
不同建群蜂数对松褐天牛肿腿蜂繁育效果的影响*
2020-10-17 03:03唐艳龙王丽娜张彦龙吴胜勇王小艺杨忠岐
林业科学 2020年9期
唐艳龙 王丽娜 张彦龙 吴胜勇 王小艺 杨忠岐
(1. 遵义师范学院生物与农业科技学院 贵州省赤水河流域动物资源保护与应用研究重点实验室 遵义 563002; 2. 中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林保护学重点实验室 北京 100091)
应用天敌昆虫花绒寄甲(Dastarcushelophoroides)、管氏肿腿蜂(Sclerodermusguani)和川硬皮肿腿蜂(S.sichuanensis)防治松褐天牛(Monochamusalternatus) 是防控松材线虫(Bursaphelenchusxylophilus)病的有效途径(Yangetal., 2014; 杨忠岐等, 2018)。松褐天牛肿腿蜂 (S.alternatusi)与上述2种肿腿蜂同属膜翅目(Hymenoptera)肿腿蜂科(Bethylidae)硬皮肿腿蜂属(Sclerodermus),是2010年于云南发现的寄生松褐天牛幼虫的寄生蜂(杨忠岐等, 2014; Jiangetal., 2015)。该蜂对松褐天牛的寄生率较高,成虫个体较大,扩散能力、攻击能力和搜索能力强,具有较好的应用前景。
当前,人工大量繁育肿腿蜂时的主要问题是替代寄主较难获得,繁育成本较高。在替代寄主有限的情况下,如何提高替代寄主的利用效率尤其重要。对于肿腿蜂这类多头雌蜂可以在同一寄主上产卵的寄生蜂而言,适当增加母蜂的数量,可以显著提高繁蜂效率:一是可以显著缩短子代发育历期,二是可以显著增加子代数量。杨文波等(2018)对管氏肿腿蜂的研究表明,随母蜂数量的增加,其子代幼虫发育历期和蛹历期均缩短,对其子代数量亦有显著影响。对白蜡吉丁肿腿蜂(S.pupariae) 的研究表明,随母蜂数量增加,其子代幼虫历期显著缩短(Gaoetal., 2016),子代数量显著增加(Weietal., 2017)。不同母蜂数量对松褐天牛肿腿蜂的子代发育历期和子代数量等有何影响未见报道。因此,本文对此开展研究,以期为松褐天牛肿腿蜂的繁育利用提供参考。
1 材料与方法
1.1 试验材料和条件
松褐天牛肿腿蜂由国家林业和草原局森林保护学重点实验室提供。替代寄主麻天牛(Thyestillagebleri)幼虫由花鸟市场购得。试验用恒温恒湿光照培养箱控制温湿度(宁波赛福实验仪器PRX-450B),温度为27 ℃,湿度为50%~70%,光周期为短光照,即L∶D=8∶16,光照强度为3 000 lx。
1.2 试验方法
接蜂前,选取健康活跃的麻天牛幼虫用作试虫,用梅特勒电子天平称取体质量介于0.25 g~0.30 g之间的天牛幼虫(绝大多数麻天牛幼虫体质量介于这个区间),装入玻璃指形管中(直径10 mm×高50 mm,有底无盖)并编号,每管1头。
选取健康活跃大小一致且经过交配 (观察到管内有雄蜂与雌蜂进行了交配)的松褐天牛肿腿蜂雌蜂接种,分别接入1~6头雌蜂,之后用医用脱脂棉堵口,保证松褐天牛肿腿蜂雌蜂不能钻出。每个处理重复30次。置于上述培养箱中培养观察。
试验观察记录松褐天牛肿腿蜂雌蜂第一次攻击寄主的时间,松褐天牛肿腿蜂麻痹寄主所需时间,松褐天牛肿腿蜂开始产卵为止,卵孵化的时间,老熟幼虫开始结茧的时间,蛹开始羽化的时间(分雌雄分别记录),最后统计雌蜂产卵前期,子代卵历期、幼虫历期、蛹历期和世代发育历期。待大多数雌蜂开始产卵时,将每个处理中未产卵的雌蜂用毛笔小心去除,重新统计每个处理的建群蜂数。判断标准: 产卵中的雌蜂腹部节间膜明显伸长(周祖基等, 1997),且绝大多数时间待在天牛体上,未产卵的雌蜂节间膜无明显变化,大多数时间待在棉球上。子代雌蜂和雄蜂均羽化后,计数子代雄蜂数量,子代有翅雌蜂数量和子代无翅雌蜂数量,计算出子代雌蜂数量、子代数量、子代雄性比和子代雌蜂有翅率。子代雄性比=100%×(子代雄蜂数量/子代数量)、子代雌蜂有翅率=100%×(子代有翅雌蜂数量/子代雌蜂数量)。
松褐天牛肿腿蜂第一次攻击寄主的时间是指从接入雌蜂开始到雌蜂第一次刺蜇寄主为止,单位为分钟(min), 每20 min观察一次。松褐天牛肿腿蜂麻痹寄主所需时间是指从肿腿蜂第一次刺蛰寄主开始,到寄主完全僵直不动为止,单位为分钟(min), 每20 min观察一次。产卵前期是指从寄主完全僵直开始到肿腿蜂产下第一粒卵为止。卵历期指从松褐天牛肿腿蜂雌蜂开始产卵到卵孵化的时间。幼虫历期是指从松褐天牛肿腿蜂卵开始孵化到子代幼虫变为老熟幼虫的时间。子代雄/雌蜂蛹历期是指从松褐天牛肿腿蜂子代老熟幼虫开始结茧到蛹羽化的时间,分雌雄进行观察。子代发育历期是指松褐天牛肿腿蜂雌蜂产下第一粒卵的时间到子代蛹羽化的时间。产卵前期、卵历期、幼虫历期和蛹历期单位为天,每天观察2次,分别为早上8点与晚上8点。
1.3 数据的统计及处理分析
采用Excel 2010、SPSS进行数据处理和单因素方差分析,利用LSD法多重比较各处理间的差异水平。
2 结果与分析
2.1 不同建群蜂数对松褐天牛肿腿蜂母蜂寄生行为的影响
松褐天牛肿腿蜂第一次攻击寄主的时间随建群蜂数的增加而减少,6个处理之间差异显著(F=7.801; df=5,142;P=0.000 1)(图1A)。当建群蜂数为1头时,松褐天牛肿腿蜂第一次攻击寄主的时间最长,平均为253.9 min。建群蜂数为6头时,其第一次攻击寄主的时间最短,平均只有87.5 min。
松褐天牛肿腿蜂完全麻痹寄主所需时间随建群蜂数的增加略有增加,但6个处理间无显著差异(F=1.197; df=5,97;P=0.316 8)(图1B)。在通常情况下,寄主被蛰后,最快只需要20 min即被麻痹,多数在2 h左右,最长的需要24 h左右。
建群蜂数对松褐天牛肿腿蜂的产卵前期有影响,6个处理之间差异显著(F=5.731; df=5, 147;P=0.000 1)(图1C)。建群蜂数为1头时,松褐天牛肿腿蜂产卵前期最长,平均为8.4天,当建群蜂数为2~6头时,其产卵前期显著变短,均在7.5天左右,5个处理之间差异不显著。
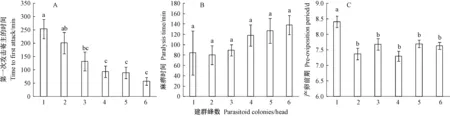
图1 不同建群蜂数下松褐天牛肿腿蜂母蜂第一次攻击寄主的时间(A)、麻痹寄主的时间(B)和产卵前期(C)Fig.1 Time to first attack(A), host paralysis time(B) and pre-oviposition period(C) of S. alternatusi female with different parasitoid colonies表中数据为平均值±标准误,不同小写字母表示在5%水平差异显著,下同。Data are mean ± SE, different lowercase letters indicate significant difference (P<0.05), the same below.
2.2 不同建群蜂数下松褐天牛肿腿蜂子代发育历期
不同建群蜂数下松褐天牛肿腿蜂子代卵历期无明显差异,平均为4.5~4.8天 (F=1.460; df=5,154;P=0.206 3)(表1)。幼虫历期与建群蜂数显著负相关 (F=17.007; df=5,154;P=0.000 1),即建群蜂数越少,幼虫历期越长,当建群蜂数为1头时,其幼虫历期最长,平均为9.6天,6头时,其幼虫历期最短,平均为8.5天,相差约1天。
建群蜂数对松褐天牛肿腿蜂雄蜂蛹历期、雌蜂蛹历期、雄蜂发育历期和雌蜂发育历期的影响类似,4个指标均随建群蜂数的增加呈下降趋势。雄蜂蛹历期介于18.7~19.7天之间,6个处理间差异显著(F=5.688; df=5,154;P=0.000 1)(表1)。雌蜂蛹历期比雄蜂蛹历期长1~2天,平均为20.0 ~21.5天之间,6个处理间差异显著(F=6.895; df=5,154;P=0.000 1)。雄蜂平均发育历期最长为34.2 天(建群蜂数为1头),最短为31.8天 (建群蜂数为5头),6个处理间差异亦显著(F=12.930; df=5,154;P=0.000 1)。雌蜂平均发育历期比雄蜂长1~2天,最长为35.8天(建群蜂数为1头),最短为32.9天(建群蜂数为5头),6个处理间差异亦显著(F=12.930; df=5,154;P=0.000 1)。

表1 不同建群蜂数下松褐天牛肿腿蜂子代发育历期①Tab.1 Developmental stage of S. alternatusi progenies with different parasitoid colonies d
2.3 不同建群蜂数下松褐天牛肿腿蜂的子代数量
不同建群蜂数对松褐天牛子代数量影响明显(表2)。随建群蜂数的增加,其子代雄蜂数(F=3.288; df=5,154;P=0.000 1)、子代无翅雌蜂数(F=5.020; df=5,154;P=0.000 4)、子代总雌蜂数(F=3.757; df=5,154;P=0.003 8)和子代总数(F=4.261; df=5,154;P=0.001 5)均显著增加。当建群蜂数为1头时,其子代雄蜂数、子代无翅雌蜂数、子代总雌蜂数和子代总数均最少,分别为3.6、46.0、51.1和54.6头。当建群蜂数为2头时,4个指标即显著增加到5.9、58.7、71.3和77.2头,之后随建群蜂数增加,4个指标均逐渐增加,至4头时,即增加到6.6、78.2、83.9和90.4头。5头时,4个指标小幅减少,至6头时,又略有增加。
松褐天牛肿腿蜂子代有翅雌蜂数和雌蜂有翅率变化不规律,总体是有翅雌蜂明显少于无翅雌蜂,其中部分处理全是无翅雌蜂。6个处理,子代雌蜂有翅率分别为10.6%、15.6%、7.2%、6.6%、5.2%和6.6%,均低于20%。

表2 不同建群蜂数下松褐天牛肿腿蜂子代数量Tab.2 Progenies number of S. alternatusi with different parasitoid colonies
2.4 不同建群蜂数下松褐天牛肿腿蜂的子代雄性比
松褐天牛肿腿蜂子代雄性比随建群蜂数的增加呈增加趋势(图2),但不同处理间差异不显著(F=0.920; df=5,154;P=0.4714),当建群蜂数为1头时,其子代雄性比最小,平均为6.3%,2~5头时,相差不大,平均介于7.9%~8.4%之间,6头时,增加到10.0%。

图2 不同建群蜂数下松褐天牛肿腿蜂子代雄性比Fig.2 Male ratio of S. alternatusi progenies with different parasitoid colonies
3 讨论
以往利用替代寄主繁育肿腿蜂探讨最佳接蜂数量的试验,都用接蜂数量直接替代了建群蜂数(周娜等, 2006; 杨希等, 2007; 姚万军等, 2008; 杨文波等, 2018),实际上,在1头替代寄主上接入多头肿腿蜂的时候,往往有一些雌蜂并不会去寄生替代寄主,而这些肿腿蜂雌蜂对子代数量的贡献为零,因此不能用接蜂数量直接代替建群蜂数。目前已经报道的硬皮肿腿蜂属寄生蜂都需要吸食寄主体液补充营养后才能完成产卵(魏可等, 2016; 田春景等, 2019),肿腿蜂在补充营养的过程中,其外部特征变化最明显的是腹部。周祖基等(1997)报道川硬皮肿腿蜂完成补充营养后,其腹部节间膜明显可见,体长可增长50%。本试验中,松褐天牛肿腿蜂补充营养后雌蜂有类似的外部特征变化。因此,试验中根据雌蜂腹部特征的变化来判断母蜂是否会产卵来确定建群蜂数量是可靠的。
肿腿蜂补充营养的时长因温度、产卵蜂数和替代寄主不同而不同,蒋学建等(2007)报道川硬皮肿腿蜂需取食5天才会产卵[替代寄主为黄粉甲(Tenebriomolitor),未报道温度],Gao等(2016)报道白蜡吉丁肿腿蜂的产卵前期为4~6天[25 ℃,替代寄主为麻天牛],管氏肿腿蜂产卵前期5~8天不等(26 ℃,替代寄主为黄粉甲) (杨文波等, 2018)。本文中松褐天牛肿腿蜂的产卵前期为7~8天(27 ℃,替代寄主为麻天牛),且随建群蜂数的增加,其产卵前期缩短,这与Gao等(2016)的结论一致。虽然不同试验中肿腿蜂的产卵前期长短不具有可比性,但说明硬皮肿腿蜂属寄生蜂都需要持续吸取寄主体液补充营养多天才能产卵。在这一过程中,个体较小的寄主有可能会被取食致死。张彦龙等(2012)报道1头松褐天牛肿腿蜂雌蜂在补充营养过程中平均可致死3.75头3龄松褐天牛幼虫,王小艺等(2010)报道白蜡吉丁肿腿蜂在寄生栗山天牛(Massicusraddei)的过程中亦会致死较多幼虫。这表明在检查肿腿蜂的防治效果时,应将肿腿蜂致死的害虫数量计算在内,单纯地用寄生率来评价肿腿蜂的防治效果是不全面的。当然,被致死的害虫往往无迹可寻不便统计,大多只能通过设置对照间接评价(唐艳龙等, 2014)或者不评价,在实际应用中,有时候寄生率不高,就直接认定其防治效果不好是不科学的。
本研究表明,松褐天牛肿腿蜂建群蜂数对其子代幼虫历期有显著影响,表现为随建群蜂数增加,其子代幼虫历期变短。这与Gao等(2016)对白蜡吉丁肿腿蜂的研究和杨文波等(2018)对管氏肿腿蜂的研究结论类似。随着建群蜂数的增加,其子代幼虫数量增加,在有限的资源下,单个幼虫获得的资源变少,其幼虫历期变短。试验结束观察寄主残体时发现,当建群蜂数为1头时,其残体较为明显,而多头建群蜂数时,其残体几乎只剩头壳。这说明在一定的资源下,适当增加建群蜂数,可以提高资源的利用效率(陈然等, 2019)。
建群蜂数对肿腿蜂子代蛹历期的影响值得关注,杨文波等(2018)报道不同蜂虫比会影响管氏肿腿蜂的蛹期(寄主为黄粉甲),而Gao等(2016)报道不同蜂虫比对白蜡吉丁肿腿蜂的蛹期没有影响(寄主为麻天牛),林芳芳等(2015)报道母蜂数量对管氏肿腿蜂的蛹期也没有影响(寄主为松褐天牛)。本试验的研究结果与杨文波等(2018)类似。建群蜂数对子代蛹历期究竟有无影响及其影响机制尚待探索。不过试验发现,松褐天牛肿腿蜂的雄蜂蛹历期比雌蜂蛹历期短1~2天,这与Gao等(2016)对白蜡吉丁肿腿蜂的研究类似,白蜡吉丁肿腿蜂雄蜂蛹期比雌蜂蛹期短1 天左右。因此,试验中发现雄蜂一般比雌蜂早1~2天羽化,羽化后的雄蜂会协助咬破雌蜂茧壳,钻入茧壳与雌蜂交配,因此,在空间受限的环境中,松褐天牛肿腿蜂属完全近交的寄生蜂,这与Tang等(2014)对管氏肿腿蜂的研究一致。
研究表明,松褐天牛肿腿蜂子代数量随建群蜂数的增加而增加。这与Wei等 (2017) 对白蜡吉丁肿腿蜂(寄主为麻天牛)和杨文波等(2018)对管氏肿腿蜂(寄主为黄粉甲)的研究结果类似。在实际应用中,并不是母蜂数量越多越好,Wei等 (2017)表明白蜡吉丁肿腿蜂蜂虫比在4∶1时,既能获得较多的子代数量,又能保证子代的质量。而杨文波等(2018)认为用黄粉甲繁育管氏肿腿蜂的最佳蜂虫比是8∶4。此外,周娜等(2006)报道青杨天牛繁育管氏肿腿蜂的最佳蜂虫比为1∶1,杨希等(2007)报道利用松褐天牛繁育管氏肿腿蜂最佳蜂虫比为3∶1,姚万军等(2008)报道意大利蜜蜂(Apismelliferaligustica)蛹繁育管氏肿腿蜂最佳蜂虫比为2∶1。可见,繁育寄主不同,最佳蜂虫比也完全不同。本试验中,利用麻天牛幼虫繁育松褐天牛肿腿蜂,当建群蜂数为4头时,即能获得较大的子代数量,之后,随建群蜂数的增加,其子代数量没有明显增加。
已有的研究表明,硬皮肿腿蜂属寄生蜂的确会产出大量的雌蜂,而雄蜂数量往往只有几头甚至1头;本试验中发现松褐天牛肿腿蜂亦是如此,松褐天牛肿腿蜂子代雄蜂数量普遍少于10头,大多数处理子代雄性比不到10%。对于具有近交特点的寄生蜂而言,少量的雄蜂即可满足交配,因此,母蜂会倾向于产生更多的雌蜂,但为了确保其后代种群中有雄蜂出现,雄蜂先产显然是最有利的进化稳定性策略。Kinoshita等 (1998)、 Moore等 (2005)、Raja等 (2008)对榕小蜂(Liporrhopalumtentacularis)的性别分配研究证实了寄生蜂存在雄性先产的策略。肿腿蜂母蜂是否会通过先产雄蜂来控制子代性别,松褐天牛肿腿蜂雄蜂先于雌蜂羽化是由其生物学特点决定的还是因为母蜂先产了雄蜂,这些问题还需要更多的试验进行研究。
4 结论
不同建群蜂数对产卵母蜂的寄生行为有影响,表现为随建群蜂数的增加,松褐天牛肿腿蜂母蜂第一次攻击寄主的时间和母蜂的产卵前期均缩短。
不同建群蜂数对松褐天牛肿腿蜂子代发育历期有显著影响,随建群蜂数增加,其子代幼虫历期和蛹期均显著缩短,但卵历期无差异。
随建群蜂数的增加,松褐天牛肿腿蜂子代数量显著增加,建群蜂数为4头时,其子代数量接近最大,因此,利用麻天牛作为繁育寄主生产松褐天牛肿腿蜂时,最佳接蜂数量为4头。松褐天牛肿腿蜂子代性别总体高度偏雌,大多数处理子代雄性比低于10.0%,随建群蜂数的增加,其子代雄性比呈小幅增加趋势。
猜你喜欢
湖北农业科学(2021年22期)2021-12-13
南方林业科学(2021年3期)2021-07-28
农家科技下旬刊(2020年5期)2020-06-19
吉林农业(2019年7期)2019-04-01
中国蜂业(2019年9期)2019-01-07
中国森林病虫(2017年2期)2017-12-18
中国稻米(2017年2期)2017-04-28
中国茶叶(2016年7期)2016-01-18
意林(2015年1期)2015-05-14
中国蜂业(2012年8期)2012-08-15
