苜蓿种植和农田土壤微生物碳源利用率 Fig.3 Utilization ratio of soil microbial carbon sources of farmland and alfalfa planting soil
2.4 不同年份苜蓿种植和农田土壤微生物群落功能多样性指数
由表1可知,土壤微生物培养120 h,H、U和S以及AWCD值均随土壤深度增加而逐渐变小,表明微生物活动的代谢能力减弱。土壤深度0~30 cm,H、U、S值处理间差异不显著;土壤深度30~60 cm,H、S值均表现为6 a最高,其中,6、13 a的S值与15 a之间差异显著,苜蓿种植土壤与农田土壤间差异显著。各处理AWCD值在0~30 cm土壤深度无显著差异;30~60 cm土壤深度表现为6 a最高,与农田土壤差异显著。同时可以看出,随着苜蓿种植年限的增加,各土层H、U值均减小,表现为6 a>13 a>15 a。
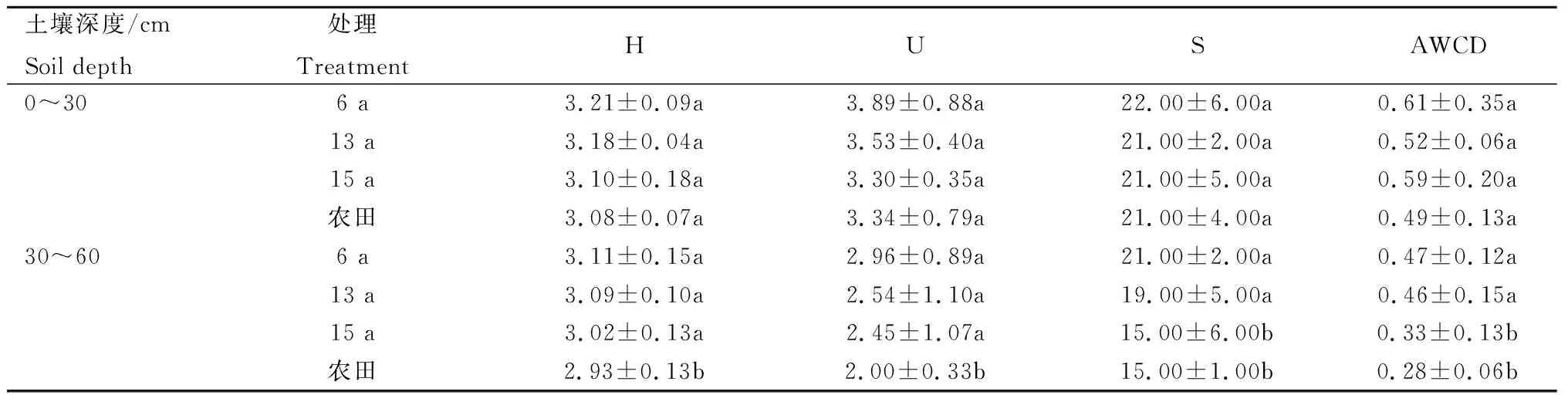
表1 苜蓿种植和农田土壤微生物群落功能多样性指数Tab.1 Functional diversity index of soil microbial communities of framland and alfalfa planting soil
2.5 不同年份苜蓿种植和农田土壤微生物群落功能多样性的主成分分析
用PCA法对土壤微生物群落的碳源利用情况分析,了解不同苜蓿种植年限土壤微生物群落功能多样性。根据此原则,在0~30 cm土壤深度的31个因子中提取了7个主成分,累计贡献率为93.90%,其中PC1为48.26%,权重最大,PC2次之,为14.60%,PC3—PC7较小;在30~60 cm土壤深度提取了8个主成分,累计贡献率为95.59%,其中,PC1为35.98%,权重最大,PC2次之,为17.73%,PC3—PC8较小。可见,前2个主成分可以用来解释土壤微生物群落功能多样性差异变化。
由图4a可知,在PC1轴上,虽然各处理正负方向均有分布,但6 a主体在正方向,农田、13 a和15 a主体在负方向;在PC2轴上,农田分布在负方向,得分-0.29~-3.44,其余处理基本分布在正方向,得分-1.68~5.56。由图4b可知,在PC1轴上,6 a均分布在负方向,农田和13 a均分布在正方向,15 a虽然正负方向均有分布,但主体基本分布在负方向;在PC2轴上,6 a均分布在负方向,农田虽然正负方向均有分布,但主体基本分布在负方向,13 a和15 a均分布在正方向。从图4a可以看出,0~30 cm
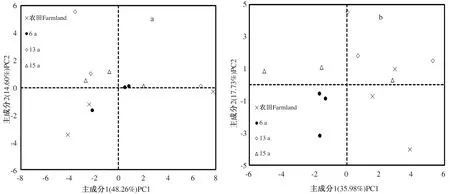
图4 苜蓿种植和农田土壤微生物碳源利用主成分分析Fig.4 Principal component analysis of soil microbial carbon sources of farmland and alfalfa planting soil
土壤深度农田投射点分布在第三、四象限,其余处理投射点大多分布在第一、二象限;从图4 b可以看出,30~60 cm土壤深度6 a投射点分布在第三象限,农田投射点主要分布在第四象限,其余处理投射点均分布在第一、二象限。这表明不同处理土壤微生物群落功能多样性出现分异,对各类碳源的喜好不一。
从表2可知,在0~30 cm土壤深度,PC1载荷因子绝对值在0.5以上的有14种,主要包括碳水化合物类(4种)、氨基酸类(4种)、羧酸类(4种)、胺类(1种)、酚酸类(1种),PC2载荷因子绝对值在0.5以上的仅有5种,包括碳水化合物类(1种)、氨基酸类(1种)、羧酸类(1种)、酚酸类(1种)、聚合
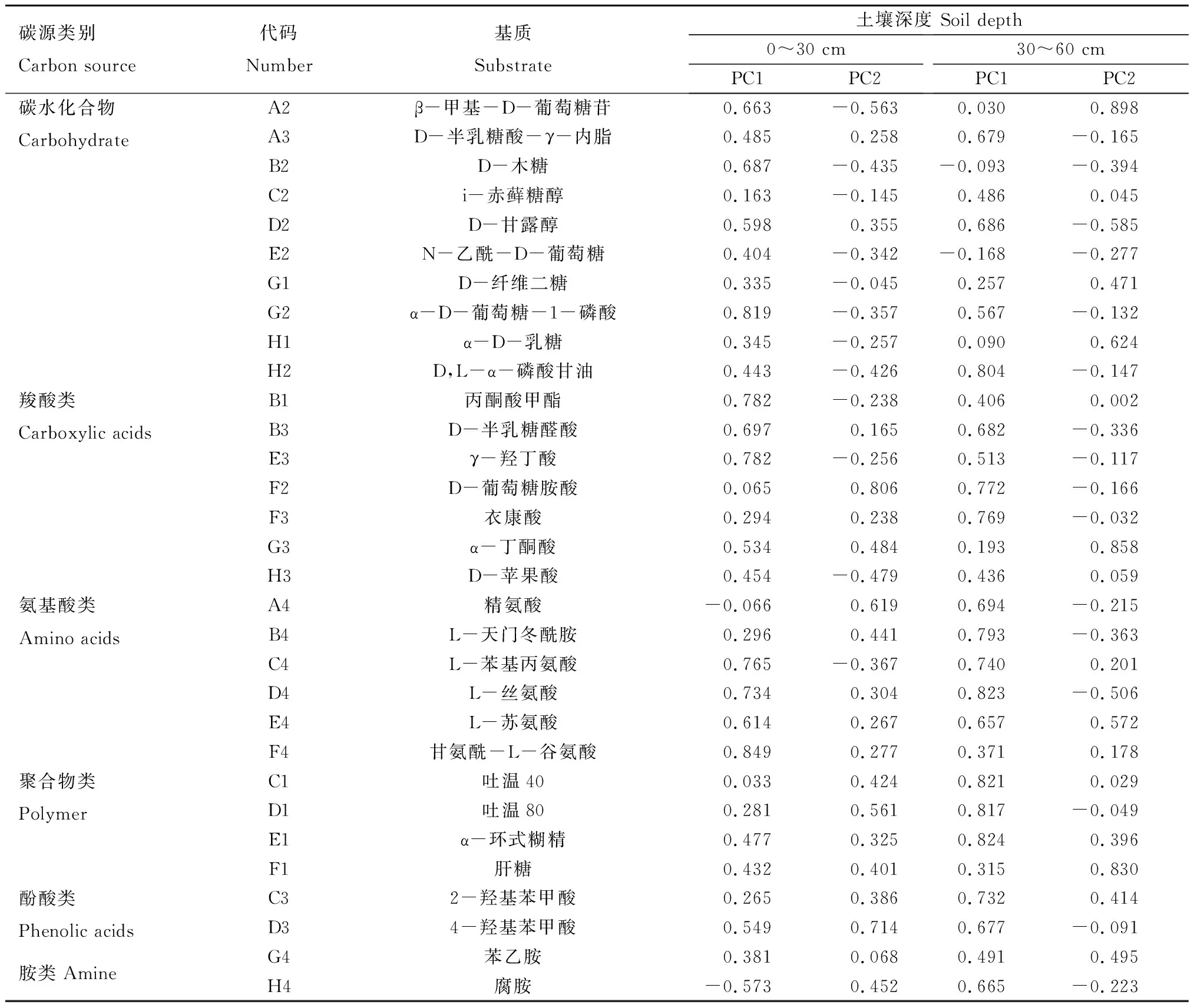
表2 31种碳源的主成分分析载荷因子Tab.2 Loading factors of principle components of 31 substrates sole-carbon sources
物类(1种)。说明碳水化合物类、氨基酸类、羧酸类是0~30 cm土壤深度土壤微生物利用的主要碳源类型。在30~60 cm土壤深度,PC1载荷因子绝对值在0.5以上的碳源有19种,主要包括碳水化合物类(6种)、氨基酸类(5种)、聚合物类(3 种)、羧酸类(2种)、酚酸类(2 种)、胺类(1种);PC2载荷因子绝对值在0.5以上的碳源仅有7 种,包括碳水化合物类(3种)、氨基酸类(2种)、聚合物类(1种)、羧酸类(1种)。说明碳水化合物类、氨基酸类、聚合物类是30~60 cm土壤深度土壤微生物利用的主要碳源类型。
3 结论与讨论
土壤碳组分是评价土壤环境质量的重要指标之一,其在改善土壤结构、减轻水土流失、维持植物营养的有效性方面具有重要作用。不同作物类型下土壤由于受来自植物的凋落物[33]、根系分泌物[34]等物质的质量和数量不同[35],土壤有机碳组分分布情况不同。本研究中,SOC、MBC含量均表现为6 a显著高于15 a,由于紫花苜蓿为多年生植物,随着种植时间延长,地上生物量逐渐增加,进而导致枯枝落叶和根系分泌量增加,提高了土壤中的SOC和MBC含量,为微生物的生长与繁殖提供了充足的碳源[13-14],当苜蓿种植超过一定年限后,土壤环境恶化,板结严重,土壤孔隙度降低,不再适宜于微生物的生存与繁殖,此外地上生物量减少,使得固定到土壤中的碳素减少,呈现出MBC含量降低的趋势[36]。
AWCD值反映土壤微生物利用碳源的能力和代谢活性的大小[37-38],在一定程度上能反映土壤中微生物种群的数量和结构特征[39]。AWCD值的结果表明,在培养中期(24~144 h),土壤微生物对碳源利用强度不断增强。培养后期(144 h之后),微生物对碳源的利用趋于稳定状态,存在较明显的适应期、对数期和稳定期等阶段,符合一般微生物利用基质的规律[40]。本研究发现,伴随土壤深度的增加,AWCD值降低,微生物自身代谢活动受到抑制。苜蓿为多年生草种,根系发达,随着苜蓿种植时间的延长到达旺盛期后地表覆盖物不断增加,凋落物和根系分泌量增加,促进微生物繁殖,此时苜蓿根瘤菌的固氮作用较强,使土壤有机碳和养分含量增加[41-42],影响土壤微生物代谢活性。但当苜蓿种植年限达到一定的阶段,土壤板结问题日益严重,导致土壤中的孔隙度降低,空气含量亦下降,生存条件恶化导致微生物总体活性下降[36]。而农田土壤作物根系不发达,微生物活性明显弱于苜蓿地,使其土壤微生物代谢活性低于苜蓿土壤。
H、U、S值能代表土壤微生物利用碳源的能力以及多样性,是分析群落功能多样性的重要指数[43]。H值越高,表明所含物种越丰富;U值越高,表明群落均一程度越高;S值越高,表明可利用碳源种类越多[44]。本研究发现,随着苜蓿种植年限增加,H、U值均呈下降趋势,0~30 cm和30~60 cm土壤深度2种指数并未表现出明显差异;但30~60 cm土壤深度, H、S值表现为6、13 a与农田差异显著。苜蓿属密集型草本,地面附着物效果好,可保水,长期种植苜蓿且未翻耕,植物残体落叶等进入土壤转化为腐殖质,使有机物质含量提高,为微生物的生长和繁殖提供了充足的底物[45],因此土壤微生物丰富度增加。苜蓿种植年限长达15 a以后开始减产,养分含量降低与生长衰退同时发生[36],有机物质的减少使微生物丰富度随之减少。
植被通过影响土壤环境而改变土壤微生物生长环境,土壤微生物碳源代谢类型与作物种植体系以及耕作措施有显著的相关性[46]。本研究发现,种植苜蓿降低了土壤微生物对碳水化合物类和羧酸类的利用率,提高了氨基酸类、聚合物类、酚酸类和胺类的利用率。同时发现,农田土壤微生物利用碳水化合物类较多,但对聚合物类、酚酸类和胺类的利用率相对较低,特别是土壤深度30~60 cm。这种差异主要源于不同种植体系引起土壤性质发生改变,土壤微生物与土壤团聚体数量及质量之间存在密切联系,而土壤团聚体的改变与土壤环境微生物的迁移与代谢过程紧密相关[47]。不同种植年限苜蓿土壤长期以来一直未进行耕作扰动,而农田土壤由于种植一年生粮食作物需要连年耕作,耕作后土壤团聚体数量和直径远小于未耕作的土壤[48],耕作措施改变土壤结构进而影响到土壤微生物群落特征[47]。而且,不同作物对土壤微生物群落的影响不同[49],不同作物的根系分泌物可能使土壤微生物对某些碳源的利用能力有一定的抑制作用,使某些碳源利用能力消弱,这与有利于利用此类碳源的土壤微生物数量减少有一定关系。
主成分分析可用来解释不同处理土壤微生物碳源利用是否存在差异。土壤深度0~30 cm和30~60 cm各处理土壤微生物对碳源的利用出现了较好的分离,碳源的代谢表现出明显变化。6 a比较集中在某一特定区域,而其他2个种植年限分布较分散,说明6 a土壤微生物代谢功能性较好,对碳源的利用情况较稳定。不同种植年限苜蓿改变土壤微生物群落功能多样性是由微生物对PC1和PC2上载荷较高的碳源利用差异引起的,土壤深度0~30 cm在PC1和PC2上载荷较高的主要碳源为碳水化合物类、氨基酸类、羧酸类;土壤深度30~60 cm在PC1和PC2上载荷较高的主要碳源为碳水化合物类、氨基酸类、聚合物类,说明这3类碳源可作为不同种植年限土壤微生物碳源利用的敏感碳源。
Biolog生态微平板方法只能检测土壤中快速生长或富营养微生物的活性,而不能用来解释绝大部分土壤中其他类型的微生物活性[31],因此采用Biolog生态微平板方法来研究土壤微生物群落的代谢及相关生理生化过程可能存在一些偏差。因此,今后的研究应进一步将Biolog生态微平板法与16S rDNA方法结合起来,并结合冗余分析(RDA)和典型相关分析(CCA)等方法,以期为黄土高原雨养农业系统苜蓿草地的可持续利用与当地农业的发展提供科学依据。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05 07:20:20
现代畜牧科技(2021年9期)2021-10-13 06:38:52
现代畜牧科技(2021年4期)2021-07-21 06:13:04
昆钢科技(2021年6期)2021-03-09 06:10:20
少儿科技(2021年8期)2021-01-02 10:01:24
婚姻与家庭·性情读本(2019年11期)2019-11-18 01:48:46
保健与生活(2019年7期)2019-07-31 01:54:07
中国三峡(2017年4期)2017-06-06 10:44:22
为了孩子(3~7岁)(2016年6期)2016-05-14 03:09:17
电源技术(2016年9期)2016-02-27 09:05:25
