
碳源和细胞分裂素对‘库尔勒香梨’离体叶片再生不定芽的影响
2020-09-09 01:25孙清荣关秋竹陶吉寒孙洪雁
山东农业科学 2020年8期
孙清荣,关秋竹,陶吉寒,孙洪雁
(山东省果树研究所,山东 泰安 271000)
果树作物的离体叶片不定芽再生是体细胞诱变、外源基因遗传转化及无性系变异研究的基础,是利用生物技术创制新种质、培育突破性品种的基础。离体叶片不定芽再生研究已在很多种果树品种上获得了成功[1-7],梨树的不定芽诱导研究在很多品种上也有成功报道[8-11]。
但不同品种间,其再生难易、再生途径不同,对应的诱导培养基及获得的不定梢再生率及不定梢生长表现等都存在较大差异。
库尔勒香梨是新疆梨系统中最具代表性和特色的优良品种。因梨果香气浓郁、皮薄肉细、汁多渣少等特点,深受消费者的喜爱。但香梨存在果型不整、不抗腐烂病、抗寒性较低、产量不稳定、品质下降等问题,影响了香梨的生产和销售,亟需进行品种改良和优化。体细胞诱变和遗传转化是加速品种改良的有效育种途径。体细胞诱变和遗传转化方法应用的前提是离体叶片高效不定芽再生体系的建立。关于库尔勒香梨离体叶片不定芽再生研究虽有报道[11,12],但存在再生率较低和可重复性较差的问题。本研究的目的是提高库尔勒香梨的离体叶片不定芽再生率及可重复性,克服不定芽玻璃化现象的发生,提高不定芽再生的应用效果,为利用生物技术改良这一品种奠定技术基础。
1 材料与方法
1.1 试验材料
山东省果树研究所组织培养室继代繁殖保存的库尔勒香梨试管苗。
1.2 试验方法
从继代增殖生长4周的试管苗顶端切取展开或微展开的幼嫩叶片,根据叶片大小横切3~5刀,接种到装有不定芽诱导培养基的三角瓶内。每瓶接种10片叶,每处理接种3瓶,计30片叶,算作一个重复,重复3次。叶片接种后先连续暗培养3周,然后转到光下培养。培养至7周时统计产生不定芽的叶片数并计数每片叶的不定芽数,计算叶片不定芽再生率及平均单位叶片不定芽数。
叶片不定芽诱导培养基:基本培养基为NN69,碳源为蔗糖和D-山梨醇,细胞分裂素为BA(3 mg/L,5 mg/L)和 TDZ(1 mg/L,2 mg/L),共组成8种处理(见表1),每一处理都添加0.3 mg/L NAA。
1.3 统计分析
采用DPS v7.5分析软件进行数据统计分析,用最小显著差异(LSD)法进行不同处理间差异显著性比较分析。
2 结果与分析
2.1 碳源对离体叶片再生不定芽的影响
2.1.1 碳源对叶片不定芽再生率的影响 在细胞分裂素相同的条件下,总体上表现D-山梨醇比蔗糖作碳源更有利于提高不定芽再生率,D-山梨醇作碳源的平均再生率(70.2%)明显高于蔗糖的(55.7%);但平均每叶不定芽数两种碳源之间无明显差异,D-山梨醇上平均每叶不定芽数为1.74个,蔗糖上为1.65个(表1)。
细胞分裂素为BA时,D-山梨醇作碳源的不定芽再生率显著高于蔗糖的(表1)。细胞分裂素为TDZ时,在较低浓度(1 mg/L)时,D-山梨醇和蔗糖处理之间无显著差异;在较高浓度(2 mg/L)时,D-山梨醇处理显著高于蔗糖处理。以D-山梨醇作碳源、添加细胞分裂素BA时,库尔勒香梨的不定芽再生率较高,在添加5 mg/L BA的培养基上再生率最高,达96.3%。
2.1.2 碳源对不定芽生长的影响 以D-山梨醇作碳源比蔗糖作碳源诱导产生的不定芽(梢)玻璃化率高(图1 A,B,C,D),最高达60.8%(表1);以蔗糖作碳源的不定芽再生率虽然低于D-山梨醇,但诱导产生的不定芽(梢)玻璃化率低,均在10%以下(表1),正常健壮不定芽(梢)居多(图1 E,F,G,H)。其次,D-山梨醇作碳源还诱导产生只有叶片但没有明显生长点的异常不定芽(图1 I)。
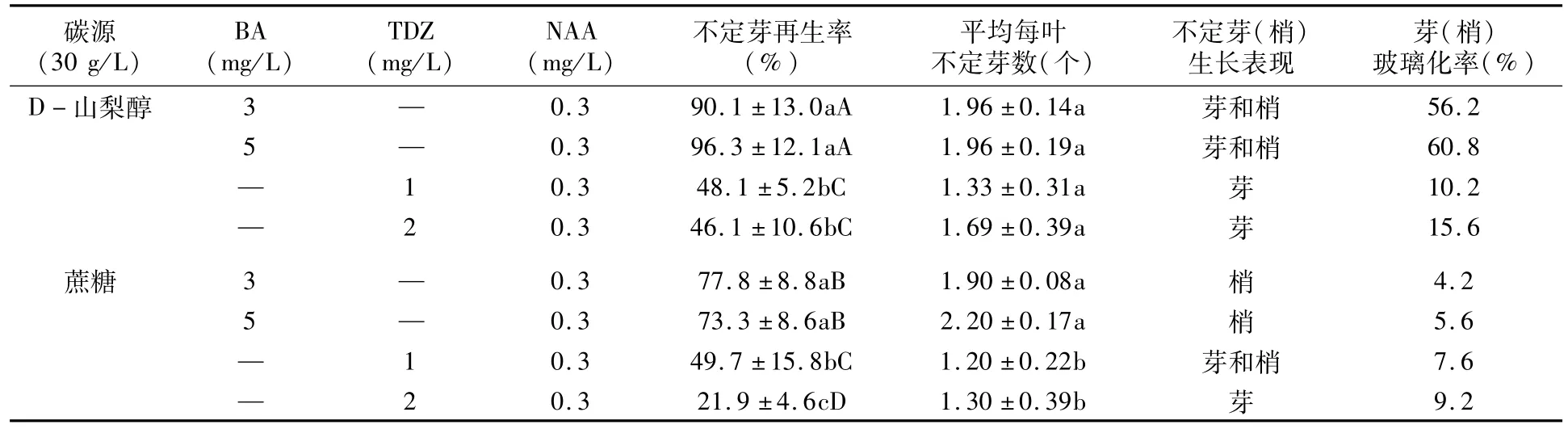
表1 碳源和细胞分裂素对库尔勒香梨叶片再生不定芽(梢)的影响

图1 库尔勒香梨离体叶片不定芽(梢)生长表现
综合比较不定芽的再生率和玻璃化率,蔗糖作为诱导库尔勒香梨离体叶片再生不定芽的碳源要优于D-山梨醇。
2.2 细胞分裂素对库尔勒香梨叶片再生不定芽的影响
2.2.1 细胞分裂素对不定芽再生率的影响 库尔勒香梨离体叶片不定芽(梢)再生率,两种碳源上都表现细胞分裂素BA显著高于TDZ(表1),表明诱导库尔勒香梨离体叶片不定芽(梢)再生BA比TDZ有效。碳源为D-山梨醇时,BA不同浓度间及TDZ不同浓度间的再生率都无显著差异。碳源为蔗糖时,TDZ较低浓度(1 mg/L)处理的不定芽再生率显著高于较高浓度(2 mg/L)处理,而BA不同浓度间差异不显著。平均每叶不定芽数,依据碳源的不同,BA和TDZ表现也不同。当碳源为D-山梨醇时,BA和TDZ之间差异不显著;当碳源为蔗糖时,平均每叶不定芽数BA处理显著高于TDZ处理。在蔗糖和5 mg/L BA组合的诱导培养基上,平均每叶不定芽数最多,为2.2个(表1)。
2.2.2 细胞分裂素对不定芽伸长生长的影响不受供试碳源种类和细胞分裂素浓度的影响,BA诱导的不定芽在诱导培养基上可直接伸长生长形成不定梢(图1 A,B,E,F);而 TDZ诱导的不定芽,除了1mg/L TDZ和蔗糖组合培养基上可直接伸长生长形成不定梢(图1 H)外,其余TDZ处理仅能诱导出不定芽(图1 C,D,H),需转移到不加TDZ的生长培养基上才能继续生长形成不定梢(资料没列出)。表明,BA比TDZ更易诱导不定梢的形成,BA诱导的不定芽不需更换培养基一步培养就可获得不定梢,而TDZ诱导的不定芽需要两步培养才能形成不定梢。
综合不定芽再生率及不定芽生长表现,本研究得出诱导库尔勒香梨离体叶片不定芽再生的最适宜碳源为蔗糖,适宜的细胞分裂素为BA。
3 讨论与结论
碳源种类对库尔勒香梨离体叶片不定芽再生率及其生长表现都有重要影响。对不定芽再生率,山梨醇比蔗糖有效,诱导获得的不定芽再生率高;但对不定芽的生长表现,蔗糖优于山梨醇。这一结果与山梨醇对苹果砧木离体叶片再生不定梢的研究结果不同[5],山梨醇能明显提高苹果砧木‘54-118’的叶片不定梢再生率,且不定梢健壮;而本研究中D-山梨醇虽然诱导库尔勒香梨叶片获得了较高的不定芽再生率,但不定芽玻璃化率也高,不是期望的健康不定芽,应用价值小。这表明基因型不同,适宜的碳源也不同,应根据不同基因型筛选其适宜的碳源。
细胞分裂素BA和TDZ对梨离体叶片不定芽再生的影响因品种不同而不同[13,14]。野生西洋梨的叶片不定芽再生,BA比TDZ有效[13],西洋梨品种‘红考密斯’叶片不定梢再生,BA比TDZ有效[9],‘中梨1号’叶片的不定芽再生,TDZ比BA有效[14]。本研究中,BA比TDZ更有利于提高库尔勒香梨不定芽再生率和促进不定芽的伸长生长,与钟颖等[11]报道的TDZ诱导库尔勒香梨叶片不定芽再生率达84.9%的研究结果不一致,这可能是由于基本培养基和再生途径的不同所致。本研究的基本培养基为NN69,再生途径为直接诱导再生不定芽;而钟颖等的研究所用基本培养基为1/2MS,再生途径为先诱导愈伤,再由愈伤诱导不定芽,不定芽再生周期较长,不是最佳诱导方法。对‘金花’梨,BA和TDZ诱导的叶片不定梢再生率没有显著差异,但对‘鸭梨’,BA能成功诱导不定芽再生,而 TDZ不能诱导不定芽再生[8]。综上所述,虽然TDZ的细胞分裂活性比BA高,但在诱导叶片不定芽再生上,TDZ并不适宜所有品种,应针对不同品种筛选其适宜的细胞分裂素种类及浓度,才能达到最佳再生效果。
猜你喜欢
中国果业信息(2022年4期)2022-11-22
今日农业(2022年16期)2022-09-22
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
今日农业(2020年15期)2020-12-15
中国医学创新(2017年5期)2017-03-16
奥秘(2016年10期)2016-12-17
中学教学参考·理科版(2016年2期)2016-04-05
园艺与种苗(2015年10期)2015-02-27
农产品市场周刊(2014年35期)2014-12-18
中国青年(1991年4期)1991-09-27
