
不同林龄尾巨桉人工林根系-土壤C、N、P生态化学计量特征
2020-10-10 03:08竹万宽许宇星王志超杜阿朋
桉树科技 2020年3期
竹万宽,许宇星,王志超,杜阿朋
(国家林业和草原局桉树研究开发中心,广东 湛江 524022)
在陆地生态系统中,植物作为生产者将光合产物以根系分泌物、溢泌物和脱落物的形式释放至土壤[1]。土壤、根系之间通过物质和信息的交换形成复杂的交互作用关系。土壤C、N、P是森林土壤养分的重要组成部分,也是植物生长发育必需养分元素,其含量变化和循环特征对植物生长和生理机能具有重要的调控作用[2-3]。因此,研究土壤-根系C、N、P的含量变异特征及其动态分布对于揭示系统养分循环特征具有重要意义。生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是C、N、P)平衡的科学,以及元素平衡对生态交互作用的一种理论[4-6]。作为一个新兴的生态学研究领域和生态学、生物化学及土壤化学研究新方向[4,7],被越来越多地应用到评估C、N、P等养分元素在土壤、植物中的循环特征及相互作用机制的研究当中。生态化学计量学已成为连接分子、细胞、种群、群落和生态系统等不同尺度生物学的新工具[8],在消费者驱动的养分循环、限制性养分元素判别以及全球碳氮磷循环等方面发挥重要作用[9]。
内稳性理论是指生物有机体在面对外界环境中元素可利用性变化时保持自身化学计量特征相对稳定的能力[10],是生态化学计量学的核心概念之一[11]。植物体对生态化学计量内稳性的调节功能反映了植物的生理和生化分配作用对其外部环境的响应机制,并通过根系对营养元素吸收和释放进行调节[12]。
桉树(Eucalyptus)是世界三大速生丰产树种之一。因具有良好的经济、生态和社会效益得到广泛引种和种植,截至2018年底我国桉树人工林面积已达5.46 × 106hm2[13]。非科学的经营措施(如超短轮伐期、连栽等)已造成地力衰退等问题[14],成为制约人工林生产力的主要原因[15]。因此,研究桉树人工林生态系统生态化学计量学特征可以为桉树可持续经营提供依据。目前,对桉树人工林生态化学计量学研究主要集中在器官、凋落物和土壤等尺度[13,16-17],尚未见报道土壤-根系生态化学计量特征。本研究以雷州半岛不同林龄尾巨桉(E.urophylla×E. grandis)人工林为研究对象,以空间代替时间,采集不同林龄桉树人工林土壤、根系样品,测定并分析其 C、N、P含量差异特征及元素化学计量比格局及内稳性特征,以期为桉树高效可持续经营养分管理提供数据支持。
1 研究区概况
本研究样地位于雷州半岛北部广东湛江桉树人工 林 生 态 系 统 定 位 研 究 站 (21°15′53″N ,110°05′39″E)。地处北热带湿润大区雷琼区北缘,为海洋性季风气候。最高海拔220.8 m,最低海拔80 m。年平均气温23.1℃,最热月(7月)平均气温28.8℃,最冷月(1月)平均气温 15.6℃,≥10℃的活动积温8 373℃。年平均降水量 1 567 mm,年相对湿度80.4%,年均日照时数1 937 h。试验地土壤类型为玄武岩发育的砖红壤,土壤肥力中等,土层厚度达84 cm以上。林下植被种类较为丰富,植被组成以草本为主,如南美蟛蜞菊(Wedelia trilobata)、牛筋草(Eleusine indica)、马唐(Digitaria sanguinalis)、山管兰(Dianella ensifolia)、芒萁(Dicranopteris dichotoma)等。灌木有五色梅(Common lantana)、白背叶(Mallotus apelta)、鹅掌柴(Schefflera octophylla)、盐肤木(Rhus chinensis)等。
2 研究方法
2.1 样地选取
2017年7月以空间换时间的方法在研究区选取立地条件相似、不同生长阶段2、6和9 a生尾巨桉人工林为研究对象。3个林龄尾巨桉人工林初始造林密度均为1 666株·hm-2,于造林前穴施桉树专用基肥。每个林龄随机设置3个20 m × 20 m样地,样地调查因子包括林分保留密度、树高、胸径、叶面积指数、林下植被等。样地信息概况见表1。
2.2 样品采集与处理
于2018年6月对各样地进行每木检尺,测量胸径和树高,并统计株数。按照胸径序列,将林木划分为基数基本相等的大、中、小3个径级,每个径级选择1株标准木。将标准木在地径处伐倒后采用全挖法并按照细根(≤2 mm)、中根(2 ~ 5 mm)和粗根(≥5 mm)规格采集活根样品,3株标准木获得的同规格混合成1份样品,使用流水冲洗根表泥土后装入牛皮纸袋,标记带回实验室。植物样品在65℃烘箱中烘干至恒质量,经粉碎过100目筛,用于测定C、N、P含量。在各样地内,采用“S”形路线设置5个样点,分别挖取土壤剖面,环刀法(100 cm3)取样用于测定土壤物理性质,同时采集0 ~ 20 cm,20 ~ 40 cm和40 ~ 60 cm层次土壤,同层土壤混合均匀装袋标记后置于含冰袋的保温箱中,带回实验室后一部分进行风干,研磨,过筛,用于测定土壤C、N、P含量。
林木根系和土壤有机C采用高温外热重铬酸钾氧化-容量法测定;林木根系和土壤全 N采用凯氏定氮法测定;林木根系全P采用硫酸-双氧水消煮,钒钼黄比色法测定,土壤全P采用氢氧化钠熔融,钼锑抗比色法测定。
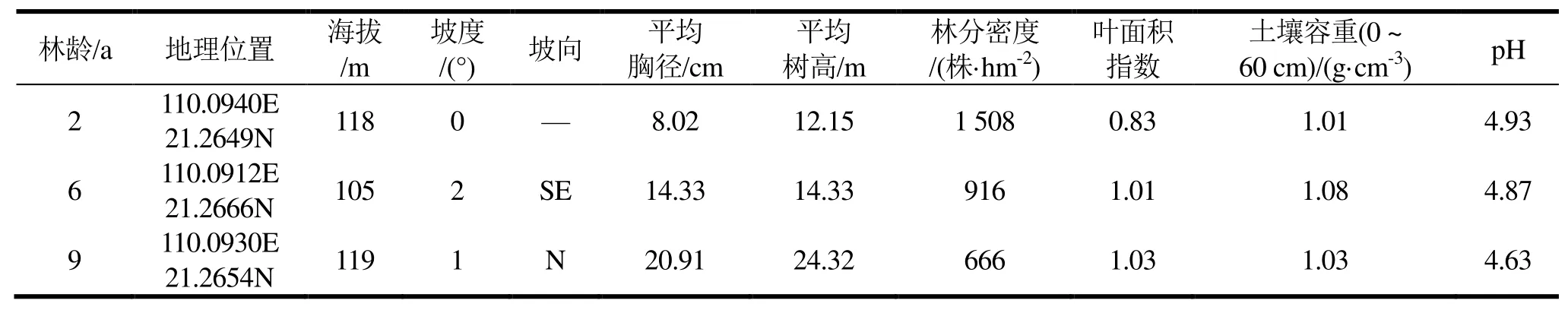
表1 样地基本概况
2.3 数据处理
植物根系和土壤的C、N、P化学计量比采用质量比表示[12]。生态化学计量内稳性指数(H)的计算采用以下模型[4]:

经对数转换后为:
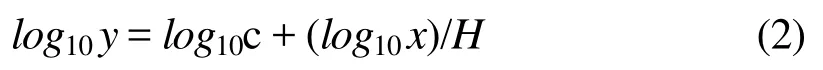
式中:y表示有机体中的元素含量或元素化学计量比值,x表示对应的环境供应营养元素含量或元素化学计量比值,c为常数。
根据相前研究将内稳性划分为以下类型[11,18-19],若拟合方程未达到显著水平(α=0.05)或1/H≤0,为严格内稳态;0<1/H<0.25,为内稳态;0.25<1/H<0.5,为弱内稳态;0.5<1/H<0.75,为弱敏感型;1/H>0.75,为敏感型。
试验数据采用 EXCEL 2016进行初步统计整理,使用SPSS 19.0软件对不同林龄桉树林根系、土壤C、N、P含量及C:N、C:P和N:P进行方差分析和显著性检验(P=0.05)。用Pearson法分析林木根系、土壤C、N、P含量及C:N、C:P和N:P相关性,根系与土壤C、N、P含量及化学计量比的相关性采用粗根、中根和细根的平均值与0 ~ 60 cm土层平均值进行处理分析。图形制作使用SigmaPlot 14.0软件。
3 结果与分析
3.1 不同林龄尾巨桉林根系、土壤C、N、P含量
如图1所示,2、6、9 a生尾巨桉林根系有机C含量分别为 470.60 ~ 503.40、480.30 ~ 482.70和458.80 ~ 512.90 g·kg-1,全N含量分别为 3.94 ~ 5.01、2.49 ~ 2.88 和 3.81 ~ 4.52 g·kg-1,全P 含量分别为0.29 ~ 0.32、0.31 ~ 0.41 和 0.20 ~ 0.33 g·kg-1。2 a和6 a不同径级根系有机C、全N和全P含量均无显著差异(P>0.05),9 a不同径级根系有机C含量表现为粗根>中根>细根,全N含量无显著差异(P>0.05),全 P含量为中根和细根显著高于粗根(P<0.05)。根系有机C和全N平均含量均表现为随林龄增大呈先减后增趋势,全P含量为9 a最小。

图1 不同林龄尾巨桉林根系C、N、P含量
由图2可知,2、6和 9 a生尾巨桉林土壤有机C 含量分别为10.60 ~ 25.16、11.04 ~ 18.59和16.05 ~25.72 g·kg-1,全N含量分别为0.93 ~ 1.88、1.09 ~ 1.36和 1.20 ~ 1.71 g·kg-1,全 P 含量分别为 0.69 ~ 0.74、0.76 ~ 0.79 和 0.85 ~ 0.92 g·kg-1。3 个林分土壤有机C含量均随土层加深递减,土层间差异显著(P<0.05)。0 ~ 20和20 ~ 40 cm土层有机C含量均为2 a和9 a显著高于6 a生(P<0.05),2 a和9 a生间无显著差异(P>0.05)。40 ~ 60 cm土层有机C含量为9 a显著高于2 a和6 a(P<0.05),2 a和6 a间无显著差异(P>0.05)。

图2 不同林龄尾巨桉林土壤C、N、P含量
3.2 不同林龄尾巨桉林根系和土壤C、N、P化学计量比
3个林龄根系C:N均表现为粗根>中根>细根,但除2 a粗根和细根差异显著外其他均无显著性差异(表2)。粗根、中根和细根C:N随林龄增大呈先增后减趋势,粗根和中根表现为6 a显著高于2 a(P<0.05),两者与9 a无显著性差异(P>0.05)。细根为6 a显著高于 2 a和 9 a(P<0.05),后两者间无显著差异(P>0.05)。根系C:N均值为6 a显著高于2 a(P<0.05)。3个林龄根系C:P除9 a粗根显著高于中根和细根外(P<0.05),其他不同径级根系间无显著性差异(P>0.05)。粗根、中根和细根C:P随林龄增大呈先减后增趋势,粗根表现为9 a显著高于2 a和6 a(P<0.05),其他林龄间无显著性差异(P>0.05)。根系 C:P均值为9 a显著高于2 a和6 a(P<0.05)。3个林龄根系N:P除2 a粗根显著低于中根和细根(P<0.05)外,其他不同径级根系间无显著性差异(P>0.05)。粗根、中根和细根N:P随林龄增大呈先减后增趋势,粗根为9 a显著高于6 a,中根为2 a>9 a>6 a,细根为2 a和9 a显著高于6 a(P<0.05)。根系N:P均值为9 a显著高于6 a(P<0.05)。
3个林分土壤C:N土层深度变化规律不同(表3),2 a为20 ~ 40 cm显著高于40 ~ 60 cm(P<0.05),两者与0 ~ 20 cm均无显著性差异(P>0.05);6 a为0 ~ 20 cm显著高于40 ~ 60 cm(P<0.05),两者与20 ~ 40 cm均无显著差异(P>0.05);9 a为0 ~ 20 cm显著高于20 ~ 40 cm(P<0.05),两者与40 ~ 60 cm均无显著性差异(P>0.05)。0 ~ 20 cm土层3个林龄C:N无显著差异;20 ~ 40 cm土层为2 a显著高于6 a和9 a(P<0.05);40 ~ 60 cm土层为9 a显著高于2 a和6 a(P<0.05)。土壤C:P土层深度变化规律一致,均表现为0 ~20 cm>20 ~ 40 cm>40 ~ 60 cm,两两之间差异显著(P<0.05)。0 ~ 20 cm和20 ~ 40 cm土层C:P为2 a>9 a>6 a,两两之间差异显著(P<0.05);40 ~ 60 cm土层为9 a显著高于其他林龄(P<0.05)。土壤N:P土层深度变化规律不同,2 a为0 ~ 20 cm显著高于其他土层(P<0.05),6 a各土层间无显著差异(P>0.05),9 a为0~ 20、20 ~ 40 cm显著高于40 ~ 60 cm(P<0.05)。除0~ 20 cm土层N:P显著高于6 a和9 a(P<0.05)外,其他土层的林龄之间无显著差异(P>0.05)。0 ~ 60 cm土壤C:N、C:P和 N:P均值随林龄增大呈先减后增趋势,但均无显著显著性差异(P>0.05)。

表2 不同林龄尾巨桉林根系C、N、P化学计量比

表3 不同林龄尾巨桉林土壤C、N、P化学计量比
3.3 尾巨桉林根系-土壤C、N、P含量相关性分析
由表4可知,根系有机C含量与根系全N、土壤有机 C 含量显著(P<0.05)或极显著(P<0.01)正相关;根系全N含量与土壤C、N、P含量均未达到显著水平;根系全P含量与土壤有机C、全P含量显著(P<0.05)或极显著(P<0.01)负相关。土壤有机C含量与土壤全N、全P含量显著(P<0.05)或极显著(P<0.01)正相关。表明根系-土壤C、N、P含量之间相互影响,影响程度存在较大差异。

表4 尾巨桉林根系-土壤C、N、P含量相关性
3.4 尾巨桉林根系-土壤C、N、P化学计量比相关性分析
根系C:N与根系N:P、土壤C:N及土壤C:P存在显著或极显著的负相关关系(表5)。根系C:P仅与其 N:P存在极显著正相关关系。根系 N:P与土壤C:N、C:P存在显著或极显著的正相关关系。土壤C:N与C:P极显著正相关。土壤C:P与N:P极显著正相关。

表5 尾巨桉林根系-土壤 C、N、P比值相关性
3.5 不同林龄尾巨桉林根系C、N、P化学计量比内稳性特征
由表6可知,2 a和6 a根系C、N、P及其化学计量比均为严格内稳态(除 6 a C:P内稳指数为1.795,为弱敏感型外),9 a根系C、N、P及其化学计量比均达到内稳态(除C:P内稳性指数为0.694,为敏感型外)。表明随着土壤营养环境的改变,6 a和9 a根系C:P呈部分比例变化。
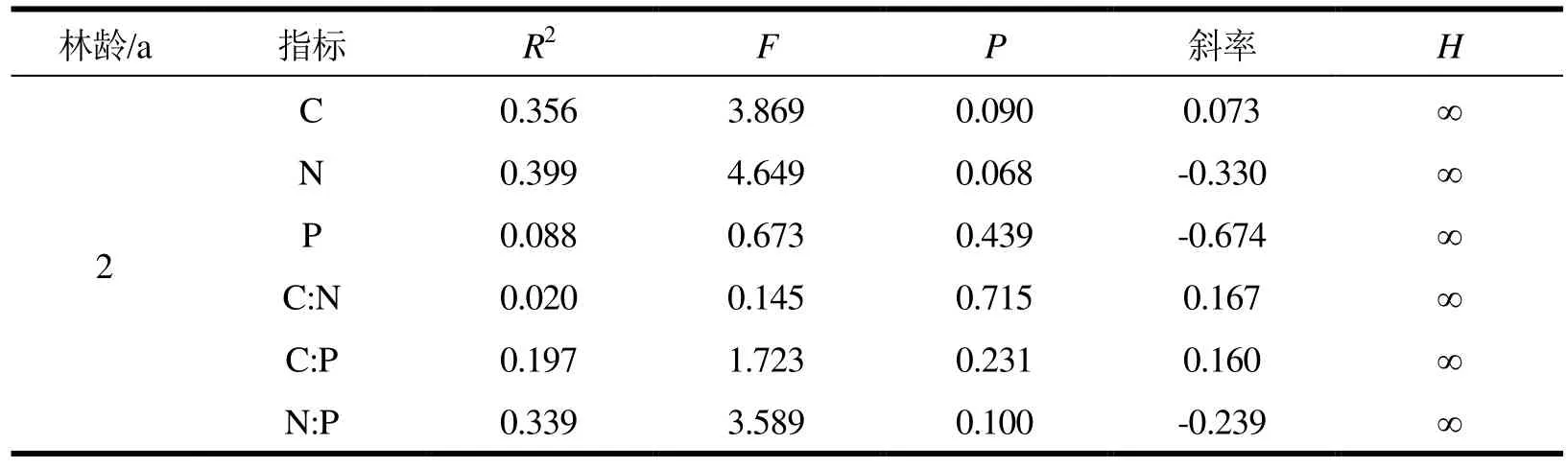
表6 不同林龄尾巨桉林根系C、N、P比值内稳性指数

续表6
4 讨论
4.1 不同林龄尾巨桉人工林根系-土壤C、N、P含量特征
根系作为森林生态系统中生物能存在的一种形式,积累和储存了大量的养分元素,其凋落分解后将养分归还土壤,直接参与森林生态系统生物循环过程[20]。本研究不同径级根系有机C和全P含量仅在9 a时表现出显著差异性,说明林龄增大对有机C和P在不同径级根系的分配影响显著,对N的分配影响不明显。这是因为随着林龄的增大,不同径级根系生物量占比及阳离子交换能力存在差异[20]。
土壤养分元素是植物体内养分元素的主要来源,林木的养分状况也反映了土壤的肥力状况[21]。研究表明,桉树造林导致热带土壤肥力下降及土壤酸化,连栽对土壤化学性质负面影响明显[22-23]。但也有研究发现桉树取代马尾松(Pinus massoniana)后土壤碱解N、速效P及pH有所提高[24],巨尾桉(E. grandis×E.urophylla)林经营18年后土壤供肥潜力提高[25],14年生桉树林土壤有机质含量提高了0.50% ~ 0.75%[26]。本研究结果表明,在2 a和6 a阶段,土壤有机C和全N含量出现降低趋势,9 a阶段得到恢复,全P含量随林龄逐渐增大。这可能是因为在9 a阶段,林木生长速率减缓,对养分的吸收利用率降低,同时凋落物不断积累和分解,养分归还土壤高于消耗。
根系与土壤直接接触,是植物生命活动的重要场所[27]。在植被-土壤系统中,土壤养分可直接影响植物对养分的吸收和利用,改变植物养分含量、化学计量比及其生态策略[28]。本研究结果发现,根系有机C主要受土壤有机C影响,根系全P含量主要受土壤有机C和土壤全P的共同影响。根系C、N、P和土壤养分元素关系密切,受土壤养分状况的影响明显。
4.2 不同林龄尾巨桉人工林根系-土壤C、N、P化学计量比特征
植物C:N和C:P可以反映N、P利用效率和植物生长速率,N:P则反映植物受N或P的限制情况。本研究中,不同林龄尾巨桉根系C:N和C:P均远高于高于全球植物根系平均水平(48.25和1158.00)[29],表明研究区尾巨桉人工林具有较高的元素利用效率。N:P均低于全球平均水平(24.00)[30],2 a和9 a均高于全国平均水平(13.5),6 a则远低于全国平均水平[31],说明随着林分的不断演替,养分限制状况的决定元素可能会出现差异。
土壤C:N:P是评价土壤质量的重要指标[31]。本研究中,土壤C:N为11.70 ~ 13.63,较为接近全球土壤 C:N平均水平(14.31)[32]和中国湿润温带土壤C:N为10~12,远低于热带、亚热带地区红、黄壤(20)[33],说明研究区土壤硝酸盐淋溶风险高[34]。C:P和N:P分别为19.25 ~ 24.87和1.66 ~ 1.84,远低于全球平均水平(186和13)[33]和森林表层土壤(81.9和6.6)[35],而研究区全P含量处于较高水平,说明研究区可能存在N素有效性低的现象。
相关性分析表明,根系N:P与C:N、C:P存在极显著相关性,说明根系对环境N、P的应对策略会影响根对碳的固存。根系C:N与土壤C:N、C:P均为显著负相关,说明了植物对营养元素的需求依赖于土壤中的营养元素,土壤有机C与根系有机C、全P的极显著相关性也证明了这一点。
4.3 不同林龄尾巨桉人工林根系内稳性特征
生态化学计量内稳性能够较好地反映生物对环境变化的生理和生化得适应,生物体内稳性的强弱与物种的生态策略和适应性有关[36]。2 a和6 a根系C、N、P、C:N、C:P、N:P均具有内稳性,即具有保持其自身化学元素组成相对稳定的能力。9 a根系C:P属于敏感型,说明二者受土壤环境的影响较大,具有一定的可塑性。这是由于整地、造林及施肥等人为措施大幅度改变了土壤环境,而根系会采取一些生理策略应对这种养分元素的剧烈变化,从而保持自身化学元素的相对稳定。随着时间的推移,植物在不同的生长发育阶段会具有不同的内稳性特征,这反映了植物在不同养分条件下对其生长过程中地上与地下器官养分分配的权衡,这也是其逐渐对环境生理策略适应的结果[37]。
猜你喜欢
防护林科技(2022年5期)2022-08-22
建材发展导向(2022年10期)2022-07-28
中国土壤与肥料(2022年4期)2022-06-14
落叶果树(2021年6期)2021-02-12
防护林科技(2020年10期)2020-12-22
防护林科技(2020年9期)2020-11-09
绿色科技(2020年9期)2020-07-17
文苑(2020年12期)2020-04-13
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
