
苹果树腐烂病菌VmSom1 基因克隆与序列分析
2020-11-02 07:23王怡霖孙庚午刁雨菲冯军利何邦令刘会香
山东农业大学学报(自然科学版) 2020年5期
王怡霖,赵 涛,孙庚午,刁雨菲,冯军利,于 涛,何邦令*,刘会香*
1.山东农业大学植物保护学院/山东省林业有害生物防控工程技术研究中心,山东 泰安 271018
2.泰安市徂徕山林场,山东 泰安 271027
3.淄博市淄川林业局,山东 淄博 255100
苹果树腐烂病(Valsa canker of apple)是由苹果黑腐皮壳菌(Valsa mali)引起的一种严重枝干病害,分布广,危害严重[1]。开展苹果树腐烂病菌致病相关机制的研究对于科学防控病害具有重要意义。目前苹果树腐烂病菌致病相关因子主要包括降解酶类物质、毒性次生代谢物、分泌蛋白、转录因子等[2-5]。降解酶类物质主要包括果胶酶和根皮苷降解酶等[6],毒性次生代谢物包括聚酮合成酶、二甲基丙烯基色氨酸合成酶、非核糖体多肽合成酶和异香豆素类物质等[6],分泌蛋白、转录调控因子如HMG[8]、VmSeb1[9]、VmPacC[10]、VmPxE1[11]、天鹅绒蛋白(Velvet protein)家族中的VmVe[12]以及VmPoDs[6]、VmXyl1[7]、VmPmk1[13]、VmE02[14]、VMAGO2[6]和编码异源三聚体G蛋白α亚基的基因Gvm2和Gvm3[15]等,上述致病相关因子在病菌的生长发育和致病过程中均发挥重要作用。
Som1(cAMP-dependent protein kinase pathway protein)是位于环化腺苷酸信号途径(cAMP-PKA)下游的重要转录因子。目前,对于Som1基因在酵母(Saccharomyces cerevisiae)中的同源基因flo8的研究较为深入,flo8可与转录抑制子Sfl1 共同调控酵母细胞间的粘附力,使酵母牢固附着并穿透培养基[16]。在丝状真菌中构巢曲霉(Aspergillus nidulans),Som1的同源基因SomA调控真菌菌丝形态建成和致病性,烟曲霉中SomA基因的缺失导致生长缓慢和无性发育的阻滞,只能形成没有进一步分化的气生菌丝,且SomA是生物膜形成所必需的[17]。在Magnaporthe oryzae中,MoSom1在孢子和附着物的发育是必需的,并且在孢子形成期间在细胞壁分化,调节黑色素沉着和细胞表面疏水性中起作用,对于亚细胞定位和生物学功能是必需的[18,19]。大丽轮枝菌(Verticillium dahliae)中,Som1调控真菌粘附和根部穿透、控制隔膜定位和空泡的大小,随后控制菌丝形成,包括气生菌丝形成和正常菌丝分支,致病性所需的发育遗传网络[20]。在模式昆虫病原真菌(Metarhizium acridum)中,Som1的同源基因MaSom1的缺失降低了分生孢子产量,延迟了分生孢子萌发,并削弱了对热和UV-B的真菌耐受性,降低了真菌的毒力。此外,MaSom1对于细胞壁完整性和分生孢子表面结构也有重要作用[21]。
在实验室前期已构建的Valsa maliATMT(Agrobacterium tumefaciensmediated transformation,ATMT)突变体库基础上[22],通过转化子表型筛选,获得了一突变株V23。应用hiTAIL-PCR方法分离克隆了侧翼序列,获得了VmSom1突变基因。本研究分析了基因序列和编码蛋白功能域特征,研究结果为进一步明确该基因功能奠定了基础。
1 材料与方法
1.1 实验材料
苹果树腐烂病菌菌株sdau11-175由本实验室分离保存;随机挑选pKO1-HPH载体所构建的ATMT转化子50个[22],所有转化子均由本实验室获得并在20%甘油4 ℃低温保存;引物合成及序列测定由北京华大基因科技有限公司完成;大肠杆菌(Escherichia coli)DH5α感受态细胞、pMD18-T载体购自宝生物工程(大连)有限公司、克隆载体pEASY-T1购自北京全式金生物有限责任公司。
1.2 突变体筛选
筛选以转化子菌落生长速率、培养学特性和致病性差异显著性为评价指标。以野生型菌株sdau11-175为参照菌株,在25 ℃PDA培养基上培养,十字交叉法测定5 d的菌落生长直径,计算菌落生长速率,实验重复5次;同时记录菌落形状、颜色和气生菌丝发达程度;准备成熟度和大小一致红富士苹果,采用五点刺伤接种法,每个苹果接种4个点,每个转化子共接种12个点,接种后使用保鲜膜保湿,置于25 ℃黑暗培养,重复3次,7 d后测定病斑直径,数据处理采用SPSS方法(P<0.05)[8]。
1.3 突变体侧翼序列克隆
以上述筛选出的突变体为目标菌株进行基因组总DNA提取,突变体T-DNA插入位点左右的侧翼序列扩增采用hiTAIL-PCR方法[23]。L(R)B2不添加5’端接头。在一级反应中,使用的模板是突变体菌株的基因组总DNA,在二级和三级反应中,使用模板则分别为上一级反应产物稀释1000倍。产物在1.5%琼脂糖凝胶中电泳,以1 kb Maker为参照,将PCR特异性产物经回收、连接、大肠杆菌转化,菌落PCR检测和华大基因测序后得到的侧翼序列进行NCBI Blastx在线分析,使用BioEdit软件结合G enbankValsa mali03-8基因组序列进行本地分析[4],获得两端侧翼序列全长。引物序列见表1,反应程序见[24]。

表1 本研究hiTAIL-PCR 引物Table 1 Primers of hiTAIL-PCR in this research
1.4 突变基因全长序列克隆
依据上述两端侧翼序列设计引物(F1-ATGAATAACGTCA ACATGGCCAATATGGA,R1-TCAC CGGACCCTAAGTCTCTATCTCCG),分别以野生型菌株sdau11-175和突变株基因组总DNA为模板进行PCR扩增,获取突变基因和含有T-DNA的突变基因全长序列。同时与已报道的V.mali03-8菌株的基因组序列进行比对分析,以确定T-DNA的插入位点及方式。
1.5 突变基因序列分析
使用NCBI的在线工具ORF FINDER和Graphics分别确定VmSom1的开放阅读框和内含子;在Genbank下载Neurospora crassa、Aspergillus nidulans、Magnaporthe oryzae、Aspergillus fumigatus、V.mali03-8、Candida albicans、Saccharomyces cerevisiae、Phaeoacremonium minimum、Diaporthe ampelina、Colletotrichum orbiculare、Colletotrichum gloeosporioides、Colletotrichum salicis、Eutypa lata、Verticillium alfalfae、Madurella mycetomatis、Ophiocordyceps sinensis、Ustilaginoidea virens、Fusarium fujikuroi、Metarhizium guizhouense以及Valsa malivar.Pyri中与sdau11-175VmSom1基因同源性高的序列,用MEGA 6采用Neighbor-Joining method方法对突变基因进行1000次置信度自展系统进化分析;使用ExPASy-ProtParam在线工具对突变基因编码蛋白质的理化性质进行分析;使用ExPASy-ProtScale在线工具对突变基因编码蛋白质的疏水性进行分析;使用在线SOPMA和Phyre2对突变基因编码蛋白的二级结构进行分析。应用在线软件CDD-search (https://www.ncbi.nlm.nih.gov)和http://smart.emblheidelberg.de/index2.cgi对蛋白功能域进行预测。
2 结果与分析
2.1 突变体筛选
与野生型菌株sdau11-175相比,随机挑选的50个苹果树腐烂病ATMT转化子中有49个生长速率、菌落特征与致病力差异不显著。转化子V23表现出生长速率显著减慢和致病力完全丧失的特征(图1)。同时V23菌落表面比较湿润,气生菌丝生长弱、不产生子实体(图2)。说明突变体V23调控的基因对苹果树腐烂病菌的生长发育及致病性有重要作用。
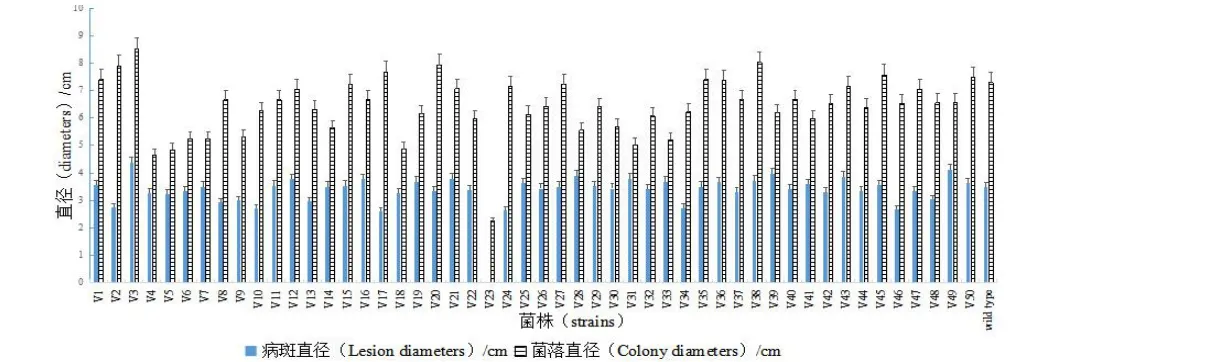
图1 苹果腐烂病菌转化子的生长速率和致病性测定Fig.1 Growth rates and pathogenicity characters of transformants of pathogen
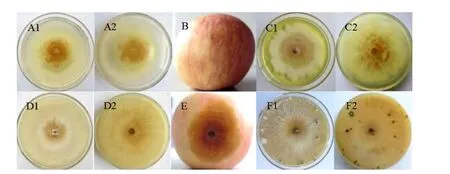
图2 突变体V23 和野生型sdau11-175 菌落形态、子实体产生和致病性测定结果Fig.2 Colony morphology,fruiting body generation and pathogenicity of mutant V23 and wild-type SDAU11-175
2.2 突变体侧翼序列及全长序列的克隆
以突变体V23基因组DNA为模板,应用hiTAIL-PCR方法进行T-DNA插入侧翼序列克隆。在突变体V23的T-DNA左边界L3得到了约1900 bp的特异性片段,在右边界R3得到了约2200 bp的特异性片段。V23突变体侧翼序列不同步骤PCR扩增结果见图3。以V.malisdau11-175基因组DNA为模板,使用两端侧翼引物序列克隆全长突变基因,其大小为2723 bp。
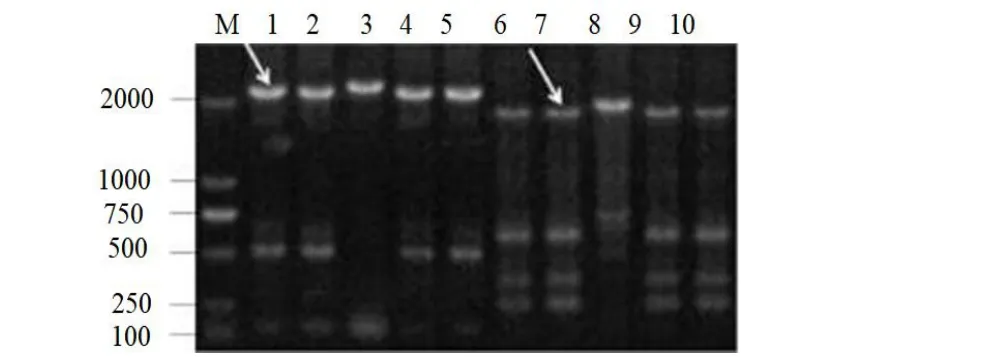
图3 突变体V23 的侧翼序列克隆Fig.3 Cloning of flanking sequences of V23 mutants
2.3 突变基因序列分析
2.3.1 突变基因核苷酸序列分析 突变基因编码区长2475 bp,编码824 个氨基酸,基因组DNA 含有5 个外显子和4 个内含子。将突变基因全长序列和V.mali03-8 序列比对,并将二者的氨基酸序列比对,去除密码子简并性因素,二者的氨基酸完全一致,说明在同一种真菌中该突变基因编码的蛋白相对保守。突变体V23 中的T-DNA 插入位点位于V.mali第12 号染色体(CM003109.1)的69537与69547 之间,在编码假定蛋白的基因VMIG_09677(GenBank:KUI74346.1)的第一个外显子中(图4)。被注释为编码蛋白Adhesion defective protein 3,该基因与Som1同源,故将其命名为VmSom1,在V.mali03-8 基因组中比对发现,VmSom1为单拷贝。

图4 突变体T-DNA 侧翼序列测序及T-DNA 插入位点Fig.4 T-DNA flanking sequences and insertion position of mutants
系统进化分析发现(图5),苹果腐烂病菌VmSom1基因与其他20种真菌中的Som1及其同源基因的亲缘关系远近各不相同,其中与同属真菌(梨腐烂病菌)真菌内Som1基因编码的氨基酸序列相似性很高,同源性高达98.79%;与不同属真菌(如稻瘟病菌、烟曲霉、构巢曲霉等)差异较大,蛋白质的特异性及功能主要取决于一级结构,推测不同属真菌的Som1蛋白功能可能存在一定差异。
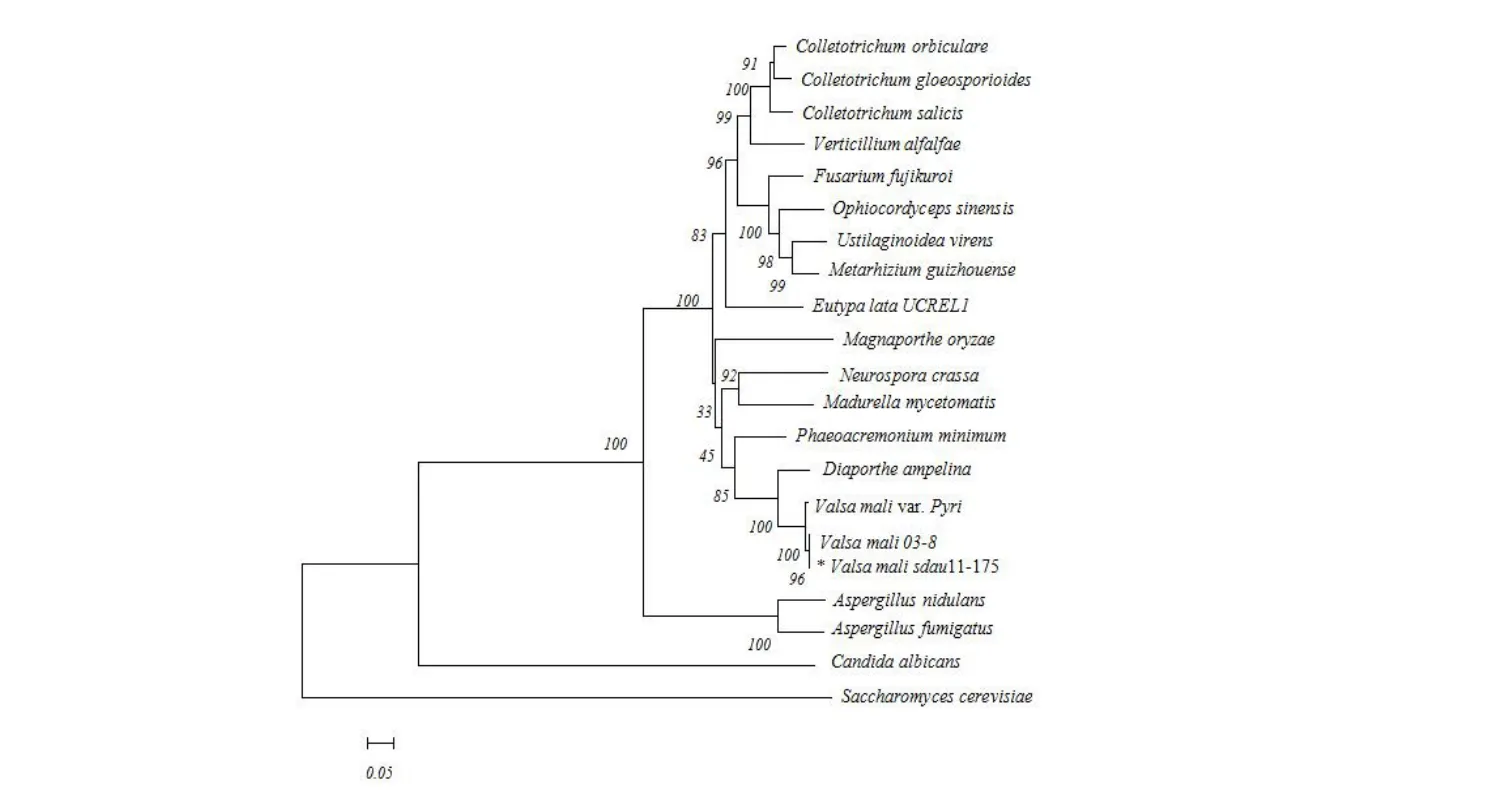
图5 苹果腐烂病菌VmSom1 蛋白的NJ 系统进化分析Fig.5 NJ Phylogenetic analysis of VmSom1 protein of Valsa mali
2.3.2 突变基因编码蛋白的功能域预测Valsa mali与Magnaporthe oryzae、Aspergillus fumigatus中均存在LisH 结构域和核定位信号肽(NLS)结构域,其中VmSom1 蛋白的LisH 结构域位于N 端43-75 aa,长为33 aa,同时在259-272 aa 处存在着核定位信号肽NLS,长为14 aa(图6)。
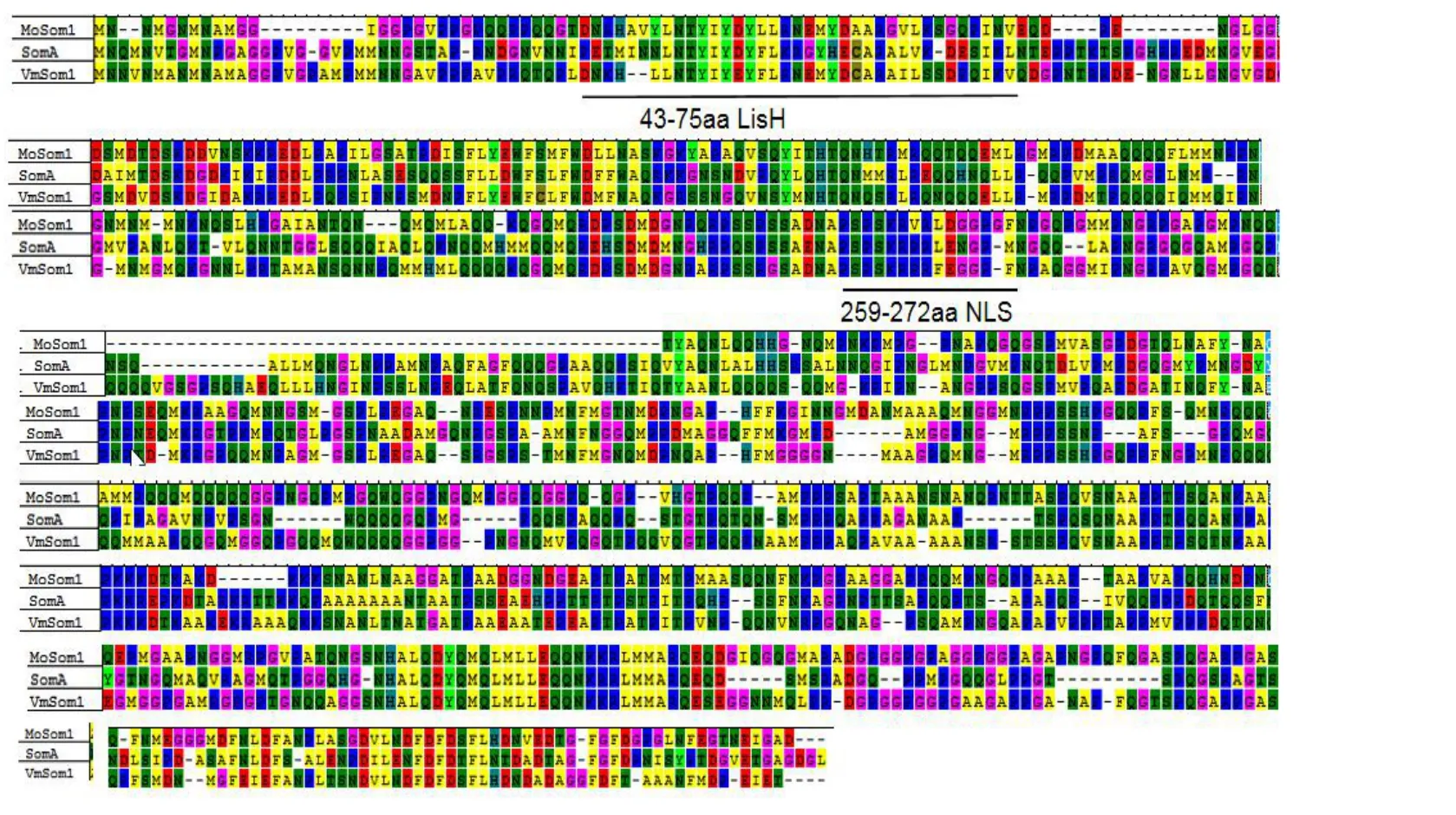
图6 三种病原菌Som1 基因编码蛋白的功能域分析Fig.6 Functional domain analysis of proteins encoded by three pathogenic Som1 genes
2.3.3 突变基因编码的蛋白质结构和理化性质分析 利用Expasy-Protparam 在线工具对蛋白的基本理化性质进行预测后发现,苹果腐烂病菌VmSom1基因的蛋白分子式为C3707H5808N1150O1211S60,相对分子质量(Mr)为87785.47,蛋白质等电点pI 为6.39,理论蛋白分子量约87.82 kDa,不稳定指数为61.07,脂肪指数为39.59,亲水性平均指数为-0.959,亲水性较强(图7)。蛋白中存在着20 种氨基酸,其中脯氨酸(13.2%)、谷氨酰胺(13%)、丙氨酸(10%)和天门冬酰胺(9.5%)含量较高,半胱氨酸(0.2%)、色氨酸(0.4%)和酪氨酸(1.1%)含量较低。使用在线Expasy-Protscale 程序分析VmSom1 蛋白质的疏水性后发现,在第134 位氨基酸处有最大疏水值为1.422,在第85 位氨基酸处有最小疏水值为-3.067。VmSom1 蛋白的二级结构预测发现,蛋白二级结构中含有α螺旋211 AA(25.61%),延伸结构37 AA(4.49%),β转角27 AA(3.28%)和无规则卷曲549 AA(66.63%)。
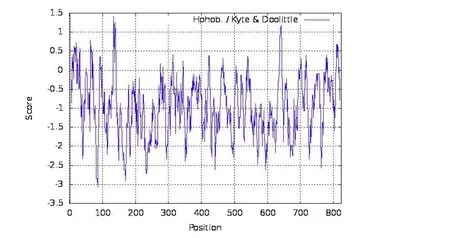
图7 突变基因编码的蛋白质疏水性图Fig.7 Protein hydrophobicity map encoded by mutant gene
3 结论与讨论
3.1 结论
本实验筛选到了V23 菌株,该菌株生长速率明显减慢,菌落表面较为湿润,菌丝生长弱,致病性完全丧失,不产生子实体。确定了T-DNA 插入突变基因位于苹果树腐烂病菌第12 号染色体69537和69547 之间。该基因的氨基酸序列与Som1基因编码的camp-dependent protein kinase pathway protein聚为一类,命名为VmSom1。基因大小为2723 bp,含有5 个外显子和4 个内含子,为单拷贝,编码区长2475 bp,编码824 个氨基酸,包含LisH 结构域和NLS 结构域,蛋白分子式为C3707H5808N1150O1211S60,理论蛋白分子量约87.82 kDa,等电点pI 为6.39,亲水性较强。
3.2 讨论
Som1 是作用于环化腺苷酸信号途径(cAMP-PKA)下游的转录因子。在丝状真菌中仅构巢曲霉(A.nidulans,Som1)、烟曲霉(A.fumigatus,SomA)、稻瘟病菌(M.oryzae,MoSom1)、大丽轮枝菌(Verticillium dahliae)和昆虫病原真菌绿僵菌(Metarhizium acridum,MaSom1)中有报道[17-21,25]。
本研究通过苹果树腐烂病菌ATMT 突变体表型筛选,获得了一突变株V23,并发现了突变基因VmSom1。苹果树腐烂病菌突变株V23 与稻瘟病菌Som1 缺失突变株(ΔMoSom1)和烟曲霉缺失SomA突变株(ΔSomA)均表现出生长减慢、致病性丧失和不产孢等相似表型[17,18,22],分析发现它们含有相同的LisH 和NLS 结构域,但是系统进化分析显示不同属中Som1基因序列差异较大,说明VmSom1可能与MoSom1 和SomA 存在相似功能,但也可能存在不同功能,需要深入研究。
本研究从转化子生长速率、菌落特征、子实体产生情况及致病性指标进行了突变体筛选,下一步需要找寻更多指标来评价突变体,继而挖掘更多的突变基因。
本研究是在Liu YG[23]和Wang Z[26]方法基础上进行,引入了接头简并引物,改良了hiTAIL-PCR法,有效提高了克隆效率且有效地降低了短链产物产生。该技术也适合于其它真菌染色体步移或侧翼序列克隆,具有高通量的特点,值得推广使用。
猜你喜欢
环球时报(2022-09-20)2022-09-20
亚热带农业研究(2022年1期)2022-08-08
蔬菜(2021年7期)2021-11-27
今日农业(2020年24期)2020-12-15
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
小猕猴智力画刊(2017年12期)2017-12-27
百科知识(2017年2期)2017-03-17
儿童故事画报·智力大王(2016年5期)2016-07-04
第二课堂(课外活动版)(2015年5期)2015-10-21
