
木荷优树自由授粉家系早期生长性状遗传变异动态规律*
2020-10-17 04:57王云鹏周志春黄少华马丽珍范辉华
林业科学 2020年9期
王云鹏 张 蕊 周志春 黄少华 马丽珍 范辉华
(1. 中国林业科学研究院亚热带林业研究所 浙江省林木育种技术重点实验室 杭州 311400; 2. 南京林业大学 南京 210037; 3. 福建省建瓯市林业技术推广中心 建瓯 353100; 4. 福建省林业科学研究院 福州 350012)
遗传变异是林木遗传改良的基础,根据林木的遗传变异规律,通过不同的育种手段,可达到目标性状改良的目的(郑勇奇, 2001)。遗传评价与选择是林木遗传改良的重要环节,通过子代测定,可选择优良家系和单株(欧阳芳群等, 2018; 林能庆, 2013),用于建立种子园和生产造林(冯源恒等, 2017)。但林木生长周期长,而且在不同季节和不同年龄表达的基因不同,以及生长性状早晚相关的不确定性(怀特等, 2013),给育种策略的科学制定带来了难度。树高和胸径是林木遗传改良的典型目标性状,受多基因控制,每年都有一定的生长量,且在生长过程中存在一定的动态规律(孙晓梅等, 2004; 韩东花等, 2019)。在林木育种计划中,准确估算性状的遗传参数对育种策略的制定至关重要,根据遗传力和早晚相关随年龄增长的变化趋势,可以确定早期选择年龄(Svenssonetal., 1999)。Diao等(2016)对日本落叶松(Larixkaempferi)自由授粉家系遗传参数和早期选择研究发现,树高和胸径的单株、家系遗传力和早晚相关随年龄增长表现出一定的动态变化,确定树高和胸径的最佳早期选择年龄分别为4年和5年。任华东等(2010)对黑荆(Acaciamearnsii)半同胞家系研究发现,树高和胸径的早晚相关主要受遗传控制,以胸径作为早期选择指标,在5~9年时对家系进行早期选择效果最佳。由此可见,利用多年的观测数据,分析生长性状随年龄增长的变化动态,可充分了解林木性状在一定时期内的遗传变异规律,有效减小育种的盲目性,从而提高选择效率。因此,研究林木的生长性状随年龄变化的动态,是林木遗传改良进程中必不可少的步骤。
木荷(Schimasuperba)为山茶科(Theaceae)木荷属(Schima)常绿阔叶大乔木,是中国亚热带常绿阔叶林的主要建群种,是我国南方最主要的珍贵优质用材、高效生物防火、生态景观和生态防护造林树种(周志春, 2019)。据前期研究,木荷1年生自由授粉家系的生长性状在家系间遗传差异极显著,且存在明显的种源效应(林磊等, 2009); 5年生时,同样存在极显著的家系效应(辛娜娜等, 2014)。然而木荷自由授粉家系的生长性状随年龄变化的遗传变异规律和早晚相关性还未见报道,这制约了木荷遗传改良的早期选择及高世代育种的推进。鉴于此,本文利用木荷优树自由授粉家系3、5和10年生时主要生长性状的观测值,剖析家系生长性状的遗传变异随年龄增长的变化规律及性状的早晚相关性,以期降低在早期进行选择时产生的偏差,为木荷育种策略的制定和改良效率的提高提供理论依据。
1 材料与方法
1.1 试验材料和设计
木荷优树家系测定林建于福建建瓯(117°58′—118°57′E,26°38′—27°20′N),年平均气温18.9 ℃,1月平均气温8 ℃,7月平均气温28.6 ℃,年降水量1 627.3~1 662.9 mm,无霜期286天,海拔140~180 m。测定林家系来源于2006年在浙江龙泉、庆元、遂昌和福建建瓯的木荷优良天然林中所选的优树。所选的优树树干通直圆满,无病虫害,高、径生长明显大于周围的3~5株对比木。2008年春用1年生裸根苗造林,家系随机分成3个试验组,3个试验组的家系个数分别为47、45和45,每个试验组均按随机完全区组设计,各5次重复,10株单列小区,株行距为2 m×2.5 m。
1.2 数据调查和处理
分别于2010年、2012年和2017年底进行全林调查。调查指标包括树高(H)和胸径(DBH)等。选用保存较完好的第1组47个家系(表1)的测定数据用于统计分析。采用SAS 8.0软件的MEAS过程进行描述性统计,GLM过程进行性状双因素方差分析,CORR过程计算表型早晚相关,PROC VARCOMP过程中的REML方法计算各方差分量,进而估算各遗传参数。
单性状分析使用混合线性模型:Yijk=μ+Bi+Fj+BFij+Eijk,式中:Yijk为第i区组第j家系第k单株的观测值;μ为群体平均值;Bi为第i区组效应;Fj为第j家系效应;BFij为第i家系和第j区组的互作效应;Eijk为机误。此外,μ和Bi为固定效应,其余为随机效应。





早期选择效率:E=(hj/hm)·(tm/tj)·rgjm,
式中:hj和hm分别为早期性状和晚期性状的遗传力平方根,rgjm为早晚遗传相关系数,tj为早期年龄,tm为晚期年龄。
根据Francis & Kannenberg(1978)基因型分组方法(grouping genotype),以年度间平均生长量的变异系数为横坐标,年度间平均生长量为纵坐标,变异系数的平均值和平均生长量的平均值为中心原点作图,对家系进行分类评定。
平均生长量:θ(tj,tm)=(ym-yj)/(tm-tj)(孟宪宇, 2006),式中:ym为性状晚期的生长量,yj为性状早期的生长量。
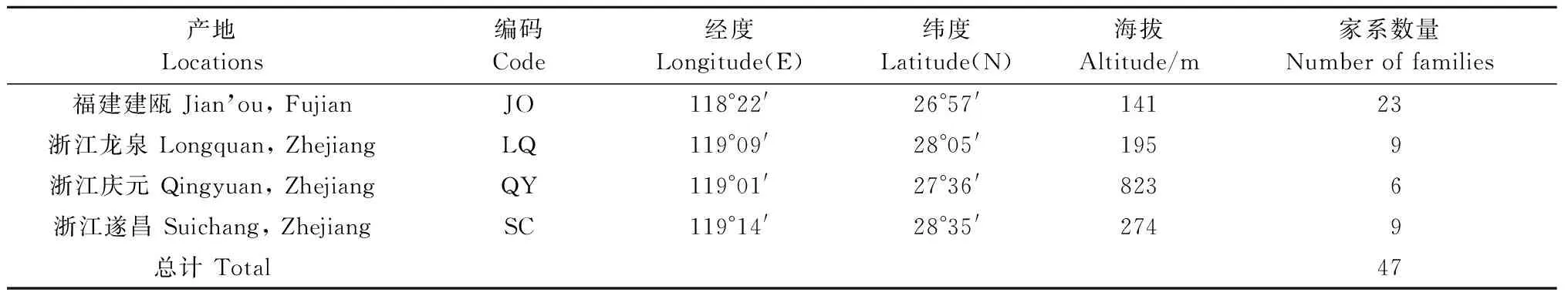
表1 参试家系的来源及数量Tab.1 The source and number of families tested in the experiment
2 结果与分析
2.1 不同年龄家系生长性状变异
木荷优树自由授粉家系树高和胸径在不同年龄时的均值差异较大(表2)。5年生树高和胸径相较于3年生分别增加了102.64%和111.11%,10年生树高和胸径相较于5年生分别增加了74.35%和104.99%,树高和胸径在3~5年间的生长量大于5~10年,说明树高和胸径的幼龄期生长量较大;树高和胸径在3~5年生间的生长率分别为50.65%和52.63%,在5~10年间的生长率分别为42.64%和51.22%,可见胸径的生长率大于树高的生长率。方差分析结果显示,在不同年龄,木荷优树自由授粉家系的树高和胸径在家系间皆存在极显著差异,且家系的遗传效应随年龄增长而增强。3、5和10年生的树高均值最大的家系比最小的家系分别高出37.88%、39.64%和49.92%,胸径均值最大的家系比最小的家系分别高出98.18%、48.83%和107.16%,说明在不同年龄,家系间胸径的变异较大。这种胸径的高生长率及其显著的变异性,为木荷的目标育种提供更多可选择的优良材料。
为更清晰地反映家系间树高和胸径的动态变化,分别从3年生树高和胸径的变化范围内,选择高、中和低的各2个家系,进行家系间生长趋势的分析(图1)。结果显示,3年生时树高和胸径处于优势的家系,在后续的生长中一直处于优势,如LQ19和JO49; 处于劣势的家系,在后续的生长中也一直处于劣势,后续很难超越优势的家系,如SC21、QY6和SC9; 而早期处于中间的家系,在后续的生长中,有的变为优势,有的变为劣势。如LQ7的胸径在3和5年生比处于优势的JO56分别低19.05%和4.81%,而到10年生高了7.37%; JO33的树高在3和5年生分别比处于劣势的JO63高4.39%和3.89%,而到10年生低了8.90%。
不同产地之间木荷家系的树高随年龄增长,差异从不显著变为极显著,而胸径则始终表现为极显著差异(图2)。树高按产地均值排序: 3年生时,庆元>建瓯>龙泉>遂昌; 5年生时,建瓯>庆元>龙泉>遂昌; 10年生时,建瓯>龙泉>庆元>遂昌。庆元产地家系的树高随年龄增长,长势减缓,而建瓯和龙泉产地的家系却长势增强,这可能与庆元产地的家系来自高海拔有关。在不同年龄,不同产地间家系的胸径生长较稳定,建瓯>龙泉>庆元>遂昌。说明树高和胸径的生长显著受到产地影响。
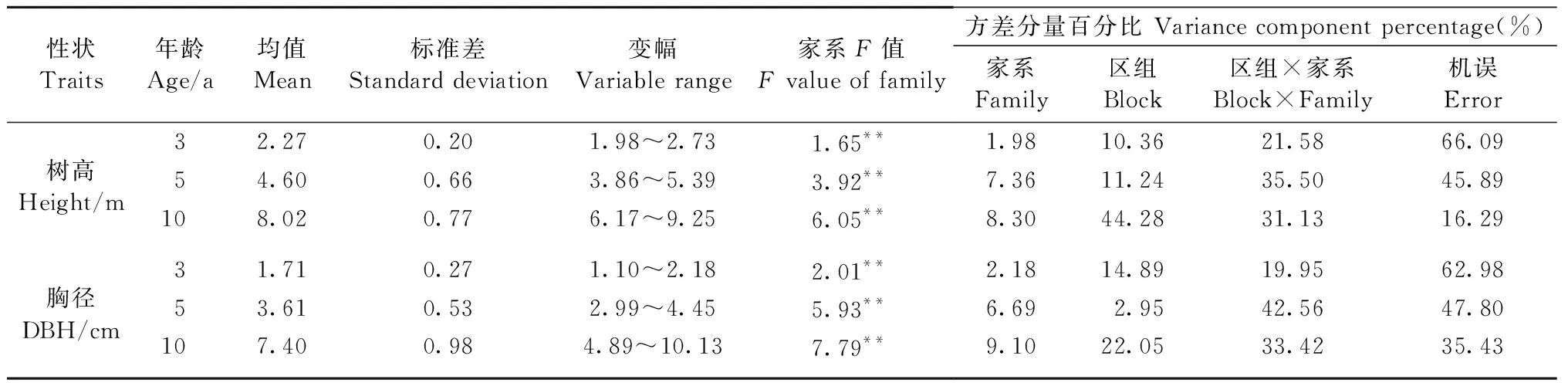
表2 不同年龄家系树高和胸径的统计和方差分析①Tab.2 Statistics and variance analysis of height and DBH for families in different ages

图1 典型家系树高和胸径随年龄变化趋势Fig.1 Age trends in mean of height and DBH for typical families

图2 不同产地家系树高和胸径随年龄变化趋势Fig.2 Age trends in mean of height and DBH for different families at different locations
2.2 不同年龄家系生长性状的遗传力和变异系数
不同年龄木荷优树家系树高和胸径的家系遗传力和单株遗传力随年龄增长而增高(图3)。3、5和10年生胸径的家系遗传力分别为0.39、0.42和0.70,树高的家系遗传力分别为0.23、0.59和0.62,都受中等偏强的遗传控制。树高生长在5~10年生时受遗传控制较强且相对稳定,而胸径的遗传力在3~5年生时变化相对平稳,但在10年生时受遗传控制较强。说明树高和胸径在不同生长阶段所受到的遗传控制强弱不同,且二者各自所受遗传控制的平稳期出现在不同年龄段。单株遗传力和家系遗传力有相似的变化趋势,3和5年生时树高和胸径的单株遗传力较低,但到10年生时单株遗传力在0.5以上,说明10年生时在选择家系的同时,可配合单株的选择。另外,3年生和10年生时胸径的遗传力高于树高,说明胸径受到的遗传控制强于树高,利用胸径进行早期选择更为可靠。
在不同年龄,家系树高和胸径的表型变异系数大于遗传变异系数(图4)。3~10年间,胸径的表型变异系数下降了2.46%,遗传变异系数提高了2.35%,这表明随年龄增长,胸径的表型变异减小,受遗传效应控制的变异增强;10年生时木荷优树自由授粉家系胸径的遗传变异系数高出树高44.87%,说明胸径因遗传效应引起的变异较大。而树高的表型变异系数和遗传变异系数相对于胸径处于波动状态,随年龄增长,树高的表型变异系数和遗传变异系数先升高后降低,说明在3~10年间树高受遗传控制引起的变异趋势不稳定。因此,以胸径为指标,结合表型变异开展早期选择意义更大。

图3 家系树高和胸径的遗传力随年龄变化趋势Fig.3 Age trends in heritability of height and DBH for families
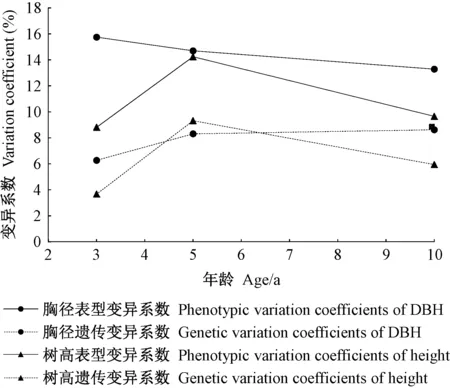
图4 家系树高和胸径的变异系数随年龄变化趋势Fig.4 Age trends in variation coefficients of height and DBH for families
2.3 家系生长性状的早晚相关
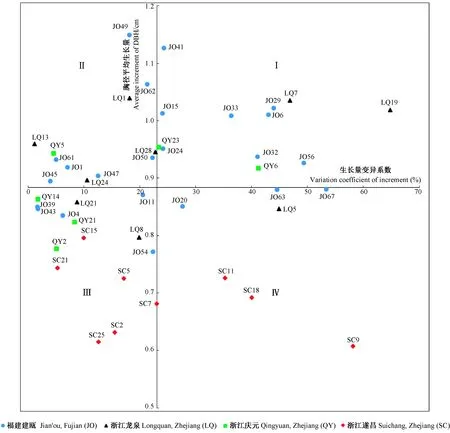
图5 不同家系胸径生长量稳定性Fig.5 Stability of DBH increment in different families
木荷优树自由授粉家系10年生树高和胸径分别比3年生增长253.30%和332.75%,分别比5年生增长74.35%和104.99%(表2),且10年生是1/3轮伐期,因此与10年生进行早晚相关分析对木荷家系早期选择具有重要意义。结果表明,3年生与10年生,及5年生与10年生的树高和胸径皆表现出显著的表型正相关和遗传正相关(表3),表型相关系数在0.20~0.59之间,遗传相关系数在0.26~0.80之间,遗传相关系数均大于表型相关系数,表明树高和胸径的早晚相关主要受遗传控制。表型和遗传的早晚相关系数随年龄增长而增大,树高和胸径5年生与10年生的相关系数均大于3年生与10年生的相关系数,表明5年生较3年生更能准确地表达10年生时的信息。另外,胸径的表型和遗传早晚相关均大于树高,因此胸径作为早期选择性状更可靠。

表3 家系树高和胸径的早晚相关Tab.3 Juvenile-mature correlation of height and DBH for families
2.4 不同家系的生长量及稳定性评价
基于以上结果,胸径受到的遗传控制强于树高,且其早晚相关也强于树高,因此选择胸径作为评价指标对47个家系进行生长量及稳定性评价。通过基因型分组法,可将家系划分为4类(图5),第Ⅰ类为高生长量低稳定性,第Ⅱ类为高生长量高稳定性,第Ⅲ类为低生长量高稳定性,第Ⅳ类为低生长量低稳定性。4类家系分别占全部家系的25.53%、25.53%、34.04%和12.77%,可见第Ⅲ类低生长量高稳定性的家系占比最多;建瓯和龙泉产地的家系分别在第Ⅰ类和第Ⅱ类中占比最大,分别占比17.02%和8.51%,而庆元和遂昌产地的家系均在第Ⅲ类中占比最大,分别占比6.38%和12.77%。最终选出胸径高生长量和高稳定性的JO49、JO62和LQ1等12个家系(表4)。另外,结合高生长和高稳定家系的选择,同时避免单株之间产生近交衰退,从入选的每个家系中选择1株胸径较大的单株,共筛选出12株高生长量高稳定性的优良单株。

表4 高生长量高稳定性家系Tab.4 Families of high growth and high stability
3 讨论
3.1 家系生长性状的遗传变异规律
林木的遗传变异是遗传改良的前提,有效的遗传变异决定了物种在育种过程中的改良潜力,掌握林木的遗传变异规律,是育种策略制定的基础。本研究结果表明,木荷优树自由授粉家系在3、5和10年生时,树高和胸径在家系间皆存在极显著的变异,这种变异随年龄的增长而增大。木荷1年生自由授粉家系的苗高和地径也存在显著的家系效应(林磊等, 2009),因而家系选择是木荷遗传改良的重要途径。本研究还发现,随年龄增长,不同产地家系的树高生长处于波动状态,庆元产地家系的树高生长在3年生时处于优势,而到10年生时长势减缓,变为劣势,这可能与庆元产地的家系来自高海拔(海拔800 m左右)有关,高生长早期表现较好(周志春等, 2006)。而不同产地家系的胸径生长较稳定,建瓯和遂昌产地家系的胸径生长分别一直处于优势和劣势。说明不同产地家系树高和胸径的生长势变化与产地的环境因子有密切关系,这也与木荷种源区划分的研究结果(张萍等, 2006; 王秀花等, 2011)一致,因此,在家系选择时要注意产地的影响,充分利用产地效应。
林木在生长过程中,由于各树种的特性差异,导致其生长规律不同。范少辉等(2008)对26年生小黑杨(Populus×xiaohei)人工林生长特性进行研究,根据树高、胸径和材积的生长量将其分为幼林期、速生期、近熟期和成熟期,发现树高生长早于胸径。金国庆等(2011)对5~25年马尾松(Pinusmassoniana)种源林研究发现,在胸径、树高和材积3个主要生长性状中,胸径的速生期出现最早。张振等(2017)发现2~5年马尾松全同胞家系林幼林期生长性状以树高的生长量最大,5年生较2年生增长了4.23倍。由此可见,性状的生长量不仅取决于测定树种,还取决于所研究的林分。本研究结果表明,树高和胸径的幼龄期生长量较大,在3~5年和5~10年,胸径的相对生长量均大于树高,说明木荷优树自由授粉家系在早期生长中,胸径的生长率大于树高。对典型家系树高和胸径的生长动态分析结果表明,早期生长较慢的家系在后续的生长过程中一直处于劣势,很难超越早期生长处于优势的家系; 早期生长较快的家系,在后续的生长中也不会出现衰退的现象; 而早期生长处于中等的家系,在后期生长中会表现出优势或劣势的现象。此结果与火炬松(Pinustaeda)(Svenssonetal., 1999)、日本落叶松(孙晓梅等, 2004)、黑荆(任华东等, 2010)典型家系生长趋势的研究结果一致,这表明在木荷优树自由授粉家系的早期进行选择或淘汰是有意义的。
遗传参数的正确评估对预期增益估计和育种策略制定尤为重要(White, 1996)。遗传力是指通过选择后,能为下一代带来的增益比例,反映性状受遗传效应控制的强弱。日本落叶松家系在4~18年间树高和胸径的家系和单株遗传力随年龄增长呈一定的动态变化,家系遗传力大于单株遗传力(Diaoetal., 2016)。任华东等(2010)发现3~15年生黑荆半同胞家系树高和胸径的家系遗传力均在0.7以上,单株遗传力在0.52~0.64之间,受较强的遗传控制,遗传力变化比较平稳。本研究发现,随着年龄的增长,木荷树高和胸径的家系遗传力和单株遗传力有上升的趋势,树高在5和10年生时受遗传控制较强,而胸径在10年生受遗传控制较强,整体而言,胸径所受到的遗传控制强于树高。在10年生时树高和胸径的单株遗传力较高,因此在10年生选择家系的同时,可配合单株选择,如此可利用这种高遗传力性状的加性遗传方差,获得最大化的遗传增益。在本研究中3年生时树高和胸径的家系和单株遗传力估值(0.09~0.39)偏小,这与1年生木荷自由授粉家系苗高和地径的家系和单株遗传力估值(0.28~0.45)偏小一致,受中等偏弱的遗传控制(林磊等, 2009),可能原因是在幼龄期性状受到的遗传控制较小,家系间基因型的差异还未完全表达。对不同年龄木荷自由授粉家系树高和胸径的变异系数分析发现,随年龄增长,胸径的表型变异系数降低,遗传变异系数升高,而树高的表型和遗传变异系数先升高后降低,处于波动状态,意味着在不同年龄,树高和胸径受遗传效应引起变异的变化趋势是不同的。此外,除5年生外,胸径的表型和遗传变异系数均大于树高,在一定程度上说明胸径受遗传效应引起的变异强于树高。此外,研究遗传参数的动态变化需要对试验林长期持续地观测,本研究用了3个时间点的观测值进行分析,后续还应加强对试验林的跟踪观测,以获得更为可靠的数据支持。
3.2 家系生长性状的早期选择
由于林木生长周期长,在很大程度上限制了林木遗传改良的进程,早期选择对于缩短育种周期和加速高世代育种具有重要作用(杨秀艳等, 2004)。生长性状的早晚相关是早期选择的途径之一,通过生长性状早晚相关的分析方法,胡德活等(2001)认为杉木(Cunninghamialanceolata)无性系的早期选择最佳年龄在3~4年之间; Xiang等(2003)发现火炬松树高和胸径的早期选择年龄分别在3年和4年,胸径的早晚相关性高于树高; Lai等(2014)认为日本落叶松无性系树高和胸径的早期选择年龄分别为2年和5年,不同年龄胸径与材积的遗传相关大于树高。但也有研究存在不同的观点,如王章荣等(1987)利用树干解析法,认为马尾松人工林早期以树高、后期以胸径为选择指标; 童再康等(1993)以2~8年生黄山松(Pinustaiwanensis)为材料测得树高的遗传较稳定,以28年生和45年生解析木为材料确定黄山松的最佳早期选择年龄在8~10年; 季孔庶等(2005)认为马尾松自由授粉家系的选择应注重树高生长量。这些研究意味着早期选择因子和最佳年龄的确定取决于测定材料和方法(Svenssonetal., 1999),不同树种的早期选择存在差异性。树干解析法只适用于单株,而研究家系的早晚相关必须有若干年的数据积累,本研究结果表明,木荷优树自由授粉家系树高和胸径的表型和遗传的早晚相关均存在显著性,且主要受遗传效应的控制。5年生与10年生的表型和遗传相关系数皆大于3年生与10年生的相关系数,因此5年较3年更适合作为早期选择年龄。而且3年生与10年生,以及5年生与10年生的胸径表型和遗传相关系数均大于树高,因此胸径作为早期选择性状较树高更能准确地表达家系10年生时的信息。早期选择效率是确定早期选择年龄的主要依据,当早期选择效率大于1时,说明早期选择是有效的,早期选择效率函数的拐点为早期选择的最佳年龄。本研究发现家系在5年生时树高和胸径的早期选择效率分别为1.07和1.19,因此进行早期选择是有效的。而且发现以胸径作为早期选择性状的最佳早期选择年龄比树高出现得早,如果以树高作为早期选择性状,最佳早期选择年龄要推迟,更准确可靠地确定早期选择最佳年龄还需持续地观测并在下一育种周期加以验证。
3.3 家系生长的稳定性评价
林木的稳定性是指在不同环境或不同时期能够调节其表型的特性,使之能适应环境变化,维持其平稳的生理和生殖特性,从而保持稳定状态(陈天华等, 1994)。Francis和Kannenberg(1978)建立的基因型分组法在一定程度上能反映表型稳定性信息,具有直观且计算简便的优点(李火根等, 1997)。本研究利用此方法基于胸径年度间平均生长量和生长量变异系数初步对47个木荷家系的生长量及稳定性进行分类评价,划分为4类: 高生长量高稳定性、高生长量低稳定性、低生长量高稳定性和低生长量低稳定性。其中,高生长量高稳定性的JO49、JO62和LQ1等12个家系适应性较强,同时筛选出的12株优良单株,可作为下世代育种亲本。在后续的研究中,还需进行多点试验,以揭示和验证这些家系的基因型与环境的互作效应。
4 结论
随年龄增长,木荷优树自由授粉家系生长性状的遗传效应增强,同时也存在明显的产地效应,因此应注重优良产地内的家系选择。在幼龄期,树高和胸径的生长量较大,且胸径的生长率大于树高,典型家系树高和胸径的变化趋势显示,在早期进行选择是可靠的。树高和胸径随年龄增长受遗传控制增强,在5年生后受较高水平的遗传控制,胸径受遗传控制强于树高,且具有较强的变异性。树高和胸径的早晚相关主要受遗传因素的控制,胸径作为早期选择性状更可靠,5年生较3年生更能表达10年生时的信息。综合遗传力、变异系数和早晚相关系数,认为在5年生时以胸径作为早期选择性状选择效率更高。选择出12个高生长量高稳定性家系和12株高生长量高稳定性单株。
猜你喜欢
养猪(2022年4期)2022-08-17
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
花卉(2021年2期)2021-01-21
林业与环境科学(2020年3期)2020-07-21
农民致富之友(2017年4期)2017-04-10
现代农业科技(2017年4期)2017-04-10
绿色科技(2017年1期)2017-03-01
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
