
清水江斑鳜COI基因序列分析
2020-10-29 11:09潘秋芝
贵州畜牧兽医 2020年5期
余 科, 安 苗, 黄 胜, 潘秋芝
(贵州大学动物科学学院,贵州 贵阳 550025)
斑鳜(Sinipercascherzeri)又称桂鱼、鳌花鱼,隶属于硬骨鱼纲,鲈形目,鮨科,鳜属,鳜鱼种[1]。斑鳜刺少肉多,肉质细嫩,味道鲜美,是鱼中佳品。清水江是贵州省第二大江,是长江上游的重要支流,干流全长400 km,途经5个主要市(县),有纵横交错的支流汇入,鱼类种群丰富,其中斑鳜也有广泛分布。动物COI基因拥有蛋白编码基因所共有的特征,即密码子第3位碱基不受自然选择压力的影响,可以自由变异[2]。目前的研究支持利用线粒体基因的生物地理学分析,给种内变异确定1个普遍的标准[3]。如果检测到高的变异时,这些个体很可能是处在不同的地理隔离种群,反映出在隔离种群形成前基因库的片段。在脊椎动物中,近缘种间COI基因的差异通常要比其他线粒体大[4~8]。为了解清水江斑鳜的遗传多样性,本试验在清水江上、下游2个站点即上游(凯里江段)和下游(天柱江段)采集斑鳜样品进行线粒体COI基因序列分析,旨在为其种质资源的开发和利用提供参考数据。
1 材料与方法
1.1 样品来源采集清水江流域的斑鳜种群,其中贵州凯里江段26尾,天柱江段25尾,共计 51尾。活鱼解剖取背部肌肉3~5 g于无水乙醇中浸泡,置于 -20 ℃保存备用。标本鱼编号后浸泡于5%甲醛溶液中,保存在贵州大学水产系标本室备查。
1.2 主要仪器PCR扩增仪(AB公司,型号PTC-200 PCR)、台式高速冷冻离心机(太仓市医疗器械厂,型号TGLL-18G)、移液枪(1 000、 200、 50 μL 等量程,瑞士,型号ACURA 825)、生物安全柜(上海力申科技有限公司,型号HFsafe 900/C)、电泳仪(北京市六一仪器厂,型号DYY-2C)、凝胶成像系统(上海山富科学仪器有限公司,型号BioSens SC 710)、紫外分光光度计(型号Thermo Nano Drop 2 000 C)。
1.3 主要试剂血液/细胞/组织基因组DNA提取试剂盒、DL 2 000 DNA Marker、Loading Buffer、DNA Green、2×TaqPCR MasterMix(北京天根生化科技有限公司),三羟基甲基氨基甲烷(Tris)、乙二铵四乙酸钠盐(TEDTA·2Na)(天津化学试剂厂),4S Green Plus无毒核酸染料(上海生物工程技术有限公司)。
1.4DNA提取利用DNA提取试剂盒提取样品肌肉基因组DNA,通过1.0%琼脂糖凝胶电泳检验其完整性,再利用紫外分光光度计测定吸光值,最后置于 -20 ℃保存备用。DNA提取步骤:取肌肉样品25 mg用无菌吸水纸吸干并剪碎至肉末状,然后加入缓冲液GA 200 μL,振荡直至彻底悬浮。在得到的上层清液悬浮液中加入Proteinase K溶液20 μL混匀并置于56 ℃水浴,接着在离心管中加缓冲液GB 200 μL,充分振荡混合均匀,然后在70 ℃水浴中放置10 min后加入无水乙醇200 μL。将所得离心管内溶液和絮状沉淀用移液枪加入到1个吸附柱CB3中,接着加入缓冲液GD 500 μL并离心30 s,再加入漂洗液PW 600 μL并离心30 s,最后加入洗脱缓冲液TE 50~200 μL并再次以12 000 r/min条件下离心2 min,将溶液收集到离心管中即为DNA提取液。
1.5 引物设计、PCR扩增及测序参照NCBI中斑鳜(S.scherzeri)COI基因序列(登录号:JQ010988.1),运用primer 5软件设计引物。引物序列为F1:5’-ATGTAAAACACCTCGGGG-3’;R1:5’-TATGGGGGT TCAACTCCT-3’。送至上海生工生物工程股份有限公司合成。PCR反应体系30 μL:TaqPCR MasterMix 15 μL,上、下引物各1.5 μL,DNA样品 3 μL,用ddH2O补齐至30 μL。反应程序:94 ℃预变性3 min,94 ℃变性40 s,61.2 ℃退火1 min,72 ℃延伸50 s,72 ℃终延伸7 min(重复35个循环)。通过1.0% 琼脂糖凝胶电泳检验,最后加入Marker 2 000 作为参照,将制胶板放入电泳仪槽中,通电并设置好时间(30 min)和电压(110 V),在凝胶成像系统下观察,选择条带清晰即特异性强的样品送至上海生物工程有限公司进行双向测序。
1.6 数据分析原始序列经SepMan软件(DNA Star软件包)进行拼接、校对,去掉两端不稳定序列后,形成的完整序列在NCBI进行BLAST同源性比对。经软件MEGA6.0比对,统计其碱基组成、单倍型间的平均遗传距离、多态位点。基于K2双参数模型构建邻接(Neighbor joining,NJ)系统进化树。利用软件DNASP分析单倍型多样度(Hd)、核苷酸多样性(Pi)及平均核苷酸差异数(K)。
2 结果与分析
2.1PCR扩增利用扩增引物(F1R1)对51尾斑鳜样品进行PCR扩增,发现所有样品均扩增出约1 500 bp条带(见图1)。经去除两端不稳定序列,最终得到序列长度为962 bp,共编码320个氨基酸。
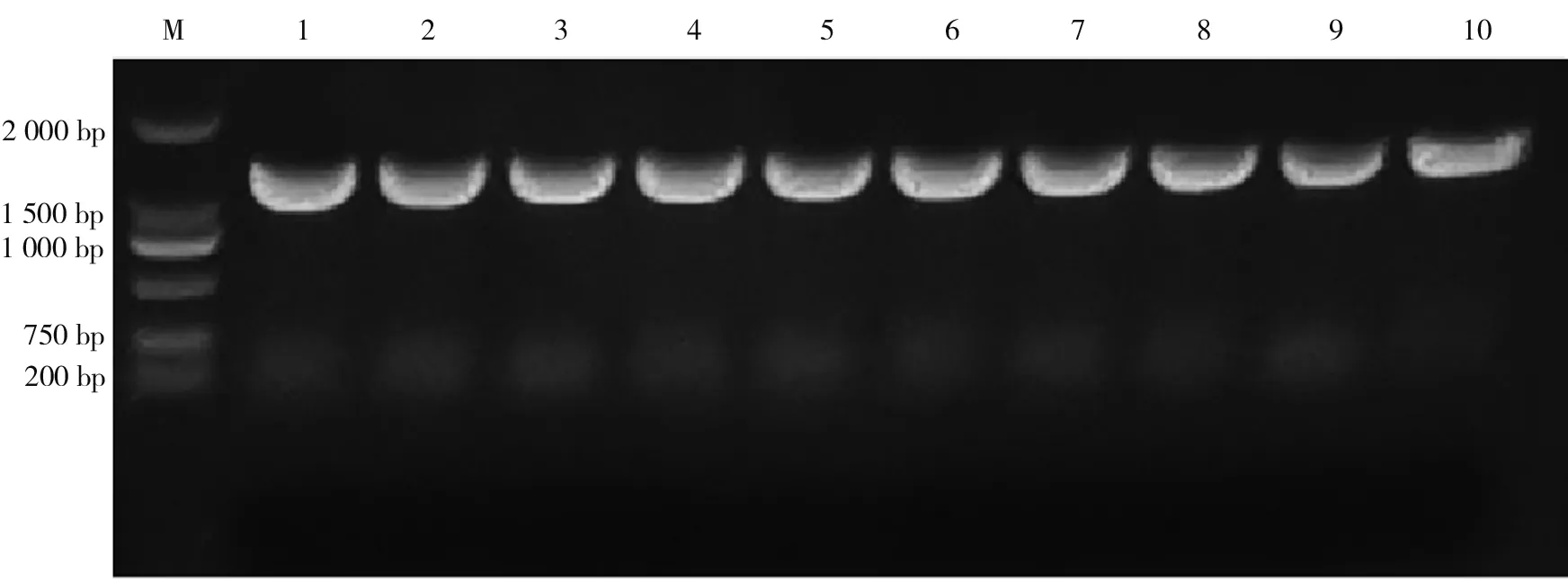
M:DL 2 000 DNA Marker; 1~10:部分PCR扩增产物图1 电泳检测产物
2.2 碱基组成及变异位点经上海生物工程有限公司双向测序得到51个样品共界定9种单倍型。由表1可见:碱基T(U)的含量为25.6%~25.9%(平均值为25.8%),C的含量为17.7%~18.0%(平均值为17.8%),A的含量为27.5%~27.6%(平均值为27.5%),G的含量为28.8%~28.9%(平均值为28.9%)。其中A+T的平均含量为53.3%,大于C+G的平均含量(46.7%)。在基因序列中A、T(U)、G的含量较高,C的含量相对较低。在962 bp长度的基因序列中有9个多态位点,未检测到插入或缺失位点。其中转换、颠换、转换与颠换共存位点数分别为7、2、0个;转换与颠换比为1∶13.881。多态位点数占总位点数的0.94%,其中转换、颠换的位点数分别占总位点数的0.72%、0.208%。

表1 单倍型碱基组成
2.3 核苷酸变异位点由表2可见每种单倍型的变异位点。在位点256(nt)Hap8中由碱基G转换成A;在位点331(nt)Hap6、Hap8中由碱基C转换成T;在位点344(nt)Hap9中由碱基T转换成C;在位点379(nt)Hap5中由碱基C转换为T;在位点401(nt)Hap4中由碱基T转换为C;在位点403(nt)Hap4中由碱基C颠换为A;在位点419(nt)Hap7中由碱基G颠换成T;在位点446(nt)Hap2、Hap7、Hap9中由碱基T转换为C;在位点449(nt)Hap3中由碱基C转换成T。没有转换与颠换共存位点。主要是以T与C之间转换的变异位点为主。

表2 核苷酸变异位点
2.4 单倍型组成通过软件DNASP5分析,51个清水江斑鳜样品中含有9个单倍型。由表3可见:单倍型Hap1有6尾斑鳜是同一单倍型,其中凯里江段和天柱江段各有3尾;单倍型Hap2有29尾斑鳜是同一单倍型,其中凯里江段和天柱江段分别有15尾和14尾;单倍型Hap5有5尾斑鳜是同一单倍型,其中凯里江段和天柱江段分别有4尾和1尾;单倍型Hap6有6条斑鳜是同一单倍型,其中凯里江段和天柱江段分别有4尾和2尾。其余的(Hap3、Hap4、Hap7、Hap8、Hap9)单倍型各有1尾斑鳜为同一单倍型,且都是天柱江段。

表3 单倍型组成 个
2.5 遗传多样性在51个样品中,凯里江段的26个斑鳜样品核苷酸多样性为0.001 09,平均核苷酸差异数为1.049 23,单倍型多样度为0.630 77;天柱江段的25个斑鳜样品核苷酸多样性为0.001 31,平均核苷酸差异数为1.260 00,单倍型多样度为0.683 33。整个清水江斑鳜种群中核苷酸多样性为 0.001 19,平均核苷酸差异数为1.143 53,单倍型多样度为0.650 20。
2.6NJ系统进化树及其遗传距离参照NCBI中的相似种群序列构建NJ系统进化树(1 000次抽样,bootstrap检验),其中有鄱阳湖斑鳜[S.scherzeri(JQ010985)Poyang Lake]、湘江斑鳜[S.scherzeri(JQ010988)Xiangjiang River]、湖南斑鳜[S.scherzeri(EF143391)Hunan Changde]、洞庭湖斑鳜[S.scherzeri(NC015815)Dongting Lake]、朝鲜斑鳜[S.scherzeri(EF143392)Korea Changju]等,另外选择同科的少鳞鳜[C.whiteheadi(JK149811)]等作为外群基因序列进行建树。由图2可见:清水江斑鳜与另外8支参照斑鳜归为1支,与斑鳜同科的少鳞鳜分为另1支,说明与少鳞鳜的亲缘关系较远。从进化树中可以看出,长江流域的斑鳜聚为1支,置信度很高,说明长江流域的斑鳜种群间也存在基因交流的情况且亲缘关系较近;清水江斑鳜与鄱阳湖斑鳜互为姐妹支,亲缘关系很接近。

图2 NJ系统进化树
2.7 单倍型之间的遗传距离由表4可见:9种单倍型的最大遗传距离为0.004 2,最小遗传距离趋近于0,平均遗传距离为0.002 6。其中凯里江段平均遗传距离为0.001 1±0.000 6,天柱江段平均遗传距离为0.001 3±0.000 6。这2个江段种群间的遗传距离为0.001 2±0.000 6。
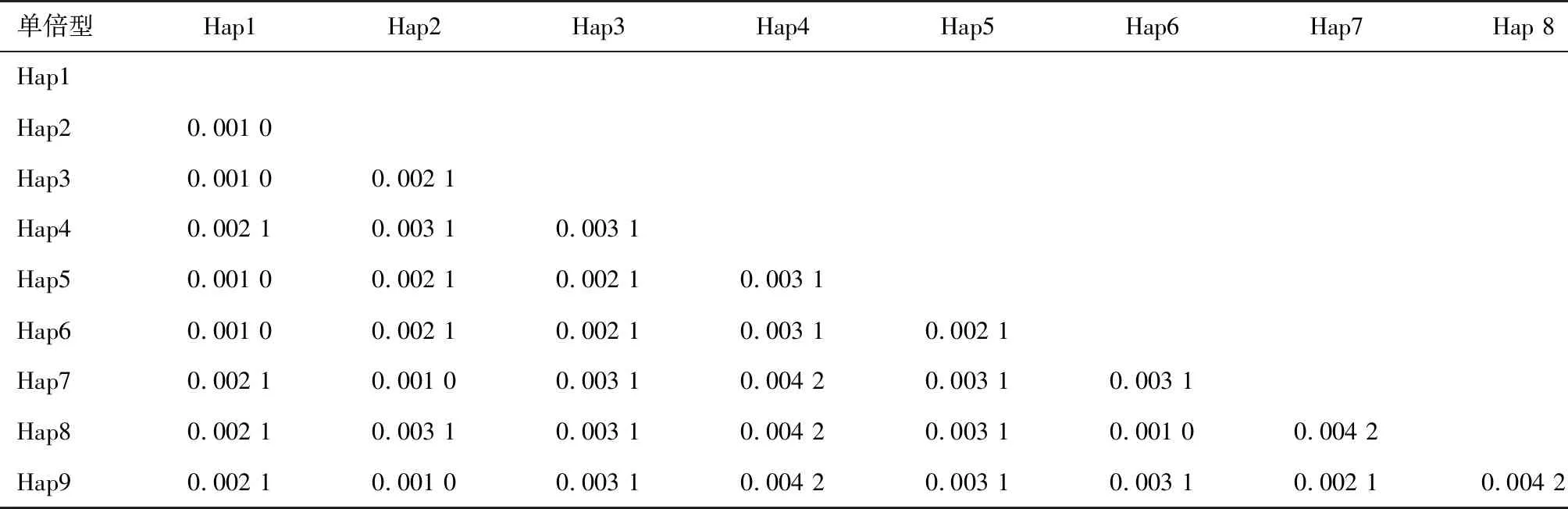
表4 单倍型之间的遗传距离
3 讨论
3.1 碱基组成及多样性本研究中,清水江斑鳜核苷酸条形码区段不存在碱基的插入、缺失,T(U)、C、A、G的碱基平均含量分别为 25.8%、17.8%、27.5%、28.9%,其中A+T的平均含量(53.3%)大于C+G的平均含量(46.7%)。研究结果与肖同乾等[9]对8种鲤鱼养殖品种的研究结果相似(T、 C、 A、G碱基的平均含量分别为 26.8%、29.0%、29.7%、14.5%,A+T的含量为56.5%,明显高于G+C的含量 43.5%)。本研究中斑鳜群体在碱基上存在明显差异,表现出明显的偏倚性。张凤英等[10]对3种鲳属鱼类线粒体COI基因序列的研究中也存在类似现象。其次转换发生的频率远远大于颠换,这与Desalle R T等[11]提出的结论一致,造成这类情况的原因是由于DNA在进化过程中,分歧较近的物种间发生同义突变所造成的,转换与颠换的比值与时间成反比。Grant W S等[12]将遗传多样性分为4种类型。第1类:单倍型多样度高(Hd>0.5)和核苷酸多样性高(Pi>0.005);第2类:单倍型多样度高(Hd>0.5)和核苷酸多样性低(Pi<0.005);第3类:单倍型多样度低(Hd<0.5)和核苷酸多样性高(Pi>0.005);第4类:单倍型多样度低(Hd<0.5)和核苷酸多样性低(Pi<0.005)。结合本文试验数据,凯里江段和天柱江段斑鳜的单倍型多样度都高,核苷酸多样性都低,属于第2类,但凯里江段的核苷酸多样性和单倍型多样度均低于天柱江段的核苷酸多样性和单倍型多样度,说明在清水江斑鳜群体中天柱江段的遗传多样性水平比凯里江段高,遗传资源也比凯里江段丰富[13,14]。在清水江斑鳜群体中核苷酸多样性为0.001 19,均低于清水江鲤鱼群体(0.004 51)[3]和清水江白甲鱼种群(0.010 7)[15]。再由单倍型多样度与浙江省3个斑鳜野生群体细胞色素b基因序列进行比较,清水江斑鳜的单倍型多样度为0.650 2,浙江省野生斑鳜群体的单倍型多样度为0.572~0.688[16]。很显然,清水江斑鳜群体的单倍型多样度介于斑鳜野生群体之间且数值都偏高。综上所述,在清水江的鱼类群体中,斑鳜的遗传多样性没有鲤鱼和白甲鱼丰富,其遗传资源相对来说也较为匮乏,但是相对于浙江省野生斑鳜来说,清水江斑鳜遗传资源更为丰富。
3.2 遗传距离与系统进化树遗传距离指不同种群之间存在着基因差异的现象,这种基因之间的差异可用特定数值来衡量,数值越大,遗传距离越远,反之遗传距离越近。在遗传距离的分析中,天柱江段的遗传距离大于凯里江段,说明天柱江段斑鳜种群的遗传变异程度比凯里江段高。系统进化树是被认为具有共同祖先的物种经过不断演化,最后呈现出各个物种之间亲缘关系的树形图。在图2中能直观地看到各物种之间的亲缘关系以及遗传距离的远近,每个节点代表其各分支的最近共同祖先,其数值代表置信度,置信度越高,说明2者由同一祖先演化而来的概率越大。当然也可能是同一祖先演化而来,但是在演化过程中发生变异或者因为地理隔绝造成了此类情况[17]。而节点间的线段长度对应演化距离,线段越短,演化距离短。清水江斑鳜的51个样品9个单倍型全聚为1支,置信度达100,表明清水江的2个斑鳜地理群体之间的亲缘关系极其相近;除此之外鄱阳湖种群与之聚类在一起,亲缘关系接近,且与其余几个斑鳜种群间互为姊妹支,说明生物在生长进化历程中可能是遗传的不定向性及生存环境之间的不同,从而造成了这种差异。
3.3 清水江斑鳜资源的保护通过对清水江斑鳜COI基因序列的分析,以及系统进化和多样性分析,进而为保护其种质资源和生态环境多样性提供了1份数据参考。遗传多样性可反映物种进化潜力,多样性丰富程度越高,适应环境的能力就越强[18]。经实验得出数据,在清水江所采集的2个斑鳜群体中,天柱江段与凯里江段遗传多样性存在一定的差异[19]。通过分析,我们应对凯里江段的水资源及其鱼类种质资源进行保护,尽量减少地理隔绝所造成物种间的基因交流受限和杜绝电捕等破坏生态平衡的事件。维护生态平衡,不破坏生物食物链各上下级之间的关系。不管是鱼类还是其他生物,都要尽量避免朝着不利于其自身发展的方向演变。在今后的研究中,应多对清水江不同河段的斑鳜种群间的遗传差异进行分析,为保护斑鳜的种质资源,改良斑鳜的优良品种提供更加充分的依据[20]。
4 结论
由评价遗传多样性的2项指标可见,清水江斑鳜的遗传多样性较为丰富,且2个群体的遗传谱系更接近于洞庭湖和鄱阳湖的斑鳜群体;其中天柱江段斑鳜的遗传多样性比凯里江段斑鳜高,遗传资源更为丰富。
猜你喜欢
贺州学院学报(2021年1期)2021-06-08
中国水利(2020年23期)2020-12-24
科学导报(2020年27期)2020-05-13
艺术品鉴(2020年1期)2020-01-19
艺术品鉴(2019年7期)2019-09-02
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
晚晴(2016年6期)2016-05-14
学习月刊(2015年23期)2015-07-09
原生态民族文化学刊(2014年3期)2015-02-03
中山大学学报(社会科学版)(2014年2期)2014-03-01
