
苦荞遗传群体农艺性状分析
2020-09-18 09:13:04陈博宁章洁琼崔娅松霍冬敖陈庆富
耕作与栽培 2020年3期
陈博宁,章洁琼,崔娅松,霍冬敖,陈庆富
(1.贵州师范大学,荞麦产业技术研究中心,贵阳 550025;2.贵州省农作物技术推广总站,贵阳 550001)
苦荞是双子叶蓼科(Polygonaceae)苦荞属(FagopyrumMill)植物。中国是苦荞的起源地和多样性中心,有着丰富的野生资源和栽培品系。到目前为止,我国已收集的荞麦资源有2 800余份[1]。苦荞因其独特的抗旱耐瘠、营养均衡、生育期可塑性强等特性,已成为中西部生态脆弱地区重要的粮经作物[2-3]。荞麦是我国传统出口粮食之一,作为一种药食同源的粮食作物,含有丰富的氨基酸、矿物质及微量元素,并富含生物黄酮类活性物质,具有降三高、防癌抗癌等多种保健功能,有着极高的营养价值,在国际市场上一直处于供不应求的地位[4]。随着工业化进程加速,我国城镇化率亦有了较大提升。然而,在城镇化迅猛发展的过程中,土地资源无节制开发,对国家粮食安全和生态安全造成了威胁。针对耕地资源供给的刚性制约,如何在有限的耕地上获得更多的粮食产出成为育种家们亟需破解的难题。同时,苦荞分子育种基础研究滞后,育种效率低,难以满足市场需求。基于构建的遗传连锁图谱,苦荞产量相关性状QTL定位研究取得了一定的进展。研究结果表明,在单株粒重和主茎分枝数方面各检测到一个主效的QTL位点qYPP 1和qBN 3,置信区间分别为19.4 cM和22.9 cM[5-6]。主效QTL位点的发掘对于解析产量性状形成的遗传机理奠定了坚实基础。因此,利用遗传分离群体开展农艺性状解析,筛选综合性状优异苦荞资源对苦荞品种改良有着重要意义。
本研究以“黑米荞×不落粒野苦荞”构建的F2代群体197个单株为材料,分析了株高、主茎粗、主茎分枝数、主茎节、百粒重的分离情况,比较了不同性状间的相关性,筛选出一批具备高产潜质的良好株系,为苦荞品种选育提供理论支撑。
1 材料与方法
1.1 实验材料
供试材料由贵州师范大学荞麦产业技术研究中心提供。母本材料为黑米荞:早熟、中秆、小粒、黑色、易脱壳、分枝多;父本材料为花药迟开裂型黑米荞:花药迟开裂、晚熟、分枝少、黑色、中秆。
1.2 亲本和遗传群体栽培
供试亲本材料与其衍生的F2分离群体于2016年8月播种于贵州省黔南州长顺县广顺镇核子村基地。种植于同一平整地块,常规田间管理,行距40 cm、行长1.8 m,株距6 cm。
1.3 性状考察
株高:塔尺测量植株主茎顶端至基部的长度(cm)。
主茎粗:游标卡尺测量植株主茎粗细(mm)。
主茎分枝数:植株主茎着生的有效分枝数(个)。
主茎节:植株主茎分节数目(个)。
百粒重:利用SG考种分析仪测定饱满籽粒(g,仪器由杭州万深检测科技有限公司生产)。
1.4 数据分析
试验采用Microsoft Excel 2016软件和SPSS 20.0软件进行数据处理和统计分析。运用R 3.6.3统计软件(https://www.R-project.org/)基于离差平方和法(Ward)的聚类分析。
2 结果与分析
2.1 苦荞亲本主要农艺性状的差异
两亲本材料在株高、主茎粗、主茎分枝数、主茎节、百粒重等性状上均呈现显著差异。花药迟开裂型黑米荞在上述性状上均高于母本黑米荞(表1)。

表1 亲本农艺性状差异
2.2 苦荞F2群体农艺性状的变异分析
分离群体主要农艺性状表型变异范围较大,株高的变异范围为82~162 cm,平均值为128.7 cm,变异系数在5个性状中最小;主茎粗性状群体平均值为0.71 cm,变异范围最小值为0.27 cm,最大值为1.37 cm,变异系数为20.59%;主茎分枝数的变异系数高达26.39%,变异范围为2~10,平均值为6.59;主茎节数在群体中的平均值为13.08,变异范围最小值为7,最大值为18,变异系数为16.02%;主茎分枝数的变异系数为25.58%,变异范围为0.6~2.02 g,平均值为1.51 g。株高、主茎粗、主茎分枝数、主茎节数峰度和偏度数值符合正态分布,百粒重性状表现为近似正态分布(表2,图1)。
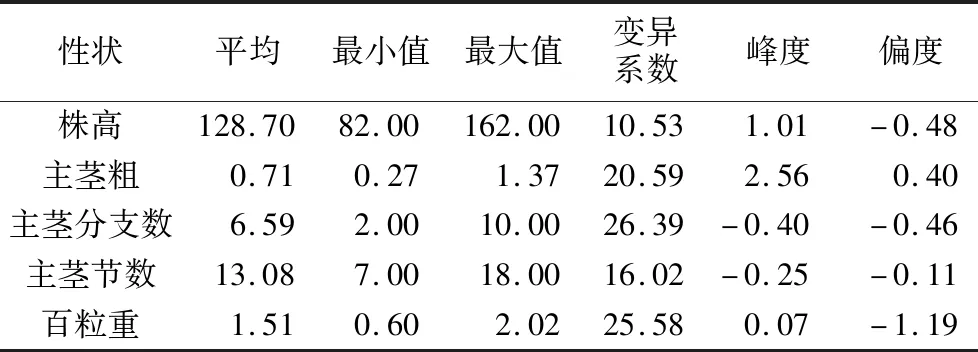
表2 分离群体农艺性状差异
2.3 苦荞F2群体农艺性状相关性分析
由表3可知,株高与主茎粗、主茎分枝数、主茎节数呈极显著正相关,与百粒重呈显著负相关;主茎粗与百粒重不存在相关性,与主茎分枝数、主茎节数呈极显著相关;主茎分枝数与主茎节数呈极显著正相关,与百粒重无相关性;主茎节数与百粒重亦无相关性。
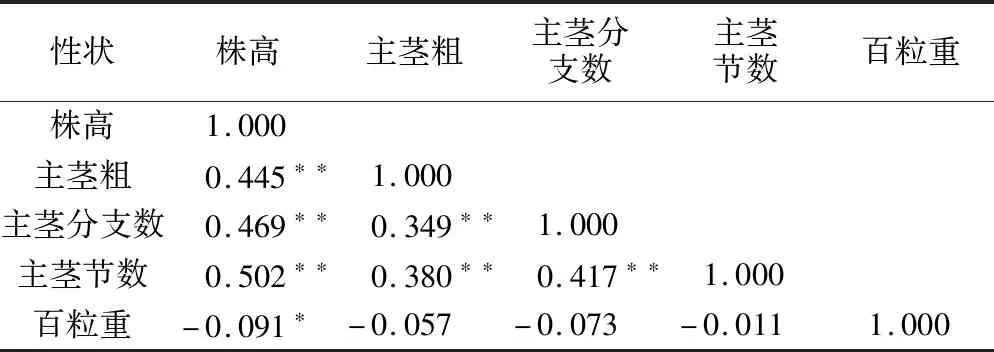
表3 群体农艺性状相关性分析
2.4 苦荞极端种质的初步筛选
综合遗传群体农艺性状进行聚类分析,结果(表4)表明,遗传群体可划分为3个类群,并依据不同亚群
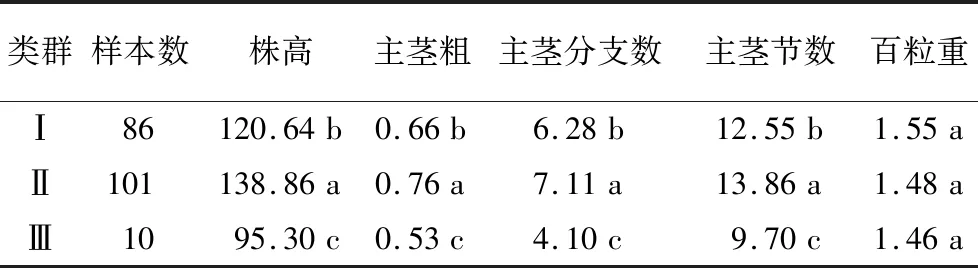
表4 苦荞遗传群体农艺性状聚类和差异分析
5个农艺性状参数进行方差分析。结果显示,Ⅱ群中株高变异范围为129~162 cm,均值为138.86 cm,显著高于Ⅰ、Ⅲ群;在主茎粗、主茎分枝数以及主茎节数等性状上,Ⅱ群均值均显著高于Ⅰ、Ⅲ群;但百粒重数值在3个亚群中并无差异。
3 讨 论
3.1 苦荞农艺性状解析
我国作为荞麦的主产区之一,亦是荞麦的起源地和遗传多样性中心,种质资源丰富,在世界上具有举足轻重的地位[7]。自上世纪80年代初启动对荞麦的系统研究工作以来,荞麦育种工作者采用引种、诱变育种、集团混合选择育种和多倍体育种等方法在荞麦育种改良方面取得了丰硕成果[8-10]。
苦荞农艺性状(株高、主茎分枝数、主茎粗、主茎节数等性状)在驯化过程中受到长期的人工选择,不同品系间变异范围十分广泛。高金锋等对80份西藏苦荞资源进行主茎分枝数、千粒重和单株粒重等产量相关性状测定和评价,结果表明:主茎分枝数变异范围为3.30~15.30,变异系数29%;千粒重变异范围15.40~32.40 g,变异系数12.8%;单株粒重变异范围为2.00~7.10 g,变异系数27%[11]。屈洋等通过对83份来自我国西南苦荞产区(贵州、四川、云南)和北方苦荞产区(陕西、甘肃、宁夏)的种质资源进行了千粒重、主茎分枝数等产量相关性状测定与评价,其中主茎分枝数性状:陕西苦荞资源变异系数最高,为26%,变异范围为4.00~9.50,贵州苦荞资源变异系数最低,为12%,变异范围为4.30~7.30;千粒重性状:宁夏苦荞资源变异系数最高,为26%,变异范围为19.59~32.68 g,甘肃苦荞资源变异系数最低,为6%,变异范围为18.93~21.99 g[12]。李春花等对来源于云南44个县市的48份苦荞资源进行主茎分枝数、千粒重和单株粒重等农艺性状进行测定和评价,结果表明:主茎分枝数变异范围为4.40~9.60,变异系数15%;千粒重变异范围为16.40~22.40 g,变异系数6.6%;单株粒重变异范围为2.70~12.40,变异系数34%[13]。本试验与上述研究研究结果相符,不同生态区苦荞农艺性状的研究表明:苦荞农艺性状在不同品种间遗传变异丰富,且性状分离成连续变异,遗传改良潜力较大,农艺性状的解析将为苦荞品种改良提供重要的理论支撑。
3.2 基于复合农艺性状参数对苦荞选育的影响
荞麦的分子遗传学研究刚刚起步,且苦荞是严格自花授粉作物(花蕊小、开花时间短、人工杂交需借助特用放大镜),导致苦荞遗传群体组配、遗传连锁图谱构建、产量相关性状QTL定位等研究相对缓慢。在配置遗传群体进行遗传学研究的基础上,基于调查的5个农艺性状对197份分离群体单株进行聚类分析,类群间的方差分析揭示:3个亚群间百粒重性状存在细微差距,但差异并不显著。在株高性状上,Ⅲ群变异范围为82~102 cm,均值为95.3 cm,株型高度的降低会起到减少营养生长耗能进而促进单株粒重增加[14-15]。但Ⅲ群在主茎粗、主茎分枝数以及主茎节数等性状上均在亚群中数值最低,主茎分枝数又是决定荞麦有效花序发育的重要正相关性状[16-17],与粮食丰产密切相关。因而在后续品系培育中应侧重在综合指标最优的Ⅰ群中选择。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29 08:55:54
安徽农业大学学报(2022年3期)2022-10-25 12:32:36
种子(2021年7期)2021-08-19 01:46:58
河北科技师范学院学报(2020年1期)2020-07-02 03:37:22
青年歌声(2018年5期)2018-10-29 03:18:40
作文周刊(高考版)(2016年16期)2017-06-01 15:41:00
中国马铃薯(2017年1期)2017-03-02 09:15:46
广东饲料(2016年3期)2016-12-01 03:43:12
党建文汇·下(2016年5期)2016-05-14 11:34:41
奥秘(2016年4期)2016-04-21 17:31:28
