
电针运动区对大脑中动脉闭塞大鼠脑神经再生机制影响研究*
2020-09-17 08:32王玥琪杨春壮
针灸临床杂志 2020年8期
马 英,刘 星,王玥琪,杨春壮
(牡丹江医学院,黑龙江 牡丹江 157011)
缺血性脑中风是常见的急性脑血管病,一直是基础和临床医学科学研究的重点、难点和热点。临床实践证实头针能够改善中风患者的神经功能缺损,促进功能恢复,减轻脑水肿,且取穴精简、操作简便、不易诱发肌痉挛,是中医治疗缺血性中风的常用且有效的方法[1]。本研究采用线栓法制作大脑中动脉闭塞(Middle cerebral artery occlusion,MCAO)大鼠模型,以电针百会穴透太阳穴为干预措施,观测针刺后MCAO大鼠大脑海马内巢蛋白(Nestin)和神经生长因子(NGF)表达的变化规律,从中枢神经功能重建和再生角度,探寻针刺运动区治疗缺血性中风的机制。
1 材料
1.1 动物
所采用SPF级SD大鼠由牡丹江医学院医药研究中心提供[实验动物许可证号:SYXX(黑)2015-007],体质量200~350 g,雌雄各半。
1.2 主要试剂
兔抗Nestin多克隆抗体(BA1289)(博士德生物公司);PrimeScriptTM RT-PCR试剂盒(TaKaRa公司);PCR引物由上海生物工程公司合成。
2 方法
2.1 动物分组
72只大鼠随机分为假手术组、模型组和针刺组,每组各24只。
2.2 MCAO造模
参照马英等相关研究[2]方法制作。先以10%水合氯醛(0.3 mL/100 g)腹腔注射麻醉大鼠,颈前正中纵行切口,长约2.0~2.5 cm,钝性分离并结扎右侧颈总动脉(CCA),并穿一线备用。分离颈内动脉(ICA)和颈外动脉(ECA),结扎ECA,用微动脉夹夹闭ICA,在近CCA分叉处剪一“V”字型切口,将直径为0.2~0.25 mm尼龙线头端涂抹硅胶脂,沿CCA插入ICA,同时放开微动脉夹,轻轻牵拉使栓线进入颅腔,插入深度18~20 mm,稍遇阻力即停,造成MCA阻塞。将CCA上的备线扎紧。1 h后缓慢退出部分线栓进行再灌注。假手术组只做手术暴露颈部动脉,结扎CCA,但不予线栓。
2.3 针刺方法
按中国针灸学会实验针灸委员会实验动物穴位图谱[3],选取大鼠头穴运动区,即“百会”穴透“太阳”穴,由顶骨正中向耳前方斜刺。用0.19 mm×10 mm毫针于术后大鼠清醒即开始行双侧针刺并接G6805-Ⅱ型电针治疗仪(四川恒明科技开发有限公司),断续波,以大鼠安静耐受微颤为度,每天针刺1次,留针30 min。模型组和假手术组略做捆绑,不作针刺处理。
2.4 横木行走实验
治疗第3天、7天、14天后处死前分别进行。横木是长80 cm、宽2.5 cm的方木,距离地面10 cm平放,使大鼠在横木上行走[4]。
评分标准:0分:上即跌落;1分:可滞留但不能行走;2分:可行走,时有跌落;3分:可行走,但左侧肢体不能主动向前移动;4分:行走时有多于50%行程的滑步;5分:行走时有少于50%行程的滑步;6分:行走时没有滑步。
2.5 样本采集
治疗第3天、7天、14天后禁食过夜,各组动物分别随机选取8只用10%水合氯醛腹腔麻醉后,断头取脑,迅速在冰盘上修取MCA供血区(右侧缺血侧)的海马。
2.6 Western blot法检测Nestin蛋白表达
取海马组织加入裂解液,按说明书提取总蛋白,测定蛋白浓度。凝胶电泳分离蛋白质,电转至PVDF膜,用含脱脂奶粉的TBS室温封闭2 h,加入Nestin抗体(兔抗大鼠Nestin抗体,1∶200) 4℃孵育过夜。用含Tween-20的 TBS洗涤3次,加入辣根过氧化物酶标记的羊抗兔IgG(1∶2 000) 室温孵育1 h,重复洗涤3次,ECL液显色,X片显影、定影。用Image-Pro Plus 6.0图像分析测定Western blot条带灰度值,结果以Nestin与内参照甘油醛-3-磷酸脱氢酶(GAPDH)灰度值的比值作为目的蛋白的表达水平。
2.7 实时定量RT-PCR检测NGF mRNA基因表达
采用Trizol一步法试剂盒提取神经组织细胞总RNA,鉴定总RNA纯度、完整性,取总RNA逆转录为c DNA。用于反应的引物分别为:NGF,上游引物为5′-TCCACCCACCCAGTCTTCCA-3′,下游引物为5′-GCCTTCCTGCTGAGCACACA-3′;GAPDH,上游引物为5′-CCTTCCTGGGCATGGAGTCCTG-3′,下游引物为5′-GGAGCAATGATCTTGATCTTC-3。表达采用2-ΔΔCT方法计算,每个样品独立检测3次,所得均值以GAPDH为内参统计。
2.8 统计学方法
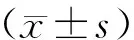
3 结果
3.1 各组大鼠横木行走实验评分
表1显示,假手术组在第3、7、14天横木行走实验评分均为6分,表明神经功能没有出现缺损。与假手术组比较,模型组和针刺组评分均降低,差异有统计学意义(P<0.01),且在第14天,针刺组比模型组评分提高,差异有统计学意义(P<0.01)。
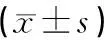
表1 各组大鼠横木行走实验评分比较
3.2 各组大鼠海马Nestin蛋白表达
图1显示,假手术组、模型组和针刺组均可见Nestin表达。
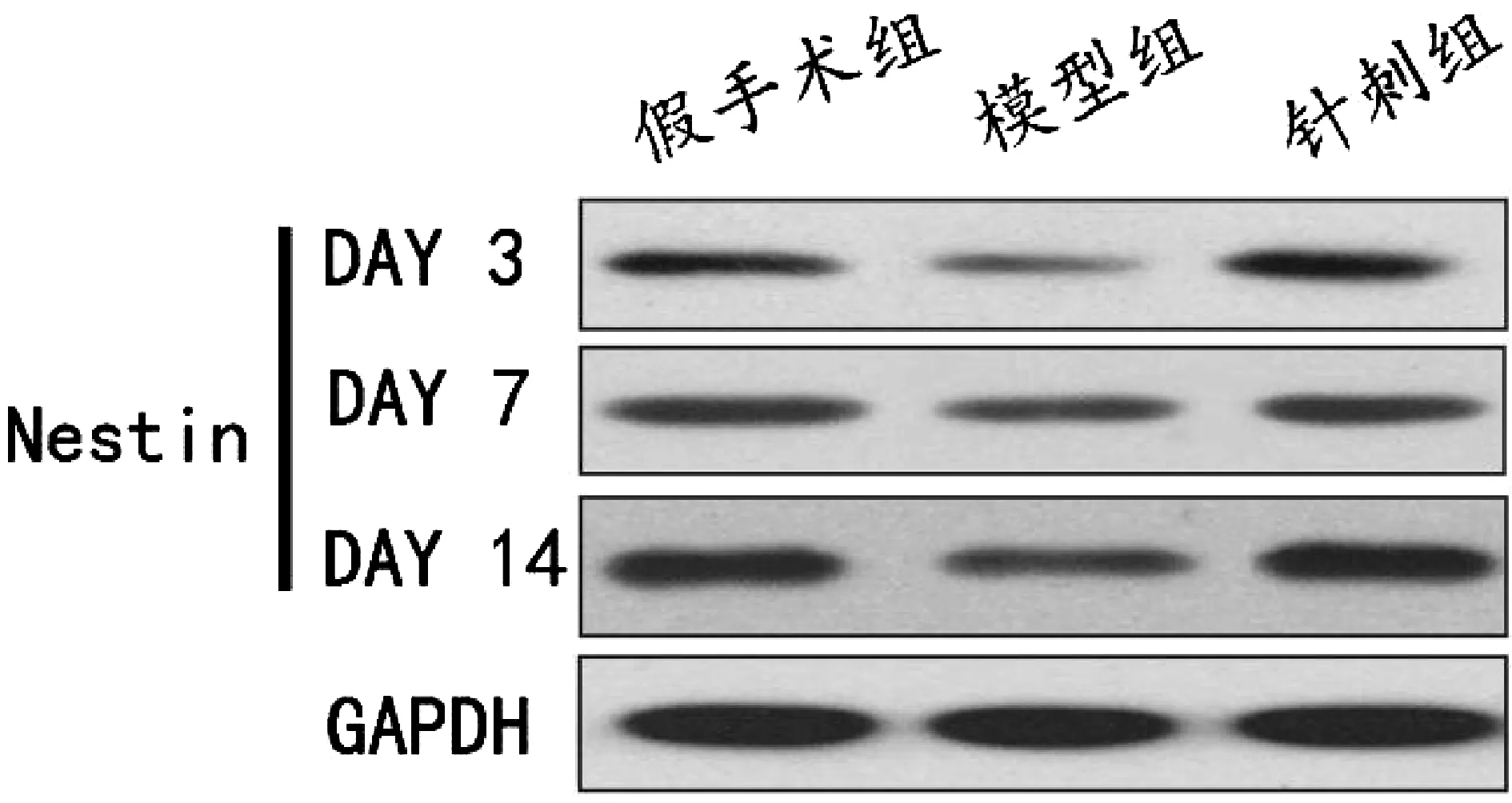
图1 各组脑缺血大鼠Western blot法Nestin蛋白表达
图2显示,Nestin蛋白表达水平模型组与假手术组比较显著降低(P<0.05);针刺组明显高于模型组(P<0.01),且14 d达最高水平。
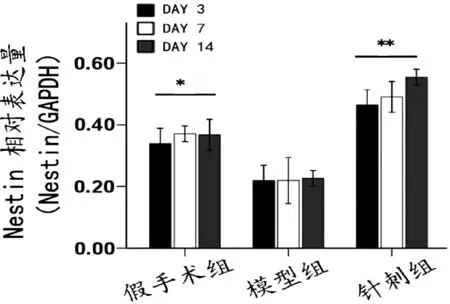
注:与模型组比较,*P<0.05,**P<0.01。图2 各组大鼠海马Western blot法定量检测Nestin蛋白表达
3.3 各组大鼠海马NGF mRNA表达
图3显示,假手术组NGF mRNA表达在术后并无明显区别,模型组自第7天提升量最为明显,接近假手术组的水平,14 d表达下降。针刺组与模型组比较3 d、7 d、14 d都有显著差异(P<0.01)。

注:与模型组比较,**P<0.01。图3 各组大鼠海马NGF mRNA的表达水平
4 讨论
缺血性脑中风属中医“风”“痨”“鼓”“膈”四大难证之一[5]。迄今为止,脑中风造成的神经细胞损伤和死亡尚无确定的治疗方法。因此,如何积极有效地促进脑缺血后受损脑组织的修复与神经功能恢复是目前研究的重点、难点和热点,也始终是人们追求的理想的治疗境界。近现代中医认为中风的主要原因是肝阳化风、气血并逆、直冲犯脑[6-7]。“病变在脑,首取督脉”,督脉有沟通阴阳、总摄诸经之能力,为治疗缺血性脑中风的首选。百会穴为督脉要穴。选取运动区即百会穴透太阳穴,在此处行焦氏头针[8]有醒神开窍、安神定志、平肝熄风的作用。运动区贯穿头部顶、额、颞3区,形成百会-承灵-悬厘-太阳穴的一穴区贯穿多穴的特点,且跨越3条阳经(督脉、足太阳膀胱经、足少阳胆经),从头至足,纵贯全身,实可统调一身之阳气。电针刺激后将产生的生物效应传送至大脑皮层,使穴位刺激点与病灶兴奋点间相互联系、相互作用,激活、加强休眠状态下的脑细胞和受损脑神经细胞的修复能力,促进脑组织的可塑性调节,调节血管舒缩功能,改善神经功能障碍,增进功能恢复[9]。
肢端精细运动和协调功能损害是脑缺血损伤后的主要症状。利用横木行走实验可较好地检测神经功能障碍。本实验结果显示,模型组和针刺组的评分均显著低于假手术组(P<0.01)。模型组自第3天开始评分升高,第7天最高,第14天下降,提示MCAO大鼠神经功能在一定时间段可有部分代偿性自我修复能力,但这种能力持续短暂。与模型组比较,针刺组随着治疗时间的延长横木实验评分呈现明显升高趋势,针刺14 d差异具有差计学意义(P<0.01),表明针刺运动区能明显改善脑缺血损伤后大鼠神经功能缺损的症状,进而降低缺血缺氧对组织的损伤,促进受损区的修复和功能重建,且治疗效果随着针刺时程的延长而提高。
巢蛋白(Nestin)亦称神经上皮干细胞蛋白,是一种Ⅵ型中间丝蛋白,在神经前体细胞一过性表达,当神经前体细胞转化为神经元之后由其他的中间丝蛋白所取代,故与神经细胞分化密切相关[10-11],常作为NSCs(神经干细胞)特异性的标记物。Nestin为 NSCs的自我更新所需[12]。本实验结果表明假手术组、模型组和针刺组海马组织内均可见Nestin表达。脑缺血损伤发生初期,模型组中Nestin蛋白表达水平较假手术组显著降低(P<0.05),这主要是因为大脑中动脉供血区组织破坏严重,难以恢复对损伤的有效修复,不能进一步激活机体的自我保护机制。已有研究表明,机体成年后,室管膜细胞基本处于“静止”状态,而一旦遭受损伤,则可应激进入活跃的分化状态。但这种内源性神经细胞的再生并不能很好修复缺血区损伤,加之诱导NSCs迁移与分化的微环境条件不具备,因此需要采取有效的干预措施来增强NSCs的增殖与分化并促进神经功能的恢复[13]。本实验经电针刺激运动区后,针刺组中Nestin蛋白表达水平明显高于模型组(P<0.01),且14 d组为表达最高。这表明电针运动区可激活内源性NSCs,使其增殖和分化,帮助神经元的再生和细胞骨架的重构,促进神经组织的重建和神经功能的恢复。
NGF对调节神经细胞的生发、分化、维持成熟神经细胞生存以及对损伤神经的修复具有重要意义及临床价值[14-15]。本实验假手术组NGF mRNA表达无明显差异。模型组自第3天开始表达升高,第7天达到高峰,接近假手术组水平,到第14天表达下降。提示脑缺血损伤后可应激性上调NGF表达水平,改善神经组织的病理状态,保护和修复受损神经元,减轻神经损害。但此内源性修复机制上调NGF作用短暂、表达有限,难以对损伤神经元产生全面持久保护,效应有限。针刺组不同时间点NGF mRNA的表达均高于模型组(P<0.01),说明电针运动区可有效提高MCAO大鼠海马内NGF mRNA持续性表达,稳定神经细胞存活的内部微环境和营造适合神经细胞生长的外部微环境,对受损神经细胞发挥保护作用,并对遭受可逆性损伤的神经细胞和胶质细胞进行修复,减缓脑缺血后神经细胞的死亡从而改善相伴的神经功能缺损症状[16]。
综上所述,脑缺血损伤后脑NGF mRNA代偿性增加的自我保护机制促进了神经元和神经细胞的修复,这种自我保护非常有限,而电针刺激运动区的介入可以有效促进NGF mRNA持续性表达,改善神经细胞存活的内、外部微环境,同时激活了处于“静止”状态的内源性NSCs,使其增殖和分化,促进神经组织的重建和神经功能的恢复。
猜你喜欢
上海针灸杂志(2022年10期)2022-10-28
世界科学技术-中医药现代化(2022年3期)2022-08-22
世界科学技术-中医药现代化(2021年8期)2021-12-21
昆明医科大学学报(2021年10期)2021-12-02
昆明医科大学学报(2021年8期)2021-08-13
家庭医药(2021年7期)2021-07-23
天津医科大学学报(2021年3期)2021-07-21
昆明医科大学学报(2021年2期)2021-03-29
昆明医科大学学报(2020年11期)2020-12-28
中华养生保健(2020年4期)2020-11-16
