
Role of higher levels of post-challenge antibodies in protective vaccination against Leishmania tropica infection of BALB/c mice
2020-10-28 02:43MosayebRostamianHamidNiknam
Mosayeb Rostamian,Hamid M.Niknam
1Infectious Diseases Research Center,Health Institute,Kermanshah University of Medical Sciences,Kermanshah,Iran
2Immunology Department,Pasteur Institute of Iran,69 Pasteur Avenue,Tehran 13169-43551,Iran
ABSTRACT
KEYWORDS: Leishmania tropica; Vaccination; Antibody; IgG1;IgG2a
1.Introduction
Leishmaniasis is a disease caused by different species of Leishmania parasites.The main forms of leishmaniasis are visceral,mucocutaneous,and cutaneous[1].Cutaneous leishmaniasis (CL) in the Old World is mainly caused by Leishmania major (L.major) and Leishmania tropica(L.tropica).Unlike L.major,studies on L.tropica pathology and host immunity are rare[2].
There are differences between the immune responses to different Leishmania species[3-5].It is well known that,in BALB/c mice,there are differences between L.major and L.tropica with regard to the disease outcome and the intensity of immune response.L.major causes a systemic and progressive fatal disease which evokes high delayed-type hypersensitivity and immune responses,while L.tropica causes mild swelling or small lesions with low delayedtype hypersensitivity and immune responses[6-11].Additionally,we evaluated the role of immunoglobulin G (IgG) isotypes (IgG1 and IgG2a) in L.tropica infection in BALB/c mice and showed that L.tropica causes a lower IgG1 response than L.major[12].Furthermore,we found that there is heterogeneity among immune responses to L.tropica in BALB/c mice models[13].
Antibody responses may have detrimental or beneficial roles in protecting the host against Leishmania infections.It has been shown that IgG could exacerbate Leishmania mexicana infections in mice[14].A previous report had shown that IgG not only fails to provide protection against L.major,but also it contributes to disease progression[15].As a beneficial role of antibodies,it has been shown that IgG production is required for the development of Th1-dependent immunity and control of CL in mice[16].Further studies showed that beneficial effect of antibodies in the disease control is more associated with IgG2a subtype of antibodies[17-23].To tailor humoral responses to the benefit of the host,it is essential to know which isotype(s) of the antibody is protective (or pathogenic)[24].
In the present study,anti-Leishmania antibody responses were studied after vaccination and after challenge infection.These vaccines included L.major soluble Leishmania antigen (SLA),L.tropica SLA,and recombinant L.tropica stress-inducible protein-1(rLtSTI1) of L.tropica.Our previous studies have shown that these antigens can protect BALB/c mice against Leishmania challenge if they are accompanied with a suitable adjuvant[25,26].Due to the importance of humoral immune response and lack of enough information in this field,it is necessary to carry out further study regarding the role of antibodies in protection against L.tropica infection.In the present study,antibody responses against L.tropica infection in BALB/c mice models were evaluated.
2.Materials and methods
2.1.Parasite,antigens,and adjuvant
Two Leishmania species were used in this study,L.major strain MRHO/IR/75/ER and L.tropica strain MHOM/AF/88/KK27.L.major was kindly donated by M.Mohebali (School of Public Health,Tehran University of Medical Sciences,Tehran,Iran) and L.tropica was a gift from Dr.D.Sacks (Laboratory of Parasitic Diseases,National Institute of Health,Bethesda,Maryland,USA).Species confirmation,culture,and maintaining,as well as testing of virulence preservation of the parasites were the same as our previous reports[27,28].Briefly,species were confirmed by PCR using internal transcribed spacer (ITS1) specific primers and cultured in Novy-MacNeal-Nicolle (NNN) medium at 24-26 ℃.The virulence of parasites was maintained by injection into BALB/c mice and retrieval from their lymph node.After isolation from mice,the cultivation was kept to less than three passages in NNN medium to maintain the virulence.
For preparation of SLAs,cultivated parasites were harvested at the stationary phase and after three-times of washing with PBS buffer,they were lysed through five rounds of freeze-thaw (F/T)cycles.F/T suspension was centrifuged (16 000×g,4 ℃,20 min)and the supernatant was collected as SLA.rLtSTI1 was cloned and expressed using Escherichia coli expression system as previously reported[26].Briefly,L.tropica genomic DNA was extracted and the LtSti1 region was amplified by Exprime Taq enzyme (GeNet Bio,South Korea) using specific primers harboring NdeⅠand BamHⅠrestriction sites.The amplified sequence was digested by restriction enzymes and cloned in pET-15b expression vector.The vector LtSti1-pET-15b construct was transferred to Escherichia coli BL21(DE3) and the protein was expressed under certain conditions of isopropyl β-D-thiogalactoside concentration.The expressed protein was verified by Western blotting using anti-HisG-HRP Antibody(Invitrogen,Carlsbad,USA),according to the manufacturer’s instructions.Monophosphoryl lipid A (MPLA) was purchased from Invivogen (France).
2.2.Mice and blood collection
A total of 150 female,5- to 7-week-old BALB/c mice were purchased from Pasteur Institute of Iran (Tehran,Iran).Fifteen mice were used for each study group.Retro-orbital bleeding procedure was applied for collecting animal blood samples.After blood collection,it was incubated at room temperature for 15-30 min for clot formation.Then,it was centrifuged at 2 000×g for 10 min and the supernatant was harvested as serum.
2.3.Immunization and infection
Immunization and infection were done as previously reported[25,26].Briefly,mice were divided into three main groups (45 mice for each main group): 1) L.major SLA,2) L.tropica SLA,and 3) L.tropica rLtSTI1.Each main group was divided into three groups (two test groups and one control group,15 mice per group).The first main group was vaccinated by L.major SLA with/without adjuvant and challenged by L.major.The second main group was vaccinated by L.tropica SLA with/without adjuvant,and the third main group was vaccinated by rLtSTI1 with/without adjuvant.Both of the second and third main groups were challenged by L.tropica.For each main group,a group was vaccinated by phosphate buffered saline(PBS) and used as the control.Mice were vaccinated three times at intervals of two weeks into the hind footpad subcutaneously.MPLA was used as adjuvant and prepared according to the manufacturer’s instruction.Twenty µg of MPLA was administered in each vaccine preparation.Six weeks after the last vaccination,mice were challenged subcutaneously with 1 × 106L.major or 2 × 106L.tropica stationary phase promastigotes based on assigned groups.The whole experiment was performed twice.
In order to evaluate long-term antibody responses,15 additional mice were included in L.tropica rLtSTI1 main group (five mice per each group) and were vaccinated similar to the rest of the group,but were challenged 16 weeks after the last vaccination.The parasite load and antibody levels were measured six weeks after the challenge.
2.4.Evaluation of protection against Leishmania infection
Protection against challenge in L.major-infected groups was evaluated up to week 8 after challenge and in L.tropica-infected groups at two intervals of week 8 and week 16 after challenge.For L.major-infected groups,the measurement was discontinued after week 8 due to severe footpad necrosis in control groups.Reductions in lesion diameter and in parasite load were considered as protection against L.major.Reduction in parasite load in the lymph node was considered as protection against L.tropica because the challenge by this species did not result in significant lesion development.The parasite load was determined by limiting dilution assay in L.major infected mice and by real-time PCR in L.tropica infected mice as previously reported[25,26].
2.5.Antibody assay
Antibody in each experimental group was assayed using the same priming antigen e.g.the antigens of L.tropica SLA and rLtSTI1 were used for assay of antibody in main groups of L.tropica SLA and L.tropica rLtSTI1,respectively.To perform ELISA,three factors were optimized: serum dilutions (in the range of 1/500,1/1 000,1/2 000,1/4 000,1/8 000,1/16 000,and 1/32 000),dilution of anti-mouse IgG1 and IgG2a (in the range of 1/1 000 and 1/2 000),and antigen concentration (in the range of 0.2,0.5,1,and 2 µg/well).
The respective antigens (0.2 µg/well of L.major SLA,L.tropica SLA,or rLtSTI1 depending on the studied main group) were coated in 96-well ELISA plates (Greiner,Microlon) and incubated overnight at 4 ℃.After washing by 250 µL/well of PBST (PBS +0.05% v/v Tween 20) three times,the plates were blocked by 1% w/v bovine serum albumin in PBS (100 µL/well) and incubated at 37 ℃ for 2 h.The plates were washed again as above,optimized dilution of sera was added (100 µL/well) to plates in triplicate,and the plates were incubated at 37 ℃ for 2 h.After washing steps,100 µL of 1/2 000 diluted anti-mouse IgG1 or IgG2a antibodies(BD PharmingenTM) was added to the previously assigned wells and plates were incubated for 2 h at 37 ℃.The plates were washed again as mentioned,tetramethylbenzidine was added (100 µL/well),and the plates were incubated for 15 min at room temperature in a dark place.By adding 100 µL/well of stop solution (2N H2SO4),the enzyme-substrate reaction was stopped and absorbance of each well was read at both 620 and 450 nm by a microplate reader(Anthos 2020; Eugendorf,Austria).Final values were calculated by subtracting 620 nm absorbance values from 450 nm values.
2.6.Statistical analysis
Optical density (OD) of ELISA results of different groups were compared by one-tailed two-sample equal variance t-test.Data were expressed by mean ± SD or SEM.P values < 0.05 were considered significant.
2.7.Ethical statement
Experiments on animals were done following the National Institutes of Health Guide for the Use of Laboratory Animals (publication# 80-23) and the guidelines for animal care prepared by Pasteur Institute of Iran.The study was approved by the Ethics Committee of the Pasteur Institute of Iran (license number 95/0201/20704).
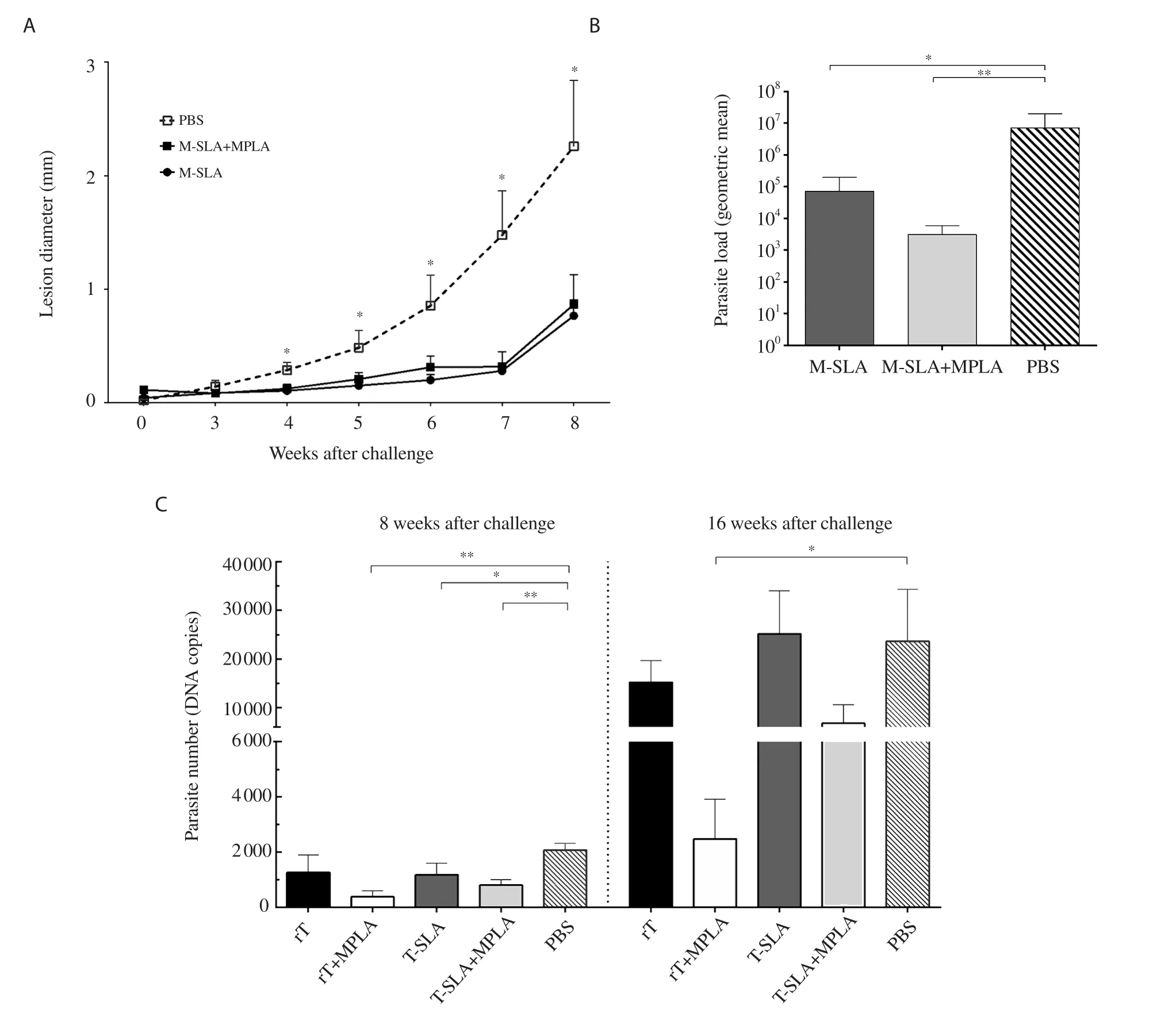
Figure 1.Effect of vaccination by three different antigens and adjuvant on protection against Leishmania challenge.Six weeks after the last vaccination,mice were challenged by stationary promastigotes of Leishmania major (L.major) or Leishmania tropica (L.tropica).(A) The thickness of L.major-infected mice footpad was determined weekly.Each point indicates values of 15 mice per group (mean ± SEM).*Significant difference in comparison to the PBS control group (P<0.05).(B) The draining lymph nodes of L.major-infected mice were removed at week 8 after challenge and were assayed for parasite load.*Significant difference compared with the PBS control group (*P<0.05; **P<0.01).Each bar shows mean ± SEM of parasite load of five mice per group.(C)The draining lymph nodes of L.tropica-infected mice were removed at week 8 and week 16 after the challenge and were assayed for parasite load.The parasite loads of five mice per group are shown as bars (mean ± SEM).*Significant difference in comparison to the PBS control group (*P<0.05; **P<0.01).The data is adapted from our previous publications with permission[25,26].M-SLA: L.major soluble Leishmania antigen,rT: recombinant L.tropica stress-inducible protein-1,MPLA: monophosphoryl lipid A,T-SLA: L.tropica soluble Leishmania antigen,PBS: phosphate buffered saline.
3.Results
3.1.Protection induced by vaccination
Lesion diameter was significantly lower in the groups treated with SLA and SLA+MPLA in comparison with the PBS group in L.major-infected mice from week 4 to the end of the study (Figure 1A).In addition,at 8 weeks after the challenge,parasite loads were markedly lower in lymph nodes of vaccinated groups compared to the PBS group (Figure 1B).
Different levels of protection were observed in L.tropica SLA and rLtSTI1-administered groups.The rLtSTI1+MPLA group showed protection 8 weeks and 16 weeks after challenge while the rLtSTI1 group showed no significant protection in both intervals.The groups of L.tropica-SLA+MPLA and L.tropica-SLA showed significantly lower parasite loads than PBS at only 8 weeks after challenge while no protection was observed at 16 weeks post-challenge (Figure 1C).
3.2.Optimization of ELISA setting by testing different dilutions of sera
Using different amounts of coating antigens,and different dilutions of sera and secondary antibodies,the ELISA was optimized.The optimized dilution for coating antigen and secondary antibody was 0.2 µg/well and 1/2 000,respectively.For optimization of serum dilution,all sera were always tested in seven two-fold dilutions of 1/500 to 1/32 000,and the best dilution was selected for further ELISA experiments.The best dilution means the dilution in which the best discrimination of all groups can be achieved.As an example,Figure 2 shows the OD450nmof three different groups of rLtSTI1+MPLA,L.tropica-SLA+MPLA,and PBS at different sera dilutions.The best dilutions were 1/2 000 and 1/4 000 because discriminations of all groups were clearly observed in these dilutions.At higher dilutions (1/8 000,1/16 000,and 1/32 000),the OD450nmwas too low to differentiate between the groups.On the other hand,at lower dilutions (1/500,and 1/1 000),OD450nmof rLtSTI1+MPLA was too high to be detected by ELISA reader (Figure 2).
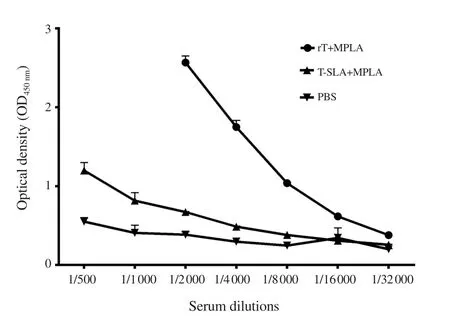
Figure 2.Results of ELISA optimization.The levels of antibody(IgG1) of seven different serum dilutions of three experimental groups(rLtSTI1+MPLA,L.tropica-SLA+MPLA,and PBS) were compared.
3.3.Higher levels of antibodies induced by vaccination of recombinant protein
Antibody response was compared against two types of antigen i.e.whole cell lysate (L.tropica-SLA) and recombinant protein(rLtSTI1).Antibody (either IgG1 or IgG2a) levels were higher in the mice vaccinated by recombinant protein with/without adjuvant compared to the mice vaccinated by L.tropica-SLA with/without adjuvant (Figure 3).
3.4.Vaccination resulted in antibody responses
All vaccinated groups (L.major-SLA,L.major-SLA+MPLA,L.tropica-SLA,L.tropica-SLA+MPLA,rLtSTI1,and rLtSTI1+MPLA)caused a higher antibody response (both IgG1 and IgG2a) in comparison to the PBS control group,although these differences were only statistically significant between some groups (Figure 4).
3.5.Pattern of post-challenge humoral response against L.tropica and L.major
In L.major SLA main group,the vaccinated groups showed lower IgG1 response than the control group after the challenge.However,there was no significant difference in IgG2a level between the vaccinated and PBS control group,8 weeks after challenge (Figure 5).The data from mice in L.major SLA main group were not included in 16 weeks-assays due to severe necrosis in the footpad.
In contrast to L.major-vaccinated groups,the levels of antibodies(both IgG1 and IgG2a) in the vaccinated group of rLtSTI1+MPLA were higher than the PBS control group at both 8 and 16 weeks after challenge.Moreover,the levels of antibodies (both IgG1 and IgG2a)were also higher in the rLtSTI1+MPLA group in comparison to the rLtSTI1 group at both 8 and 16 weeks after challenge (Figure 5 and Figure 6).
3.6.Long-term protection and humoral response in L.tropica rLtSTI1 main group
The maintenance of a protective immunity for a long time after vaccination is an important criterion for vaccines,especially for recombinant ones.The long-term immune responses were assayed only in the groups that received the recombinant protein vaccine because short-term results in our previous report had shown that this vaccine is more effective than L.tropica-SLA[26].No long-term assay was done for “L.major SLA” and “L.tropica SLA” main groups.
The rLtSTI1+MPLA group showed a lower load of parasite in comparison to the PBS group with no significant difference(Figure 7A).Like short-term assay,rLtSTI1+MPLA groups showed higher levels of antibody (both IgG1 and IgG2a) than the PBS group at long-term (16 weeks) after vaccination (Figure 7B and 7C),suggesting that the protection still remain unchanged in rLtSTI1+MPLA group until 16 weeks after vaccination.
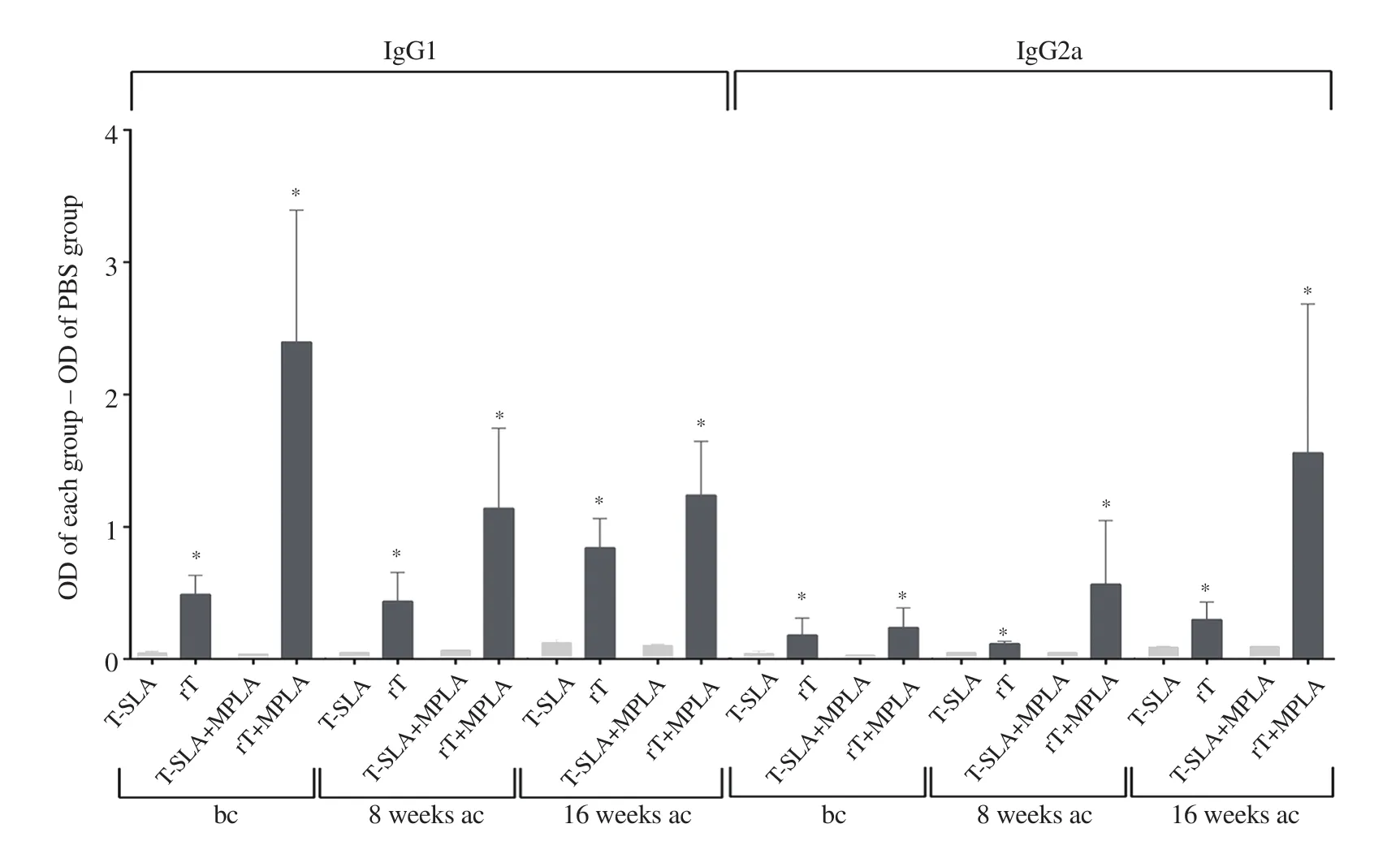
Figure 3.Levels of antibodies produced against rLtSTI1 in comparison to SLA.The OD of the PBS control group was subtracted from the OD of each antigenadministered group (L.tropica SLA and L.tropica rLtSTI1 main groups) and the resulting data were compared.*Significant difference between rLtSTI1- and L.tropica SLA-administered groups (P < 0.05).bc: before challenge,ac: after challenge.
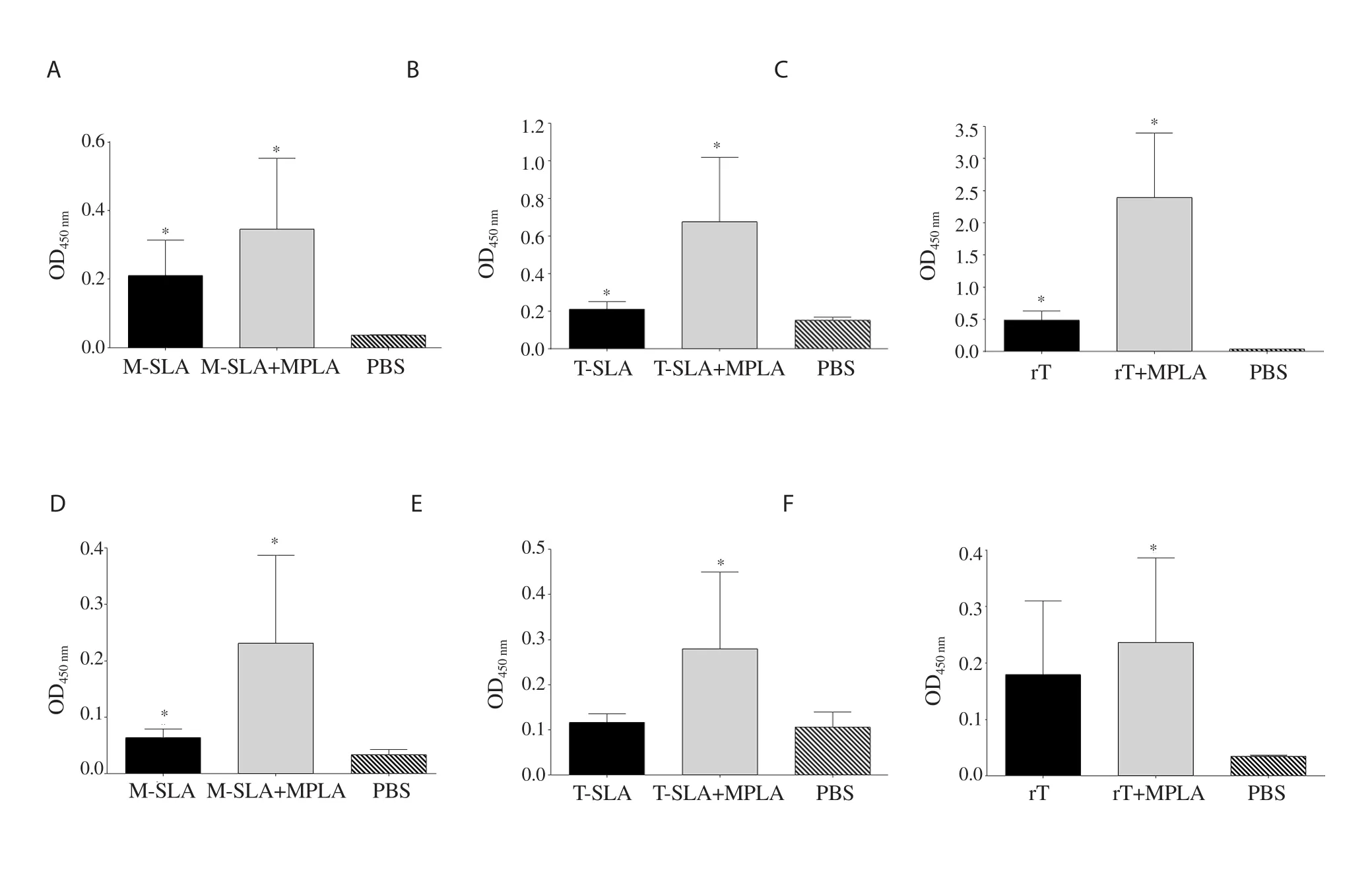
Figure 4.Humoral responses of mice after vaccination.Six weeks after vaccination,IgG1 and IgG2a response of each group was measured.The IgG1 levels of main groups of (A) L.major SLA,(B) L.tropica SLA,and (C) L.tropica rLtSTI1.(D-F) The IgG2a levels of the same main groups.Antibody response of main group L.major SLA,L.tropica SLA,and L.tropica rLtSTI1 was measured at 1/8 000,1/500,and 1/16 000 dilutions of the sera,respectively.*Significant difference between the vaccinated group and the PBS control group (P<0.05).
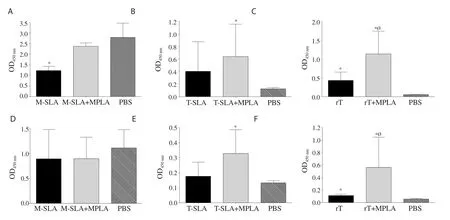
Figure 5.Humoral response of mice 8 weeks after the challenge.The IgG1 levels of main groups (A) L.major SLA,(B) L.tropica SLA,and (C) L.tropica rLtSTI1.(D-F) The IgG2a levels of the same main groups.Antibody response of main group L.major SLA,L.tropica SLA,and L.tropica rLtSTI1 was measured at 1/8 000,1/500,and 1/16 000 dilution of the sera,respectively.*Significant difference compared with the PBS control group (P<0.05).ØSignificant difference between the rLtSTI1+MPLA and rLtSTI1 groups (P<0.05).
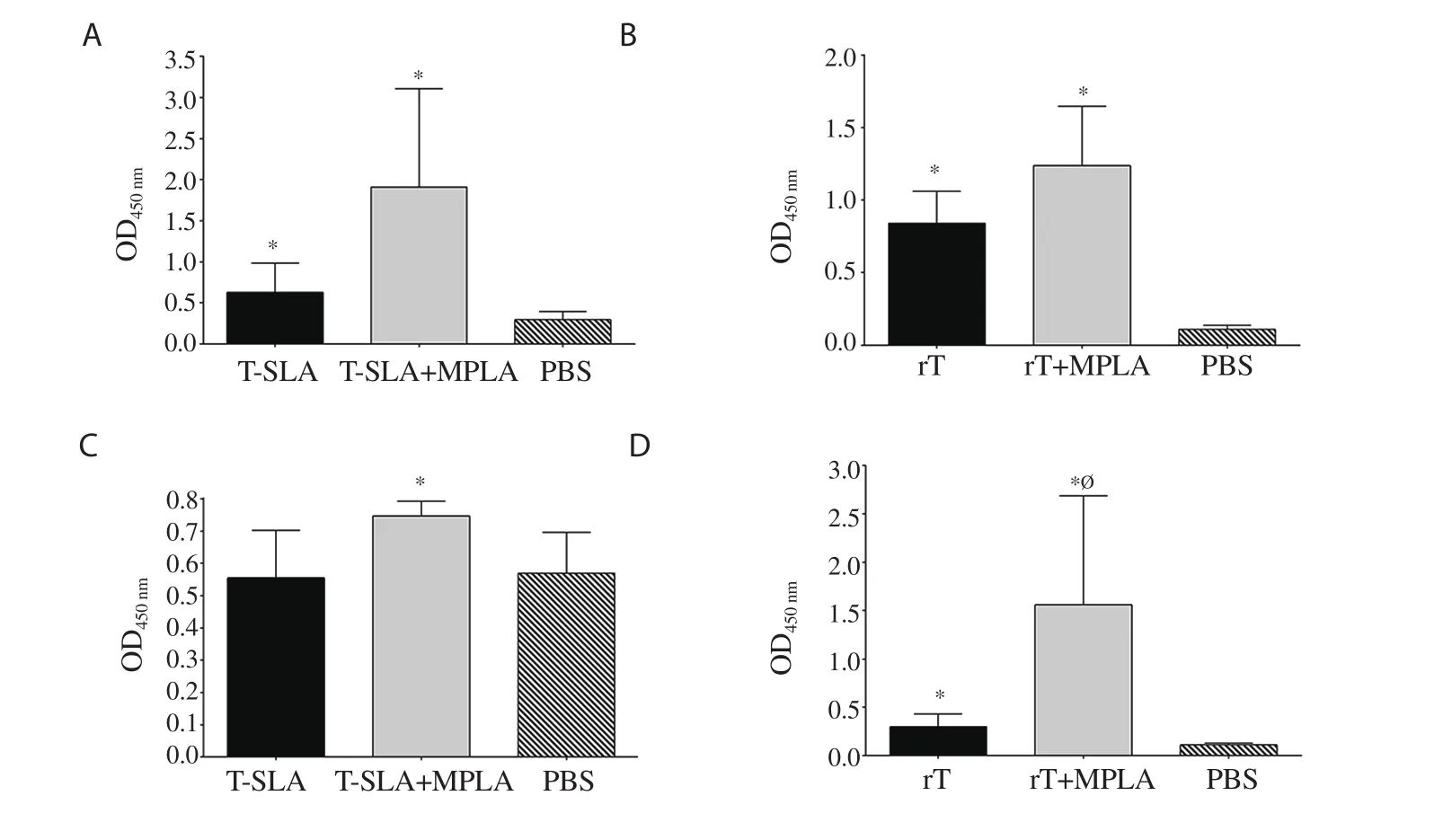
Figure 6.Humoral response of mice 16 weeks after the challenge.The IgG1 levels of main groups (A) L.tropica SLA,and (B) L.tropica rLtSTI1.(C-D) The IgG2a levels of the same main groups.Antibody response of main group L.tropica SLA,and L.tropica rLtSTI1 was measured at 1/500 and 1/4 000 dilution of the sera,respectively.*Significant difference compared with the PBS control group (P<0.05).ØSignificant difference between the rLtSTI1+MPLA and rLtSTI1 groups (P<0.05).
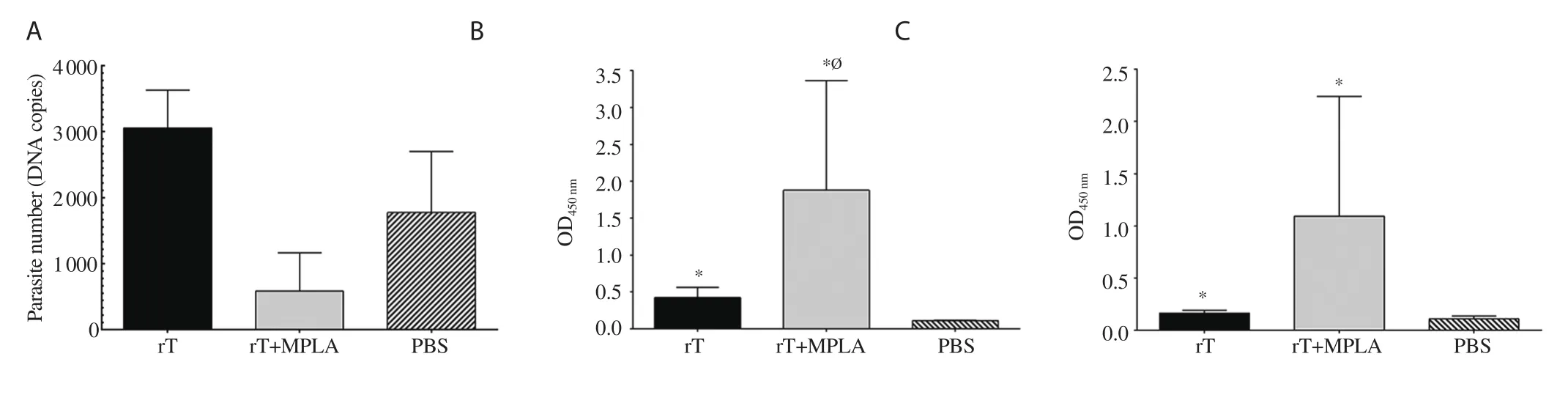
Figure 7.Protection and humoral response of L.tropica rLtSTI1 main group mice in long-term after vaccination.Five mice of L.tropica rLtSTI1 main group were challenged 16 weeks after the last vaccination by L.tropica.Mice were bled and killed six weeks after challenge and (A) parasite load,the level of (B)IgG1 and (C) IgG2a were measured at 1/4 000 dilution of the sera.The parasite loads of three mice per group were shown as bars (mean±SEM).*Significant difference compared with the PBS control group (P<0.05).ØSignificant difference between the rLtSTI1+MPLA and rLtSTI1 groups (P<0.05).
4.Discussion
Th1 and Th2 are two main types of immune responses which determine the disease outcome in leishmaniasis[29].Antibody responses are mainly switched toward IgG1 after activation of Th2 type responses mediated by anti-inflammatory cytokines such as IL-4[30,31].As reported,the fundamental immune response of BALB/c mice to L.major is Th2 type responses which result in progressive pathogenesis[29].In order to control disease in this animal model,the Th2 response should be decreased.Therefore,it seems rational that IgG1 is decreased in the vaccinated groups of our study after being challenged by L.major.Consistent to our study,previous studies on L.major have shown that protective immunity is associated with lower IgG1 response than IgG2a in resistant mice (C57BL/6)[29,32],FcγR-knockout mice[33],and protected BALB/c mice by vaccination[34,35].Similarly,mice that have been protected by vaccination against other Leishmania species show a lower IgG1 response[36-38].
We found that in BALB/c mice,a protective vaccine against L.major usually causes high antibody response after vaccination.However,after challenge,a lower level of IgG1 antibody is produced in protected mice.Concordant with our findings,previous studies have shown that vaccines that protect animals against L.major or Leishmania infantum result in high levels of antibody (both IgG1 and IgG2a) response in BALB/c mice after vaccination[39,40].How this post-vaccination high level of IgG1 leads to post-challenge protection in the mice remains unknown.
In contrast to the mice challenged by L.major,challenging BALB/c mice by L.tropica resulted in higher post-challenge antibody (both IgG1 and IgG2a) in the vaccinated groups.In addition,we observed higher post-challenge antibody response in groups showing higher protection compared to groups with lower protection,which suggests that higher antibody levels are associated with more protection in these groups.
We previously revealed that lower levels of IgG1 are produced in response to L.tropica in comparison to L.major infection of BALB/c mice[12].Moreover,previous works have reported that the immune response to L.tropica is much lower than L.major[9,41].In contrast to L.major which causes severe and progressive pathogenesis in BALB/c mice,L.tropica causes mild pathogenesis leading to a delayed and long-lasting parasite load and mild (or no) swelling in injection site.Therefore,it seems that in L.major infected BALB/c mice,the high immune response (both cellular and antibody responses) is the causative factor of the severe pathogenesis and if an agent (such as a good vaccine) decreases this intense immune response,the mice would heal.Different from L.major infection,L.tropica infected BALB/c mice shows relatively low immune response resulting in slow pathogenesis,and the mice finally heals spontaneously.Therefore,a protective agent should be able to increase the immune response (both cellular and antibody responses) to accelerate this spontaneous healing process.
There may be some differences between the immune response to a pure single protein and whole-cell antigens.In the present study,we applied a recombinant protein as a pure single protein to show these differences.Our results showed that higher levels of antibodies are induced in mice vaccinated by the recombinant protein,but the pattern of its humoral response is not different from the whole-cell antigen.The higher level of antibody induced by the recombinant protein than SLAs suggests the presence of inhibitory elements in whole-cell antigen(SLA) that suppress humoral responses raised by some Leishmania antigens.In other words,when pure antigen of rLtSTI1 was used,a high level of antibody was evoked due to the absence of these inhibitory elements.Furthermore,the results showed that the protection and high IgG1 response in the rLtSTI1+MPLA group remain unchanged until 16 weeks after the challenge,suggesting this combination(rLtSTI1+MPLA) can be used as a potential vaccine.These findings will be helpful to the future study regarding the Leishmania infection and more studies are needed for understanding the complex profile of immunity against L.tropica infection.
Conflict of interest statement
The authors declare that there are no competing interests.
Funding
This work was supported by Pasteur Institute of Iran (Research Project No.754) and Kermanshah University of Medical Sciences.
Authors’ contributions
MR and HMN contributed to the study design,material preparation,data collection and analysis.The first draft of the manuscript was written by MR.Both authors read and approved the final manuscript.
 Asian Pacific Journal of Tropical Biomedicine2020年12期
Asian Pacific Journal of Tropical Biomedicine2020年12期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Ferulic acid alleviates lipopolysaccharide-induced depression-like behavior by inhibiting inflammation and apoptosis
- Coconut oil nanoemulsion attenuates methotrexate-induced hepatotoxicity and nephrotoxicity in Ehrlich ascites carcinoma-bearing mice
- Rotula aquatica Lour.inhibits growth and biofilm formation of clinically isolated uropathogenic Escherichia coli
- Potential bioactive phytochemicals,antioxidant properties and anticancer pathways of Nymphaea nouchali
- Teicoplanin is a potential inhibitor of SARS CoV-2 replication enzymes: A docking study