
基于病毒组件的纳米材料的自组装合成、功能化及应用
2020-10-10 06:34张文静李明周维张先恩李峰
合成生物学 2020年3期
张文静,李明,周维,张先恩,李峰
(1中国科学院武汉病毒研究所,生物安全大科学研究中心,病毒学国家重点实验室,湖北武汉430071;2中国科学院生物物理研究所,生物大分子科教融合卓越中心,生物大分子国家重点实验室,北京100101;3中国科学院大学,北京100049)
生物体系中存在大量精巧的生物大分子组装体纳米机器,它们参与生命周期的各个过程,对生命过程的正常发生至关重要。研究这些天然纳米机器的组装与功能,促进人们对生物大分子组装体的理解以及再设计、再合成、再利用。其中,病毒是一类典型的生物大分子组装体,具有纳米尺度的大小、精确的三维结构,执行明确的生物学功能,是通过分子自组装合成纳米材料的天然范例,吸引了越来越多不同领域的学者和开发者的注意。病毒学与纳米技术的交叉,产生了病毒纳米技术这一新兴领域。围绕基于病毒颗粒的纳米材料,人们发展了基因工程、化学修饰、组装、矿化等各种不同的功能化改造手段,进而应用于生物递送、免疫学、生物传感、酶学、微电子学等各个领域。
1 病毒的结构
病毒是最简单的生命形式,普遍存在于不同等级的生物体中,按照其感染对象,可分为动物病毒、植物病毒、细菌病毒(又称“噬菌体”)和真菌病毒。病毒的基本结构由蛋白质外壳(即衣壳)和封装于其中的基因组核酸组成,包膜病毒衣壳外部还具有磷脂膜结构。病毒衣壳按形态可分为球形、棒状、丝状、蝌蚪状等(图1)。球形病毒衣壳具有正二十面体对称性,而棒状及丝状病毒衣壳具有螺旋对称性,蝌蚪状病毒具有复合对称性,如T4噬菌体既有正二十面体对称性的衣壳头部,又有螺旋对称性的尾鞘。正二十面体病毒衣壳的蛋白质亚基数量和排布可以由三角剖分数(T)表示,T越大粒径越大[1]。很多病毒的衣壳蛋白可以发生可逆自组装,形成与病毒衣壳类似的结构,即病毒样颗粒(viruslike particle,VLP)。病毒粒子和VLP都可称为病毒纳米颗粒(virus-based nanoparticle,VNP),作为构建复合纳米结构和材料的组件。
VNP具有特定的形状和大小,人们可以根据目的选择合适的VNP。例如,正二十面体衣壳有丰富的内腔载物空间,适合作为各种分子或纳米颗粒的容器;螺旋对称的棒状及丝状衣壳则更适合用作一维纳米支架或模板。有些VNP在特定条件下可发生形变,如烟草花叶病毒(tobacco mosaic virus,TMV)[2]、噬菌体P22[3],这种环境响应机制赋予VNP更多的材料学属性和应用潜力。目前,包膜型VNP通常以活病毒的形式在细胞中产生,包膜型VNP在细胞外的解聚再重构尚未实现,一定程度上限制了包膜型VNP的修饰改造及应用。无包膜的VNP既可以在细胞中完成组装,也可以在细胞外组装,结构可操纵性更强,因而修饰改造策略及应用研究更丰富,因此本文涉及的VNP主要为无包膜VNP。总体而言,VNP或其组分能够在细胞工厂(宿主或重组表达系统)中大量高保真生产,因此它们是一类可规模化低成本生物合成的天然生物纳米材料。

图1 VNP举例(STMV、CCMV、SV40、HPV、P22结构模型从http://viperdb.scripps.edu/获得,M13结构摘自文献[9],TMV结构摘自文献[10])Fig.1 Schematic diagrams for virus-based nanoparticles(VNPs){The structure models of satellite tobacco mosaic virus(STMV),cowpea chlorotic mosaic virus(CCMV),simian virus 40(SV40),human papillomavirus(HPV),and P22 are obtained from http://viperdb.scripps.edu/.The models of M13 and TMV are reproduced from references[9]and[10],respectively}
自然界中来自不同物种的病毒,构成了丰富的天然病毒资源库,为发展病毒纳米材料奠定了基础。同时,受天然病毒衣壳组装的启发,近年来新兴的计算机辅助自组装蛋白纳米材料(特别是笼形蛋白结构)的理性设计,为发展病毒纳米技术提供了全新的材料基础和强大的改造能力[4-8]。
2 病毒纳米颗粒(VNP)的功能化策略
2.1 基因工程
编码病毒蛋白的核酸序列很容易获取。根据VNP结构信息进行理性结构设计[11],通过基因工程可以方便地将编码目的蛋白或肽段的核酸序列整合到病毒蛋白基因中,从而将外源功能模块引入VNP的特定位置(图2)。VNP的内腔、外表面以及亚基间的界面均可进行基因工程改造。外源基因引入位点常常位于VNP的氨基末端(N端)、羧基末端(C端)、柔性环结构,这样有利于保持VNP的组装特性和外源肽段的正确折叠。
VNP的末端通常暴露在其蛋白主体骨架的外面,末端修饰可降低外源片段与病毒蛋白主体之间的相互影响。因此末端对基因工程改造的容纳度高,是基因工程改造的首选位点。VNP纯化制备的最常用方法之一是蛋白亲和纯化。大量研究在VNP末端融合亲和纯化标签以实现亲和纯化。例如,在猴病毒40(simian virus 40,SV40)VP1的N端融合多聚组氨酸标签(histidine tag,Histag)[12],在人乳头瘤病毒(human papillomavirus,HPV)L1的N端融合谷胱甘肽巯基转移酶(glutathione S-transferase,GST)[13],在口蹄疫病毒(foot-and-mouth disease virus,FMDV)的N端融合小分子泛素样修饰蛋白(small ubiquitinlike modifier,SUMO)[14]等,以实现相关VNP的纯化。不同功能的肽段/蛋白也常常融合在VNP的末端以实现药物筛选、递送、治疗等目的,如:在M13噬菌体的衣壳蛋白pⅧ的N端融合多肽,用于筛选得到对靶标具有高亲和力的多肽药物[15];在噬菌体MS2 VNP的N端融合细胞穿膜肽[16];在P22 VNP的N端融合线粒体凋亡调控肽[17];SV40 VNP的N端融合动脉粥样硬化治疗肽[18]等。另外,VNP末端还可以引入功能肽段以促进或介导其他功能元件的装载,如在乙型肝炎病毒核心抗原(hepatitis B virus core antigen,HBc)的末端引入Histag以特异性包装Ni-NTA(Ni2+-nitrilotriacetic acid)修饰的氧化铁纳米颗粒(iron oxide nanoparticle,IONP)[19]、引入亲脂性多肽以实现包装多柔比星(阿霉素,doxorubicin,DOX)[20]、引入RNA结合蛋白以实现RNA的包装[21]。
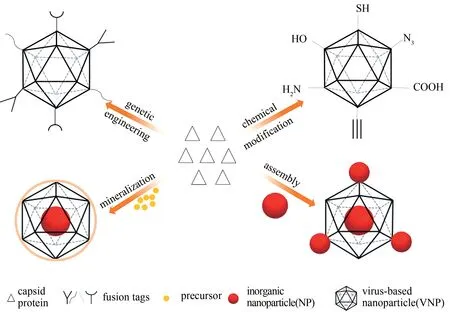
图2 通过基因工程、化学修饰、矿化和组装等策略对VNP进行功能化改造Fig.2 Functionalization of VNPs through different strategies,including genetic engineering,chemical modification,mineralization,and assembly
VNP内外表面的柔性环结构也是插入外源片段的常用位点。不同的VNP、不同位置的环结构对插入外源片段的兼容性不同。例如,SV40 VNP表面的BC环引入外源片段会破坏SV40 VNP的结构,而HI环和DE环上各发现一个可供外源片段插入的位点[22]。不同VNP表面柔性环对插入片段大小及结构的容纳度不同,需要更详细的探索,分子动力学模拟可以为此提供一定的参考。从整体上讲,VNP表面柔性环结构上可插入的外源片段涵盖短肽到分子量大于VNP亚基的蛋白质,如VNP可展示靶向整合素的RGD短肽[23]、靶向动脉粥样硬化斑块的短肽[18]、靶向人类表皮生长因子受体2(human epidermal growth factor receptor-2,HER2)的亲和配体[24-25]以及绿色荧光蛋白[26]等。
除了主要衣壳蛋白,次要衣壳蛋白或支架蛋白也可以用来融合外源多肽或蛋白。这些蛋白特异性的结合在主要衣壳蛋白上,对融合外源序列的容纳度往往更高。例如,M13噬菌体的次要衣壳蛋白pⅢ可融合展示单域抗体[27]或单链抗体[28],构建抗体的噬菌体展示文库;P22 VNP内部的支架蛋白SP上融合酶分子(乙醇脱氢酶D[29]、外切葡聚糖酶CelB[30]、Cas9[31-32]等),形成了多种VNP包装酶的纳米反应器;SV40次要衣壳蛋白VP2/VP3上融合荧光蛋白,形成了多种荧光成像方法[33-35]。人们可以通过对外源多肽序列的选择和设计,使之成为装载其他功能模块(如无机纳米颗粒)的锚定位点。
此外,对衣壳蛋白序列循环重排[36]和定向进化[37],能够大幅改变已有VNP的结构特征,可以从大量的改造文库中筛选获得具有新的组装特性[38]或者功能化位点[39]的VNP,为VNP的基因工程改造提供了新思路。
2.2 化学修饰
化学修饰可为VNP提供灵活多样的功能化方案。根据VNP的精确结构信息,能快速定位可用于修饰的氨基酸残基位点[40]。这些氨基酸有半胱氨酸[41]、赖氨酸[42]、天冬氨酸[43]、谷氨酸[44]、组氨酸[19]以及一些人工引入的非天然氨基酸(如带有叠氮基团的L-叠氮高苏氨酸和对氨基苯丙氨酸)[45]等等。通常选择暴露于VNP内或外表面以及位于亚基间界面的氨基酸残基进行生物缀合(bioconjugation),从而方便地把功能模块修饰在VNP上,常用的VNP化学修饰试剂及应用案例总结于表1。
2.3 矿化
生物矿化是在生物体特定的部位,在一定的物理化学条件和生物有机物质的控制或影响下,将溶液中的离子转变为固相矿物的过程。骨骼、牙齿和器官结石等都是通过生物矿化形成。受此启发,矿化也成为一种VNP功能化改造的方法[59]。VNP外表面和内腔均可矿化(图2)。一般策略是利用VNP上天然存在的无机物前体亲和位点或外源引入的无机物亲和肽/化合物与无机物前体发生相互作用,诱导无机物结晶成核,进而通过种子生长,形成均一有序的纳米颗粒。
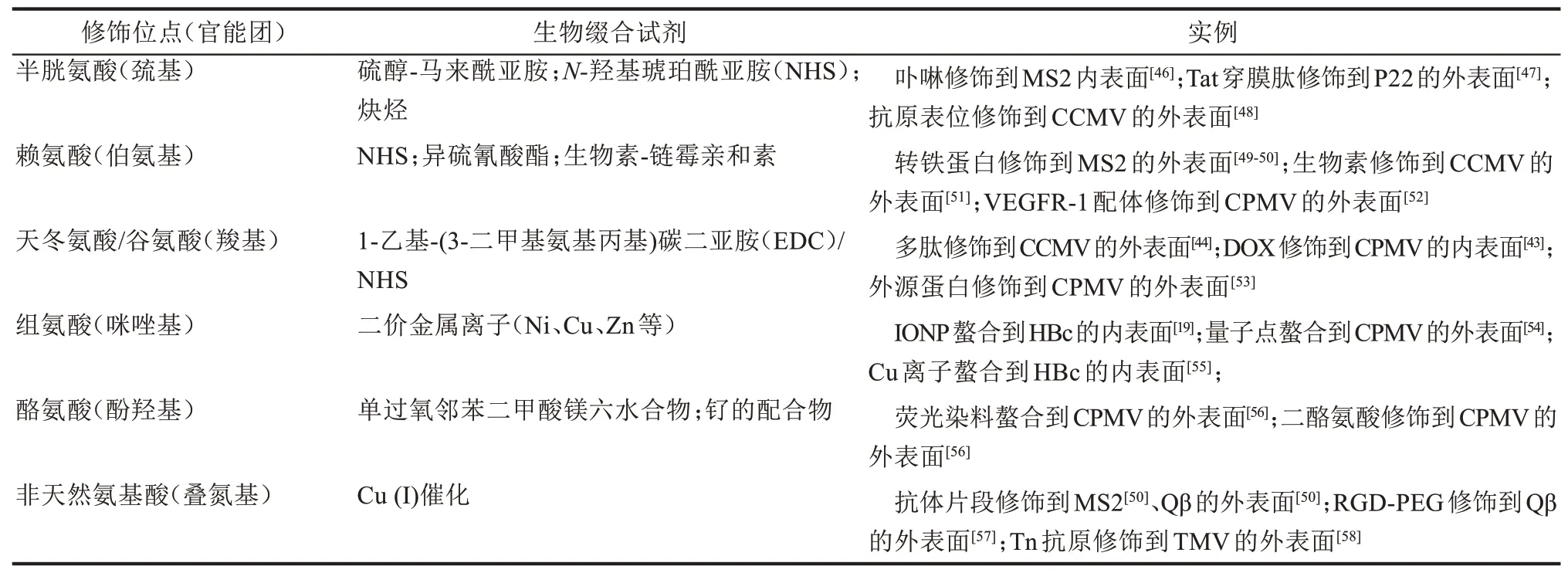
表1 VNP的生物缀合功能化实例[43-44,46-58]Tab.1 Functionalization of VNPs via bioconjugation
研究者在VNP外表面引入诱导无机物矿化的肽,成功将磷酸钙和硅沉积包裹到肠道病毒(enterovirus 71,EV71)[60-61]、口蹄疫病毒(footand-mouth disease virus,FMDV)[62]和登革病毒(dengue virus,DENV)疫苗的表面形成类似于“鸡蛋壳”的保护层,显著提高了疫苗的稳定性。Evans等[63]围绕豇豆花叶病毒(cowpea mosaic virus,CPMV)VNP外表面矿化开展了一系列工作。他们利用CPMV VNP表面对金属钯(Pd)前体的亲和性,在CPMV表面矿化形成Pd簇,以Pd簇作为后续矿化的成核位点,成功在CPMV表面生长了钴(Co)、镍(Ni)、铁(Fe)、铂(Pt)、Co-Pt和Ni-Fe等多种纳米材料。他们还通过在CPMV VNP表面覆盖一层阳离子聚合电解质,吸引负电荷的金(Au)前体,矿化了Au壳[64];通过在CPMV VNP外表面赖氨酸残基上引入负电荷的琥珀酰胺酸盐的方法吸引正电荷前体,矿化了Co及IONP[65]。
VNP的内腔尺寸,能够对矿化产物的大小进行限制,有助于形成均一的纳米颗粒。在VNP内腔矿化的一个策略是在VNP内腔引入短肽等无机物前体亲和基序。应用这种策略,人们实现了不同物理化学性质的纳米材料[如硫化铜(CuS)[55]、硫化镉(CdS)[66]、二氧化钛(TiO2)[67]、四氧化三铁(Fe3O4)[68]、Au[69]、银(Ag)[70]、Au@Ag[71]等]的矿化。另一种策略是通过溶液条件控制VNP的形变,实现前体物质在内腔的捕获。例如,利用豇豆褪绿斑驳病毒(cowpea chlorotic mosaic virus,CCMV)VNP pH响应的闸门机制,实现了前体物质在VNP内腔的捕获聚集,获得了IONP[72]、仲钨酸盐[73]和普鲁士蓝[74]纳米颗粒等。此外,对VNP内腔和外表面进行差异矿化能实现不同纳米材料性质的耦合[66]。
2.4 组装
在一定条件下,VNP可实现解聚-组装状态的相互转换,为VNP包装外源功能模块提供了可能性[75-76]。不同表面性质、组成和大小的蛋白质、核酸、药物、无机纳米材料都能被包装进VNP[77-80]。模拟病毒包装核酸的特性,赋予货物负电荷、增加货物与VNP内表面的静电相互作用,是VNP包装货物的常用策略之一。例如,表面带负电荷的金纳米颗粒(Au nanoparticle,AuNP)、量子点(quantum dot,QD)被高效包装到雀麦草花叶病毒(brome mosaic virus,BMV)VNP中[81-82]。同时,赋予衣壳蛋白与货物之间特异性亲和作用是诱导VNP包装货物的另一策略。例如,将红三叶草坏死花叶病毒(red clover necrotic mosaic virus,RCNMV)的组装起始RNA序列修饰到纳米颗粒表面,实现了RCNMV VNP包装直径3~15 nm的Au、四氧二铁酸钴(CoFe2O4)和硒化铬(CdSe)纳米颗粒[83-84]。除了病毒基因组包装信号序列,病毒内部蛋白、Ni-NTA与Histag、Spytag与Spycather、疏水相互作用等也被用来实现外源物质的选择性包装[19-20,85-87]。VNP的解聚-组装控制一般通过透析的方法,通过调节溶液的pH、盐离子强度、盐离子种类、氧化还原电势等使解聚的衣壳蛋白发生再组装,从而实现外源物质的包装。为了解决苛刻的解聚条件对货物稳定性和活性的损伤,本文作者团队[88]报道了一种利用蛋白质的表观临界组装浓度控制VNP组装包装外源物质的策略,大大扩展了VNP的载荷包装范围。
VNP外表面可以作为VNP与其他功能材料共组装的界面。通过组装可将功能材料密度可控地展示在VNP的外表面。例如,Li等通过调节SV40 VNP与AuNP之间的界面相互作用,成功构建了一系列AuNP与VNP的共组装体[89]。为了实现SV40 VNP表面纳米颗粒数量的更精确控制,他们建立了VNP的单功能化策略[90]。同时,通过调控VNP与其他材料的界面相互作用,还可使VNP形成多层级组装体。例如,利用CCMV与AuNP的静电相互作用构建三维二元超晶格[91],通过生物素与链霉素的亲和作用构建功能性CCMV三维杂合共组装体[92],利用静电驱动的层层组装效应驱动聚二甲基二烯丙基氯化铵与CPMV形成纳米薄膜[93]以及TMV层层组装形成超薄纤维膜等功能性材料[94]。
3 基于病毒组件的纳米材料的应用
3.1 生物医学成像
利用VNP进行生物医学成像,一方面是对疾病的定位和进展进行可视化诊断,旨在更早更灵敏的疾病检测;另一方面是追踪VNP纳米诊疗平台的细胞特异性摄取及活体行为(如活体分布、清除率、免疫原性等),对VNP纳米诊疗平台进评估和改进。VNP具有易于修饰、丰富的载物空间等特点,容易实现对信号分子的高密度装载以及与靶向元件联用,有利于高性能组织特异性成像,因而特别适合作为多功能成像探针。目前,VNP已用于光学成像、磁共振成像(magnetic resonance imaging,MRI)、正电子发射断层扫描(positron emission tomography,PET)等成像领域。
近年来,研究者进行了大量VNP用于光学成像的研究。例如,在SV40 VNP内腔蛋白VP2/VP3上利用基因工程引入荧光蛋白,实现了对SV40 VNP细胞摄取行为的追踪[33-35]。在BMV[95]、CCMV[96]上实现荧光染料高密度无猝灭标记,大幅提高了荧光成像的效率。QD是一种典型的荧光纳米探针,与荧光蛋白及荧光染料相比,具有亮度高、光稳定性好等特性,有利于提高成像的灵敏度及信噪比[97-98]。自2008年以来,人们进行了若干病毒偶联QD用于示踪的研究,为揭示病毒的感染机制提供了有价值的信息[99-101]。然而,在病毒外表面标记QD会对病毒-宿主相互作用造成潜在人为干扰,因而这方面的研究一直存在争议,而将QD包装在VNP内腔则能消除这种担忧。2009年,Li等[102]首次报道在VNP内腔包装可见光QD进行荧光示踪的研究,实现了对SV40 VNP细胞摄取行为的追踪,模拟了SV40细胞侵染的早期步骤[图3(a)]。2015年,进一步用VNP包装NIR-Ⅱ荧光(1000~1700 nm)硫化银(Ag2S)QD,实现了对SV40 VNP小鼠活体行为的实时动态追踪[图3(b)][103]。更有意思的是,Cui和Wang等[85-86,104-105]通过在病毒内腔的核酸或蛋白上标记QD,实现了活病毒包装QD,并成功地追踪了HIV-1的细胞侵染行为。荧光蛋白、有机荧光染料、QD等联合使用,可实现多重标记,为了解病毒的脱壳机制、VNP解聚及货物释放提供更多颇有价值的直观信息[34,104-108]。
VNP也可用以构建MRI探针。2005年,Douglas等[109]将MRI T1造影剂钆(Gd)与CCMV上的金属结合位点结合,使Gd-CCMV复合物的T1弛豫率比单独的Gd提高了10倍。随后,Kang等[110]利用P22 VNP高密度装载Gd,首次实现小鼠活体血管成像。2011年,Dragnea等[111]利用BMV VNP包装MRI T2加权成像造影剂IONP,利用MRI追踪了BMV在本氏烟草叶片细胞间的长距离迁移。以VNP作为骨架,将MRI造影剂与靶向元件整合在一起,有利于实现靶向成像。例如,在棒状噬菌体M13表面负载单分散磁性IONP,在M13末端引入SPARC糖蛋白靶向肽,实现了活体前列腺癌的特异性成像[112]。此外,也有来自不同机构的研究者利用MS2、P22、TMV、HBc等VNP装载Gd[110,113-117]、IONP[19]、锰(Mn)[118]、氙(Xe)[119-120]等造影剂,实现高信噪比MRI,显示了VNP在增强MRI方面的重要价值和应用潜力。
PET依赖于对放射性示踪剂的检测。Hooker等[41]在MS2 VNP内腔标记PET探针[18F]氟苯甲醛,实现了MS2大鼠活体行为追踪。Farkas等[121]在MS2内腔标记PET探针放射性铜(64Cu),实现了MS2及PEG-MS2荷瘤小鼠活体行为追踪,发现PEG修饰有利于减少MS2在脾中的富集,而MS2的肿瘤靶向一定程度上依赖于肿瘤微环境的高渗透长滞留效应(enhanced permeability and retention effect,EPR)效应。VNP上同时引入肿瘤靶向元件和PET造影剂,有利于实现VNP对肿瘤的主被动靶向和特异性成像[122-123]。VNP与放射性示踪剂优势互补,前者提供活体靶向性,后者作为PET成像探针,有灵敏度高、成像穿透深等优点,整合二者有利于实现活体水平高灵敏靶向成像和无创诊断。
3.2 生物传感
VNP具有高度规则有序的结构,它们尺寸、形态各异,三维结构能被精确表征到近原子分辨率,这使得人们能以近原子精准度对其进行操控以及化学特异性修饰,发展结构及表面性质高度可控的纳米骨架[124]。例如,Bruckman等[125]通过生物缀合得到苯胺低聚物(oligo aniline,OANI)修饰的TMV VNP(OANI-TMV),OANI-TMV进一步相互连接形成薄层,在感知到挥发性有机物(尤其是甲醇、乙醇)时发生形态变化从而导致电导性发生变化而检测出挥发性有机物。VNP具有易于改造的特点,修饰上不同配体便可成为不同的传感器。2001年,Dultsev等[126]通过将麦芽糖结合蛋白融合表达于M13 VNP表面,再用吸附了麦芽糖的石英晶体微天平对其进行检测,重组VNP与麦芽糖的特异性结合能通过电信号的变化检测出来,该研究为高敏感度、低消耗的病毒检测提供可能。2005年,Neufeld等[127]将报告酶(碱性磷酸酶)基因与M13噬菌体的基因融合表达于细菌中,该报告酶能与细菌周质空间的对氨基苯基磷酸酯反应,反应产物对氨基苯酚扩散出细菌并被电极氧化,他们通过检测电势的变化来判断酶是否表达,在具有合适的噬菌体和报告基因的情况下这样的系统可用于任何细菌的快速检测。
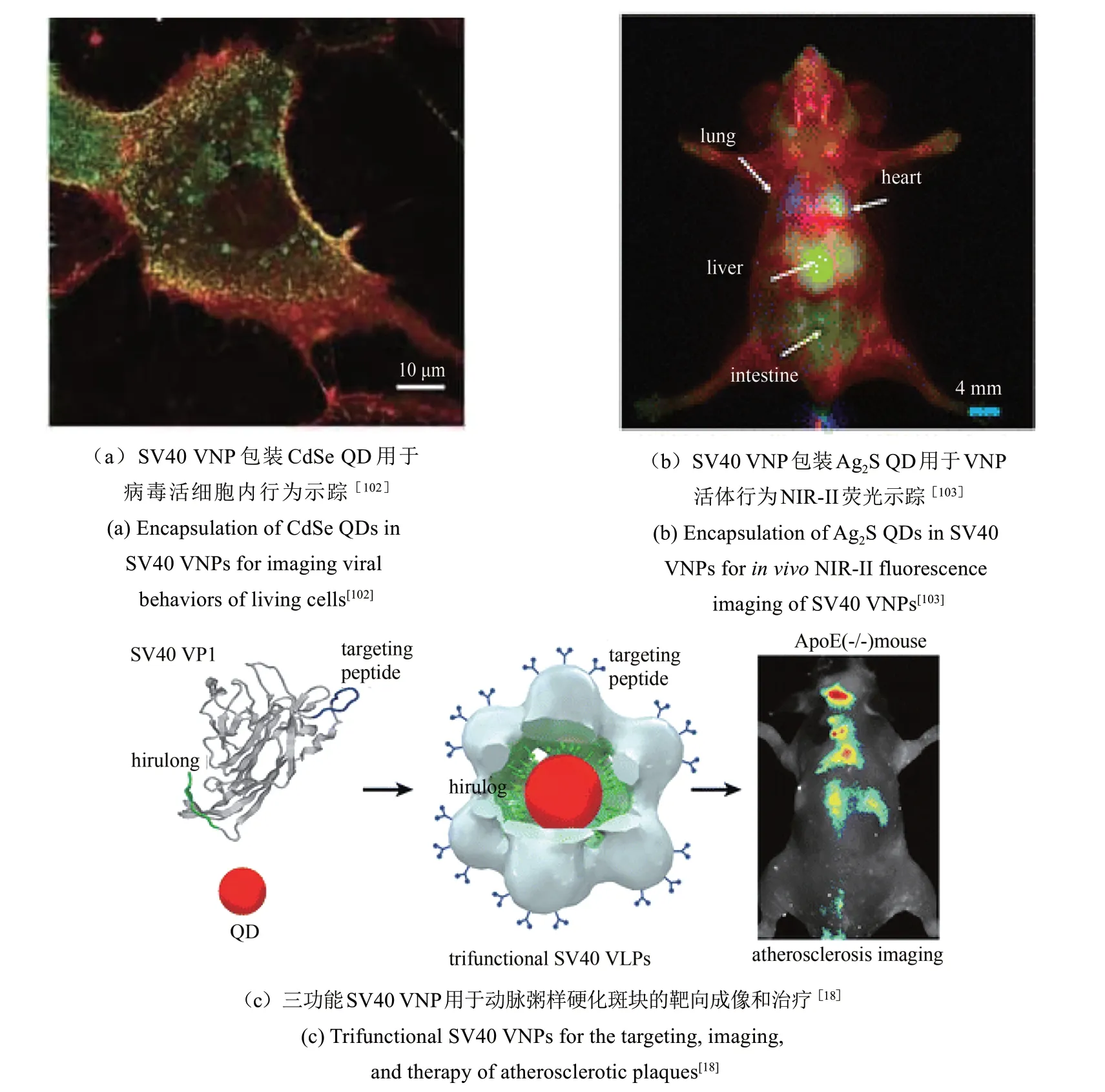
图3 VNP用于生物医学成像[18,102-103]Fig.3 VNPs for biomedical imaging[18,102-103]
3.3 核酸及药物递送
VNP一定程度上保留了跨细胞膜转运的特性,被广泛用于递送核酸、多肽、小分子、功能纳米材料等生物活性物质到细胞或活体中[128]。前文对VNP递送成像造影剂、抗原、酶分别进行了介绍,本节重点就VNP递送核酸及药物(多肽、小分子、功能纳米材料)方面的工作进行介绍。
使用VNP递送核酸的研究历史已超过半个世纪,产出了大量工作。其中一个典型代表是腺病毒基因载体[129-131]。利用噬菌体、植物病毒等也可实现基因递送,如在T7噬菌体上展示与病毒附着有关的乙型肝炎病毒(hepatitis B virus,HBV)包膜蛋白PreS1区,可使基因更有效地递送至人肝癌细胞HepG2[132]。除了基因,VNP还可递送mRNA[133-134]、小干扰RNA(small interfering RNA,siRNA)[49,135-136]和基因编辑RNA-酶复合物[137-138]等,发挥防止RNA降解、跨细胞递送、活体靶向的作用。基于VNP的核酸递送,更详细的介绍可参考Mah等的综述[131,139]。
VNP可通过基因工程进行功能化,便于靶向肽与治疗性多肽药物的整合,易于生物合成,因此是生物活性肽的良好递送平台[140]。本团队在P22衣壳蛋白上装载细胞毒性肽及组织蛋白酶B识别序列,实现细胞毒性肽在乳腺肿瘤细胞MDAMB-231中的环境响应性释放及治疗作用[17]。Sun等[18]在SV40 VNP上引入动脉粥样硬化斑块靶向肽及治疗肽,并在内腔包装荧光QD,从而构建集成像-靶向-治疗于一体的VNP平台,实现小鼠动脉粥样硬化的诊断、靶向及治疗[图3(c)]。这些研究表明,VNP对不同多肽的装载具有良好的兼容性,且易于与靶向、诊断等功能模块集成形成多功能诊疗纳米器件,具有重要临床应用潜力。
VNP还可用于递送小分子药物,与靶向元件联用,可增加治疗的特异性,减轻副作用。木槿褪绿环斑病毒(hibiscus chlorotic ringspot virus,HCRSV)同时包装聚丙烯酸和DOX以增加DOX的包封率,在VNP的外表面修饰叶酸以赋予VNP靶向肿瘤细胞的能力,大大增加了DOX的卵巢癌细胞摄取及细胞毒性[141]。TMV包装菲铂(phenanthriplatin)后,对小鼠三阴性乳腺癌肿瘤的治疗作用相比于传统抗癌药顺铂(cisplatin)或未包裹的菲铂均有明显提升[142]。Benhar等[143]在噬菌体M13和fd表面连接抗金黄色葡萄球菌的抗体和氯霉素,实现了特异性抗菌作用。Young等[144]将CCMV共价修饰钌配合物作为光敏剂,通过二极管照射产生活性氧,通过特异性抗体靶向杀灭金黄色葡萄球菌。Shan等[23]通过基因工程在HBc上引入RGD靶向肽,再通过解聚-再组装在RGD-HBc上加载吲哚菁绿(indocyanine green,ICG),从而实现U87MG肿瘤的靶向成像及光热/光动力治疗。
VNP内腔外表面具有丰富的载物空间,可通过界面设计使VNP与纳米材料共组装,因而VNP可以递送功能纳米材料。通过VNP递送纳米材料,往往可以实现多功能整合。腺病毒表面修饰1.3 nm光热转换AuNP,使光热治疗和基因治疗的联合成为可能[145]。Zhang等[146]用HBc包装四氧化三铁(Fe3O4)纳米颗粒,实现了对小鼠乳腺癌肿瘤的MRI及光热治疗。VNP的大小及形状影响其活体中的运输及清除行为,可根据应用需求选择合适的VNP[147]。生物活性物质递送是VNP的重要应用领域,受限于篇幅,更多信息可参考文献[1,45,79,148-150]。
3.4 疫苗与免疫调节剂
许多感染性疾病的病原是病毒。减毒活病毒、灭活病毒、VLP等可作为此类疾病的疫苗[151-153]。乙型肝炎及宫颈癌的疫苗就是基于病毒HBV及HPV的VLP[153]。利用化学/基因工程等修饰手段,VNP还可作为载体,用于呈递其他病毒的抗原、癌症抗原、免疫活性分子等,用于感染性疾病及癌症的免疫治疗[154-156]。VNP作为抗原载体,有利于提升抗原特异性中和抗体的滴度。基于HIV糖蛋白gp41的抗原表位连在AP205噬菌体上,可在小鼠体内产生高滴度的gp41特异性抗体[157]。抗原表位在载体上保持抗原的结构特性,有利于产生更好的免疫保护效果[158]。呼吸道合胞病毒(respiratory syncytial virus,RSV)的DS-Cav1三聚体疫苗结构与人工设计的类VNP I53-50上的三聚体成分完美对接。每个I53-50纳米球上可展示20个DS-Cav1三聚体,诱导的中和抗体应答比三聚体DS-Cav1高约10倍[图4(a)][159]。VNP表面多价高重复展示抗原有利于突破机体自身抗原免疫耐受。紧密连接蛋白18.2(claudin18.2,CLDN18.2)在多种肿瘤中高表达,在正常组织中的表达严格限制于胃黏膜中,是非常有潜力的肿瘤相关抗原。HBc表面展示CLDN18.2,可诱导机体产生高度特异性自身抗体并抑制肺转移瘤(CLDN18.2表达)的生长[160]。
VNP除递送抗原肽,还可递送编码抗原或免疫活性分子的mRNA。MS2包装前列腺酸磷酸酶(prostatic acid phosphatase,PAP)-集落刺激因子(granulocyte-macrophage colony-stimulating factor,GM-CSF)mRNA,可诱导PAP特异性抗体及T细胞应答,对小鼠前列腺肿瘤的生长具有抑制作用[133-134]。另外,某些VNP具有免疫调节剂的功能,可激活免疫系统,对和病毒无关的肿瘤具有治疗作用。2016年,Steinmetz团队[161]发现小鼠黑色素瘤、乳腺肿瘤、结肠肿瘤和卵巢肿瘤原位注射CPMV空壳,解除了肿瘤微环境的免疫抑制状态,使肿瘤的生长得到抑制甚至消除[图4(b)]。随后,研究发现马铃薯X病毒(potato X virus,PVX)[162]及木瓜花叶病毒(papaya mosaic virus,PapMV)[163]也表现出类似的免疫调节剂功能,具有抗肿瘤的效果。VNP装载化疗药物、光热治疗及光动力治疗活性物质的能力,有利于发展免疫治疗和其他疗法联合的抗肿瘤治疗方案。
3.5 组织工程
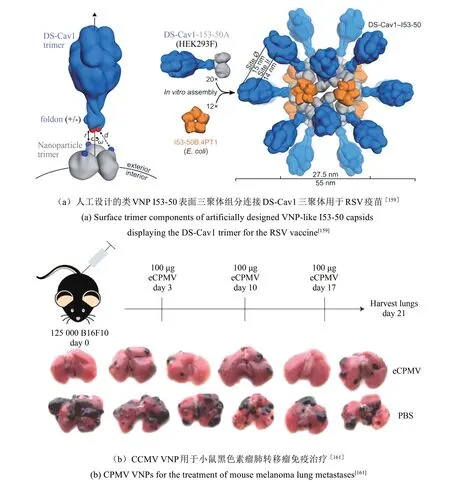
图4 VNP用作疫苗与免疫调节剂[159-161]Fig.4 VNPs as vaccines and immunomodulators[159-161]
在纳米尺度上控制仿生基质的结构和信号基序的装载对于组织再生材料的功能设计非常重要。VNP具有良好的生物相容性,已被应用于组织工程支架中,以指导细胞的生长、排列与分化。其中,棒状或丝状VNP可更好地模拟细胞外基质(extracellular matrix,ECM)的形态,因而被用于开发ECM,参与组织工程。如M13表面高密度修饰细胞黏附基序RGD或IKVAV,定向组装后形成液晶状纤维支架,能够控制神经祖细胞的增殖、分化、生长方向[164]。神经细胞在基于VNP的ECM上黏附、形态、增殖与分化,可通过调节VNP上修饰的细胞黏附基序、细胞因子等实现[165-166]。基于棒状或丝状VNP的ECM还被用于研究成骨分化。高密度展示RGD的M13填充入3D打印的生物陶瓷支架中,再植入间充质干细胞(mesenchymal stem cell,MSC)和骨缺损后,可诱导MSC的成骨分化和血管生成[167]。TMV掺入多孔硅藻酸盐凝胶中,以实现多孔凝胶支架的功能化,通过调节VNP上修饰的生物信号分子类型,从而控制支架中细胞的黏附与成骨分化[168]。基于VNP的组织工程支架实现活体应用,病毒的免疫原性及长期效应需要探究。将TMV-RGD掺入型多孔硅藻酸盐凝胶植入小鼠体内,并没表现出明显毒性并且VNP可被降解。噬菌体填充支架表现出的生物相容性和可降解性,使其作为生物医学支架材料更具可行性[169]。
3.6 纳米反应器
生物体系中存在大量的酶反应微区(microcompartment),如羧酶体、乙醇胺利用微区、丙二醇利用操纵子等,这些微区把特定的酶促反应限制在特定的区域中,实现分类管控,提高效率。受此启发,VNP受限的内腔可发展酶反应器[170]。酶可通过基因工程、非共价相互作用(如核酸适配体、静电相互作用)及共价相互作用(分选酶A介导)等方式包装在VNP内腔[78]。
VNP包装酶的第一个优势是可对酶形成保护,增强对热、pH、蛋白酶降解等条件变化的耐受性。肽酶E(peptidase E,PepE)包装在Qβ内腔,50℃30 min处理,仍保持100%活性,而游离PepE仅剩20%活性;Qβ包装PepE用蛋白酶K处理20 h还剩80%活性,而游离PepE用蛋白酶K处理10 min仅剩20%活性[171]。P22包装的磷酸三酯酶(phosphotriesterase)也具有更好的热稳定性和抗水解能力[172]。
VNP包装酶的第二个优势是可实现对受限空间中酶促反应特征的研究。通过解聚-再组装可实现每个CCMV包装一个辣根过氧化物酶(horse radish peroxidase,HRP),实现对单酶反应的研究,发现CCMV包装的HRP与游离HRP的反应特征不一样,这是由于CCMV对底物的进入及产物运出的影响所致[173]。CCMV包装的南极假单胞菌脂肪酶B(Pseudomonas antarcticalipase B,PalB)比游离酶有更高的活性,这可能是由于VNP内腔极高酶浓度导致的,说明了小体积对于有效的多酶级联催化的重要性[174]。P22内腔支架蛋白上基因工程同时引入CelB、ATP依赖性半乳糖激酶(galactokinase,GALK)、ADP依赖性葡萄糖激酶(glucokinase,GLUK),实现3种酶包装在P22内腔。CelB催化形成的两种产物恰好是GALK和GLUK两种酶的底物,在P22内腔形成代谢级联反应,GALK-GLUK-CelB-P22酶促转化效率大于GLUK-CelB-P22或CelB-P22/GLUK-P22混 合 物(图5)[175]。VNP包装级联酶,使一种酶的产物可以有效地作为下一种酶的底物,因而可提高反应效率。
VNP包装酶的第三个优势是可通过对VNP孔道的控制,实现对底物种类的筛选、对底物进入及产物输出速度的控制,从而达到调控酶反应的目的。如MS2通过解聚再组装对带有负电荷的碱性磷酸酶进行包装,通过改变VNP外壳上孔道的电荷性质能够控制酶底物的进入从而影响反应速度。在MS2孔道处引入负电荷抑制负电荷底物进入而抑制酶促反应,引入正电荷促进负电荷底物进入而促进酶促反应[176]。
VNP包装酶还存在许多挑战,如包装效率、酶活、VNP孔道对有些底物太小对有些酶太大等问题。通过摸索包装方式、控制包装酶的个数等有望解决包装效率及酶活问题。天然存在大量的VNP、通过进化修饰改造获得大量工程化VNP,人工设计肽/蛋白质组装而成的类VNP,为酶促反应提供了大量候选纳米酶促反应器,有望解决VNP孔道在酶促反应中的问题。
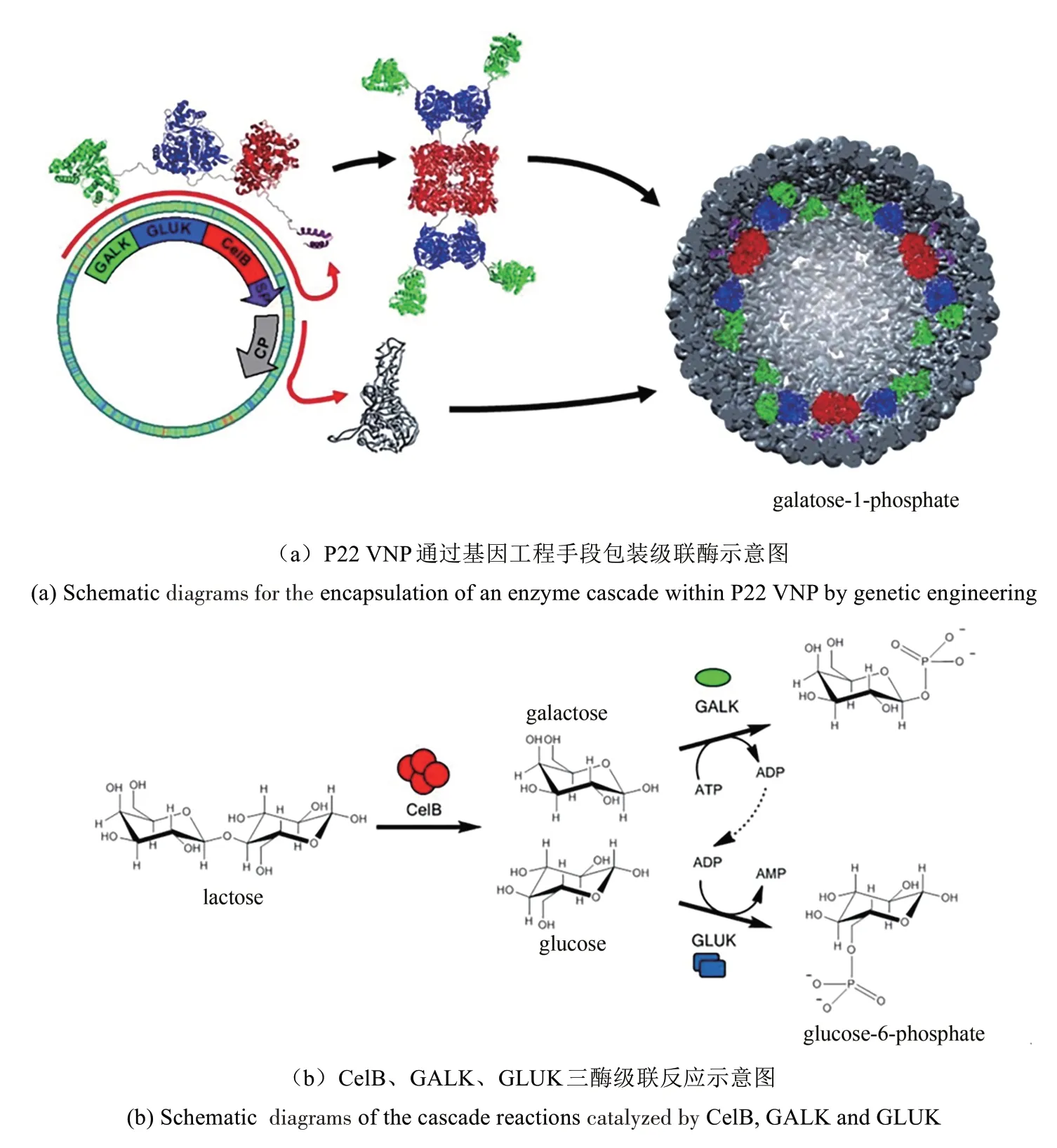
图5 基于VNP的酶纳米反应器[175]Fig.5 VNP-based nanoreactors for enzymatic reactions[175]
3.7 微电子元件
VNP蛋白骨架具有可编程、可控自组装的特性,它们能够作为模板对纳米材料(金属及其氧化物、半导体、石墨烯、碳纳米管等)进行精准吸附,实现不同物理性质的整合,形成精密微电子元件,可用于数据存储器、电池电极等。
2006年Tseng等[177]在TMV表面修饰铂纳米颗粒(Pt nanoparticle,PtNP),并在聚乙烯醇基质两极之间形成一个三明治结构,因此该纳米材料能束缚住电子且具有双稳态特性,在关闭状态具有低电导性,在打开状态具有高电导性。类似地,修饰半导体QD的CPMV VNP也具有电学双稳态性[178]。一些早期工作将氧化还原活性物质(如二茂铁和紫罗碱)定位于CPMV骨架上,发现这种多价氧化还原活性纳米颗粒具有多电子同时转移现象,说明位于CPMV骨架上的氧化还原单元之间是相互独立且在反应时不相互作用的[179-180],这种材料可用作多电子储存库或者多电子转移调节器。
病毒在微型电极材料的研发中也有独特应用价值。Nam等利用M13噬菌体作为模板生长氧化钴(Co2O3)纳米线,进而用作活性阳极材料,他们的工作表明病毒纳米材料网络结构可用于生产具有全电化学功能和更大容量的电极材料[181-182],当M13代替氧化锰(MnO)用作锂-氧(Li-O)电池正极时展示出了更好的性能[183]。另外一个值得注意的以M13为模板的电极材料的例子是利用氧化铱作负极材料。Nam等[184]先将AuNP化学吸附于M13表面,然后再进行二氧化铱(IrO2)的矿化,IrO2-Au杂化纳米线形成的多孔电极的电导性显著增强。此外,M13形成的液态晶体还具有压电效应特质。Lee等[185]在M13衣壳蛋白pVIII的N端引入谷氨酸残基以与带正电的C端产生更大的净偶极矩,使该纳米材料形成的薄膜的有效压电系数得到显著增强。
3.8 纳米光子学
许多纳米材料都具有可表征的光学性质,当它们距离足够近时,其光学性质的相互作用可能产生新的性质。2006年,Wang等[186]用芜菁黄花叶病毒(turnip yellow mosaic virus,TYMV)VNP建立起一种用于时间分辨免疫荧光分析的原型结构,采用荧光共振能量转移原理,可发展基于多价修饰VNP的传感器。2011年,Li等以SV40 VNP为骨架,构建了一系列内腔包装QD、外表面展示AuNP的杂化组装体(见2.4节),为定量研究AuNP和QD之间的光学相互作用现象提供了模型。实验结果与理论预测一致表明AuNP对QD荧光具有强烈的累积猝灭效应,而连接在VNP表面的AuNP之间仅存在微弱的表面等离子共振耦合效应(图6)[187]。而在另外一个实验体系中,包装在MS2 VNP内腔的AuNP与修饰在VNP外表面的荧光染料之间呈现距离依赖性荧光增强效应[188]。这些工作表明,得益于其精确可控的三维结构,VNP在多功能纳米材料的合成以及纳米光子学基础物理的探究中能够发挥重要作用。
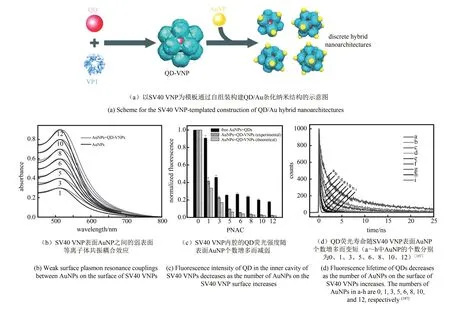
图6 VNP用于纳米光子学研究[187]Fig.6 VNPs for nanophotonics studies[187]
近年来,应用于能源技术开发的仿生光合成系统引起了人们的极大兴趣。与天然光合成系统类似,VNP能够提供亚纳米精度的发色团定位。棒状VNP具有螺旋对称性、周期性以及刚性,可作为构建仿生光捕获阵列的平台。2007年,Francis等[189]构建了以TMV纳米棒为模板的光捕获体系。衣壳蛋白和不同的发色团连接后,它们共组装形成了纳米盘或者纳米棒结构,并表现出光转移活性。在广谱性光收集三发色团系统中,总效率超过90%。在双发色团系统的皮秒时间分辨荧光实验中,187 ps内Oregon Green 488对Alexa Fluor 594的能量转移效率为36%[190]。为了构建几何学上接近自然光合成系统的光捕获结构,Dedeo等[191]通过突变构建了蛋白质N端或者C端位于纳米盘或棒中央的组装体,为在内孔修饰发色团提供了便利。发色团的朝向会影响光捕获过程中能量的转移,科学家通过引入一定数量的缺失突变来评估这种效应,结果表明TMV纳米棒状系统相较于盘状系统具有更高的光捕获效率,原因在于供体到供体转移途径的延长[192]。这项工作为优化人工光捕获纳米结构提供了思路。此外,Majima等[193]以TMV为模板,以锌(Zn)配位卟啉及自由碱基卟啉为供体和受体的实验表明能量转移效率还和骨架中基团的距离相关。
4 挑战与展望
综上所述,VNP已经成为自组装合成纳米材料的重要组件。VNP的诸多特性(包括可控的自组装、精确的三维结构和空间可寻址性、高度均质性、天然的生物相容性、易于基因工程/化学修饰、可规模化生物合成,等等),使之成为广受关注的基础生物纳米材料。本文详细介绍了VNP的功能化策略及其在生物医学成像、生物活性分子递送、免疫调节剂与疫苗、组织工程、噬菌体展示、纳米反应器、生物传感、微电子元件、纳米光子学等各个领域的应用。VNP在不同领域应用的成熟度,差异很大,意味着病毒纳米材料的发展还有很大空间,还有很多亟待解决的难题。VNP向临床转化的一个长期以来的挑战是如何精准地调控VNP的免疫反应及清除。VNP的免疫原性是把“双刃剑”,在免疫治疗时是有利的,而在活体成像和药物递送时却是不利的。VNP表面PEG修饰、包被血清蛋白等“免疫隐身”策略,在一定程度上屏蔽了VNP的免疫原性,有利于实现VNP靶向递送,但效果仍不甚理想。随着成像手段及成像探针的发展,更准确深入地探索VNP的活体行为与命运,有助于指导VNP设计、推动相关技术的临床转化。VNP在组织工程、能源、纳米反应器、纳米光子学等领域的应用,对其结构的稳定性、动态控制、物质交换控制等都有更高的要求。随着对病毒结构和组装的更深入理解、自组装蛋白质材料理性设计技术的成熟以及高通量合成生物学手段的发展,人们对VNP的操控和制造能力有望获得质的提升,主要的应用瓶颈将获得突破,也将打开病毒纳米材料广泛应用与转化的新局面。
猜你喜欢
学与玩(2022年8期)2022-10-31
河南科技(2022年8期)2022-05-31
解放军医学院学报(2022年1期)2022-04-02
科学导报(2021年82期)2021-12-17
锦绣·中旬刊(2021年10期)2021-08-23
化工管理(2021年7期)2021-05-13
理化检验(物理分册)(2021年3期)2021-03-23
建材发展导向(2020年16期)2020-09-25
汽车维护与修理(2019年14期)2019-08-08
科技创新与应用(2019年8期)2019-06-27
