
Anti-MMP-2 and MMP-9 activity of Salsola komarovii Iljin extract and its solvent fractions
2020-09-01 03:38JungHaKilFatihKaradenizGaHyunYuHojunKimJunseKimJungHwanOhJungImLeeChangSukKongYoungwanSeo
Jung-Ha Kil, Fatih Karadeniz, Ga Hyun Yu, Hojun Kim, Junse Kim, Jung Hwan Oh, Jung Im Lee, Chang-Suk Kong,, Youngwan Seo✉
1Marine Biotechnology Center for Pharmaceuticals and Foods, Silla University, Baegyang-daero 700 beon-gil 140, Sasang-gu, Busan 46958, Korea
2Department of Food and Nutrition, College of Medical and Life Sciences, Silla University, Baegyang-daero 700 beon-gil 140, Sasang-gu, Busan 46958, Korea
3Division of Marine Bioscience, Korea Maritime and Ocean University, Busan 49112, Korea
ABSTRACT
KEYWORDS: HT1080; Matrix metalloproteinases-2; Matrix metalloproteinases-9; Salsola komarovii
1. Introduction
Matrix metalloproteinases (MMPs) are a family of endopeptidases responsible for tissue development and regulation of extracellular matrix (ECM) structure. They have crucial roles in ECM-linked biological processes such as wound healing and angiogenesis[1,2].Although MMPs are secreted by various types of cells and have important roles in cellular processes, they are strictly regulated by endogenous inhibitors due to their involvement in pathological changes. Irregulated expression and activities of MMPs have been observed in the onset and progression of serious complications,diseases, and syndromes such as tumor growth and metastasis,arthritis, osteoporosis, and chronic inflammation[3-6]. Classification of MMPs has been organized according to the substrates that act on as gelatinases, collagenases, metalloelastase, matrilysins,enamelysins, stromelysins, and membrane-type[7]. The whole family of MMPs contains zinc at their active sites and their stability is achieved by additional calcium and zinc ions.
Studies provided direct evidence for the involvement of specific MMPs such as MMP-2 (gelatinase A) and MMP-9 (gelatinase B)during cancer-related changes at the cellular level[8]. MMP-2 and MMP-9 are collagen degrading enzymes that specifically act on typeⅠ, Ⅱ and Ⅲ collagens found in ECM[9]. Various cancer types such as ovarian, bladder, lung and oral were characterized by upregulated expression of both enzymes. In vivo studies on MMP-2 revealed that when the MMP-2 activity was attenuated by MMP inhibitors and/or overexpression of tissue inhibitors of metalloproteinases,tumor growth and expansion were significantly halted[10,11]. Similar outcomes were reported with the inhibition of MMP-9 activity. In addition, the introduction of MMP-9 expressing cells to MMP-9 null mice resulted in the aggressive development of squamous cell carcinoma showing the stimulatory effect of MMP-9 overexpression in tumor progression[12]. Overall, increased MMP-2 and MMP-9 activities are correlated with the progression of tumors and high mortality rates in cancer patients. Because of their direct involvement in cancer and other pathologic complications, suppressing the specific MMP expressions and activities was regarded as a valuable mode of preventing and controlling MMP-mediated damages in diseases[13].
Up to date, several studies focused on developing MMP inhibitors from both synthetic and natural origin molecules. Owing to their biocompatibility, diversity and cost-effective nature, natural products have been attracting attention for the discovery of novel MMP inhibitors with higher efficiency and lower side-effects[14,15]. These natural products vary from traditional folk medicine ingredients to the dietary plants. Halophytes, plants of salt marshes, are known for their diverse chemical content rich in phenol-based antioxidant compounds in order to withstand the high salinity environment. Numerous studies reported that extracts of halophytes such as Salicornia herbacea[16], Limonium tetragonum[17]and Glehnia littoralis[18]contained MMP-2 and MMP-9 inhibitors.Salsola komarovii (S. komarovii) Iljin is a halophyte endemic to Korean marshes and studies on Salsola species showed that this genus yielded flavonoids, coumarins, and alkaloids as bioactive constituents[14,19]. Salsola genus is known for its role in Korean folk medicine to treat inflammation and hypertension. However, only a few reports are present for any potential health benefit of S. komarovii and to the best of our knowledge, none of them reports any MMP inhibitory effect[20,21]. As a part of our continuous effort to develop natural origin MMP inhibitors, the present study investigated the MMP-2 and MMP-9 inhibitory effects of S. komarovii crude extract and its solvent fractions. Also, solvent fractions of crude extracts were analyzed to provide insights on the nature of MMP inhibitors that could be isolated from S. komarovii.
2. Materials and methods
2.1. Chemicals, reagents, and cells
Chemicals for the extraction of samples were obtained from Samchun Chemical (Pyeongtaek, Korea) unless otherwise noted.HT1080 human fibrosarcoma cell line was purchased from Korean Cell Line Bank (KCLB no. 10121; Seoul, Korea). Chemicals and reagents for cell culture and maintenance were obtained from Gibco(Carlsbad, CA, USA) unless otherwise noted. Materials and reagents for assays were acquired from Sigma-Aldrich (St. Louis, MO, USA)unless otherwise specified. Antibodies for Western blotting were purchased from Santa Cruz Bitechnology (Santa Cruz, CA, USA).
2.2. Plant material and extraction
S. komarovii Iljin was collected by hand in September 2008 at Dabudo, Ansan City, Gyeonggi-do, South Korea. Collected specimens of S. komarovii Iljin were air-dried under shade and ground to powder. The obtained powder-like sample was extracted for 24 h at room temperature, using methylene chloride (CH2Cl2,MC) and methanol (MeOH) as extraction solvents, consecutively.Two different crude extracts (MC and MeOH) were obtained by drying the extracts under pressure. These extracts were used in subsequent experiments after preparing a 10 mg/mL stock solution in 10% (v/v) dimethyl sulfoxide. MC and MeOH crude extracts were then combined and subjected to fractionation between CH2Cl2and H2O after dissolving the combined extract in CH2Cl2. The CH2Cl2layer was further partitioned between n-hexane and 85% aqueous(aq.) MeOH while the H2O layer was separated by the addition of n-BuOH. This whole procedure yielded four solvent-partitioned fractions of S. komarovii crude extract (SKI); n-hexane (0.28 g), 85%aq. MeOH (0.13 g), n-BuOH (0.51 g), and H2O (3.08 g).
2.3. Cell culture and cytotoxicity of extracts and solvent fractions
Human HT1080 fibrosarcoma cells were used in the current study due to their ability to express MMPs when treated with phorbol 12-myristate 13-acetate (PMA). Cells were cultured with Dulbecco’s modified Eagle medium and supplemented with 10% fetal bovine serum. Until the time of analysis, cells were kept in 37 ℃ incubators with a humidified atmosphere containing 5% CO2.
The potential cytotoxicity of the SKI dichloromethane (SKIMC) (10, 50 and 100 μg/mL) and methanol (SKI-MeOH) crude extract (10, 50 and 100 μg/mL) and SKI fractions (10, 50 and 100 μg/mL) was assessed by colorimetric cell viability assay based on previously described MTT formazan conversion assay[17]. Briefly,HT1080 cells were cultured to 80% confluence in 96-well plates prior to the addition of various concentrations of samples. The cell culture medium was removed after 2 days of incubation and wells were added 100 μL MTT solution (1 mg/mL). The plate was stored at 37 ℃ for 4 h without light. Finally, 10 μL of 100%DMSO was added to each well after the removal of MTT solution and absorbance values (540 nm) were measured using GENios FLmicroplate reader (Tecan Austria GmbH, Grodig, Austria).The viability of the sample-treated cells was given as a percentage compared to the untreated control group.
2.4. Gelatin zymography
Effects of SKI and SKI fractions on the production of MMP-2 and MMP-9 in PMA-treated HT1080 cells were analyzed by the gelatinolytic ability of the enzymes in the cell culture medium using gelatin zymography. HT1080 cells were grown to confluence in 6-well plates and treated with 10 ng/mL PMA along with samples except for the untreated control group. Following incubation for 24 h, conditioned cell culture media were harvested from each well.Conditioned medium was then transferred to gels and MMP-2 and MMP-9 mediated digestion of the gelatin in the gel was stimulated by storing gels in a digestion buffer (10 mM CaCl2, 50 mM Tris-HCl and 150 mM NaCl) for 48 h. MMP-2 and MMP-9 activities were detected with CAS-400SM Davinch-Chemi imagerTM(Davinch-K,Seoul, Korea) as digested clear zones on the gel following Coomassie Blue staining. Production of MMP-2 and MMP-9 was then calculated densiometrically and given as percentage compared to untreated PMA-stimulated control group.
2.5. Enzyme-linked immunosorbent assay for MMP-2 and MMP-9 release
HT-1080 cells were cultured to confluence in 6-well plates and treated with PMA (10 ng/mL), along with SKI and SKI fractions(except for the untreated control group) for 24 h. MMP-2 and MMP-9 release from cells were investigated in cell culture medium by an ELISA kit (R&D Systems, Inc., Minneapolis, MN, USA)according to the manufacturer's instructions.
2.6. Reverse transcription-polymerase chain reaction analysis
HT1080 cells were grown into confluence in 24-well plates prior to treatment. Total RNA was obtained from blank (non-stimulated untreated), control (PMA-stimulated untreated) and test (PMA-stimulated sample treated) groups using Trizol reagent (Invitrogen,Carlsbad, CA, USA). Two micrograms of total RNA were used for two-step reverse transcription. A mixture containing total RNA (2 μg), oligo dT, random primers in RNase-free water was denatured for 5 min at 70 ℃. Reverse transcription was carried out using M-MLV reverse transcriptase (Thermo Fisher Scientific, Waltham,MA, USA) following manufacturer’s instructions in a T100 Thermal Cycler (Bio-Rad, Hercules, CA, USA) with two steps of 42 ℃ for 60 min and 72 ℃ for 5 min. Amplification of reverse-transcribed cDNA was carried out with common PCR protocols using a master mix containing iTaq DNA Polymerase (Bio-Rad) and gene specific forward and reverse primers as previously reported[22]. Detection of genes was accomplished by agarose (1.5% w/v) gel electrophoresis(30 min at 100 V) after 30 cycles in a thermal cycler (T100, Bio-Rad) each consisted of 45 s at 95 ℃, 1 min at 60 ℃ and 45 s at 72 ℃.Gels were stained by keeping in ethidium bromide solution (0.1% w/v) and imaged with a UV light source (CAS-400SM Davinch-Chemi imagerTM, Davinch-K, Seoul, Korea).
2.7. Western blotting
Detection of protein levels was performed using standard Western blot protocols as previously described[17]. Briefly, HT1080 cells were grown into confluence in 6-well plates prior to treatment.Total protein was extracted from blank (non-stimulated untreated),control (PMA-stimulated untreated) and test (PMA-stimulated sample treated) groups by addition of 1 mL RIPA buffer (Sigma-Aldrich, St. Louis, MO, USA) to wells and vigorous pipetting.Cell lysates were assessed for their protein content with a BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL, USA).A part of total protein lysate (20 μg) was separated by SDS-PAGE(4% stacking and 10% separating gels). Proteins on gels were transferred to polyvinylidene fluoride membrane (Amersham; GE Healthcare, Little Chalfont, UK) for immunoblotting. Blocking of membranes was occurred by keeping membranes in 5% skim milk(v/v in TBS-T buffer) for 4 h on a shaking incubator. The blocked membrane was hybridized at 4 ℃ overnight with specific primary antibodies (1:1 000) in 1×TBST buffer containing 5% bovine serum albumin (m/v). Next, the membrane was incubated for 2 h at room temperature with horseradish peroxidase-conjugated secondary antibodies specific to primary antibodies. Membranes were stained with ECL kit (Amersham) according to manufacturer’s instructions and the protein bands were imaged using CAS-400SM Davinch-Chemi imagerTM(Davinch-K).
2.8. Statistical analysis
Numerical results were given as the average of three independent experiments ± SD where applicable unless otherwise noted. Groups in the same data series were subjected to one-way analysis of variance (ANOVA) with post-hoc Duncan’s multiple range test for statistical analysis (SAS v9.1, SAS Institute, Cary, NC, USA) and differences were defined significant at P<0.05 level.
3. Results
3.1. Effect of SKI-MC and SKI-MeOH on cell viability
Crude extracts of S. komarovii (SKI) from two different solvents,dichloromethane (MC) and methanol (MeOH) were tested for their effects on activities and expressions of MMP-2 and MMP-9. Cell viability assay revealed that both SKI-MC and SKI-MeOH with concentrations from 10 μg/mL to 100 μg/mL did not show any toxic effect on human HT1080 fibrosarcoma cells (Figure 1A). All test groups were considered as viable as the untreated control group.Cytotoxicity assay suggested that the inhibitory effect of SKIs was not due to their cytotoxicity but intervention with MMPs or their production pathways.
3.2. Effect of SKI-MC and SKI-MeOH on MMP-2 and MMP-9 release by gelatin zymography assay
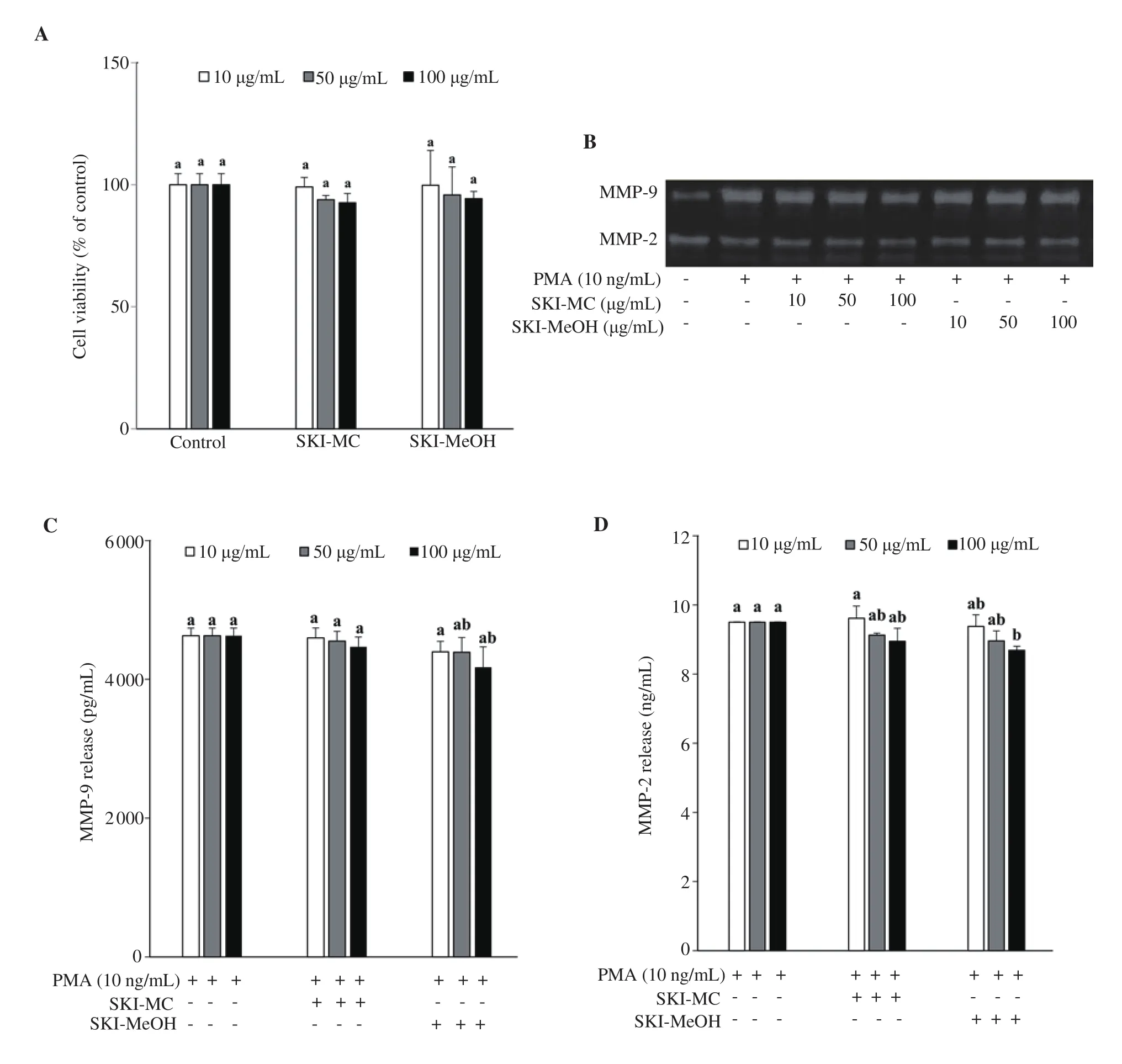
Figure 1. Effect of Salsola komarovii crude extracts (SKI) on the viability of HT1080 human fibrosarcoma cells analyzed by MTT assay (A). Cell viability was given as the percentage of untreated control cells (Control). Gelatin zymography (B) shows the inhibitory effects of SKI on activities of matrix metalloproteinases (MMP) -2 and MMP-9 from phorbol 12-myristate 13-acetate (PMA)-stimulated HT1080 cells. Extracellular secretion levels of MMP-9 (C)and MMP-2 (D) were analyzed by ELISA. a-bMeans with different letters are significantly different at P<0.05 level. SKI-MC: Methylene chloride (CH2Cl2)extract of Salsola komarovii crude extracts, SKI-MeOH: Methanol extract of Salsola komarovii crude extracts.
Treatment with SKI-MC and SKI-MeOH decreased the production of MMP-9 according to the gelatin zymography assay (Figure 1B). SKI-MC showed a dose-dependent decrease in the MMP-9 degraded areas where, at the concentration of 100 μg/mL, it was 73% of the untreated control cells. On the other hand, SKI-MeOH did not exert any dose-dependency, however, decreasing the MMP-9 degraded areas to the 92%, 96% and 95% of the untreated control cell at 10, 50 and 100 μg/mL treatment, respectively. Both samples exerted similar effects on MMP-2, however, slightly lower than that of MMP-9. At 100 μg/mL, degradation was 82% of the untreated control for SKI-MC, and 95% for SKI-MeOH. Results indicated that SKI-MeOH was unable to affect the production of MMP-9 while SKI-MC treated cells released significantly less MMP-9. Due to difference in the polarity of the solvents, it was suggested that direct inhibitors of MMP-9 enzyme release and/or production was of less polar nature as they were abundant in MC extract rather than MeOH extract.
3.3. Effect of SKI-MC and SKI-MeOH on MMP-2 and MMP-9 expression and release by ELISA, RT-PCR, and Western blotting assay
The release of MMP-2 and MMP-9 enzymes to the cell culture medium by the stimulated HT1080 cells was analyzed by ELISA.Cells induced by PMA but not treated with any samples were observed to release 4 629 pg/mL MMP-9 and 9.50 ng/mL MMP-2(Figure 1C and D). Treatment with SKI-MC did not induce any notable change in MMP-9 release at all concentrations treated and decreased MMP-2 release slightly to 8.95 ng/mL at 100 μg/mL treatment. Although SKI-MeOH produced better results in decreasing MMP-2 and MMP-9 release compared with SKI-MC, it was only able to lower MMP-9 amount to 4 175 pg/mL, and MMP-2 amount to 8.70 ng/mL at the highest treated concentration.
Effects of SKI-MC and SKI-MeOH on the intracellular expression of mRNA and protein levels of MMP-2 and MMP-9 were analyzed by RT-PCR and Western blotting, respectively. Both SKI-MC and SKI-MeOH were able to decrease the PMA-induced elevated MMP-2 and MMP-9 mRNA levels (Figure 2). MMP-9 mRNA levels were decreased by 11.20% and 27.19% after 100 μg/mL treatment with SKI-MC and SKI-MeOH, respectively. Similar results were observed for MMP-2 mRNA levels, which were decreased to 62.00% and 42.67% of untreated control cells after 100 μg/mL treatment with SKI-MC and SKI-MeOH, respectively. Results of Western blotting indicated decreased levels for both MMP-2 and MMP-9 after SKI treatment. Treatment with 100 μg/mL SKI-MC decreased the MMP-2 protein levels to 65.04% and MMP-9 protein levels to 31.95% of the untreated control cells while the results for 100 μg/mL SKI-MeOH treatment were 48.46% and 35.67% of the untreated control cells (Figure 3).
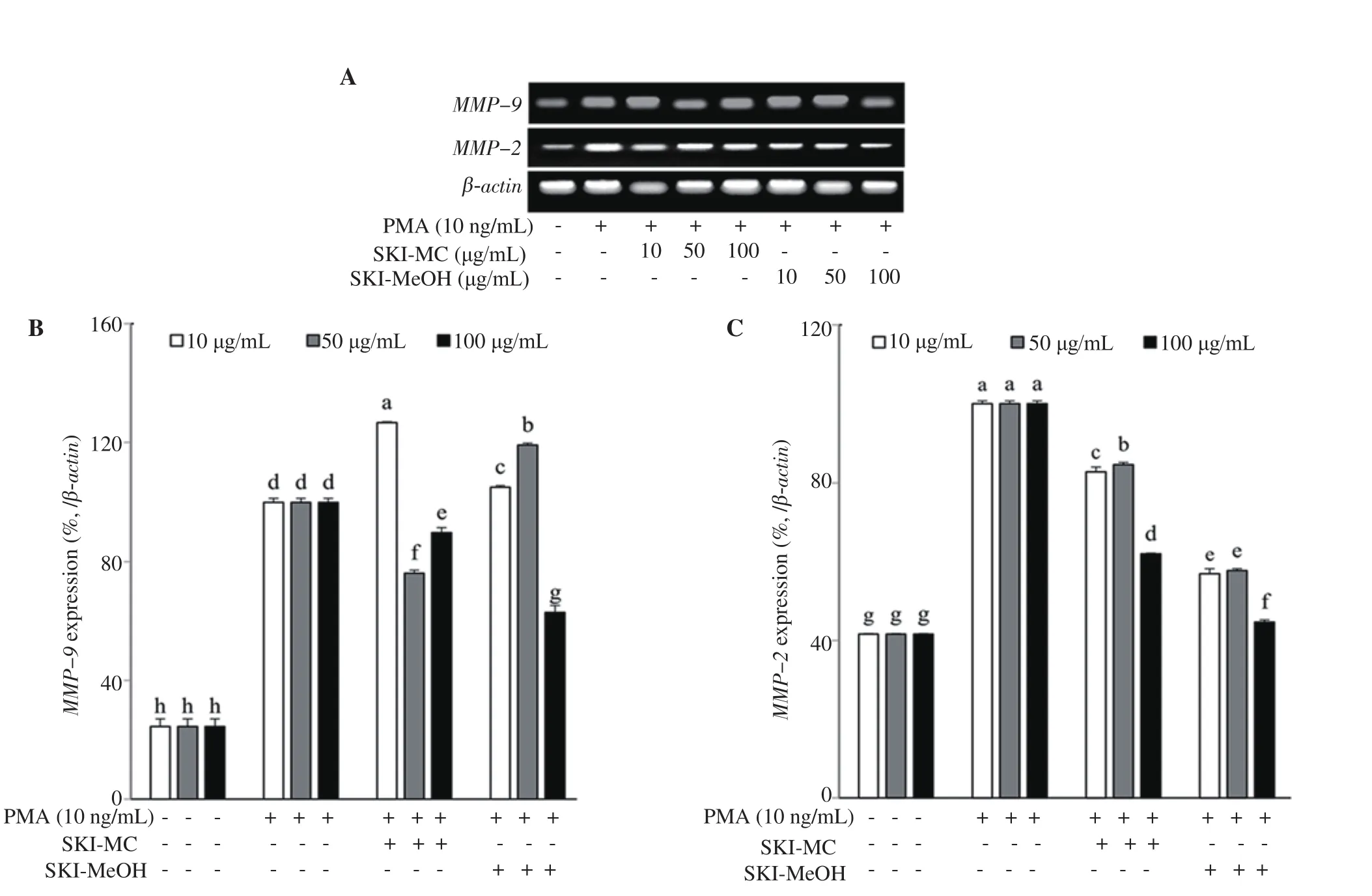
Figure 2. Effect of SKI-MC and SKI-MeOH fractions on mRNA expression levels of the MMP-2 and MMP-9 analyzed by RT-PCR (A). mRNA levels were quantified by densitometry and normalized against β-actin (B&C). Effect of fractions on mRNA levels was given as the percentage of the phorbol 12-myristate 13-acetate (PMA)-stimulated untreated control cells. a-gMeans with different letters are significantly different at P<0.05 level. SKI-MC: Methylene chloride (CH2Cl2) extract of Salsola komarovii crude extracts, SKI-MeOH: Methanol extract of Salsola komarovii crude extracts.
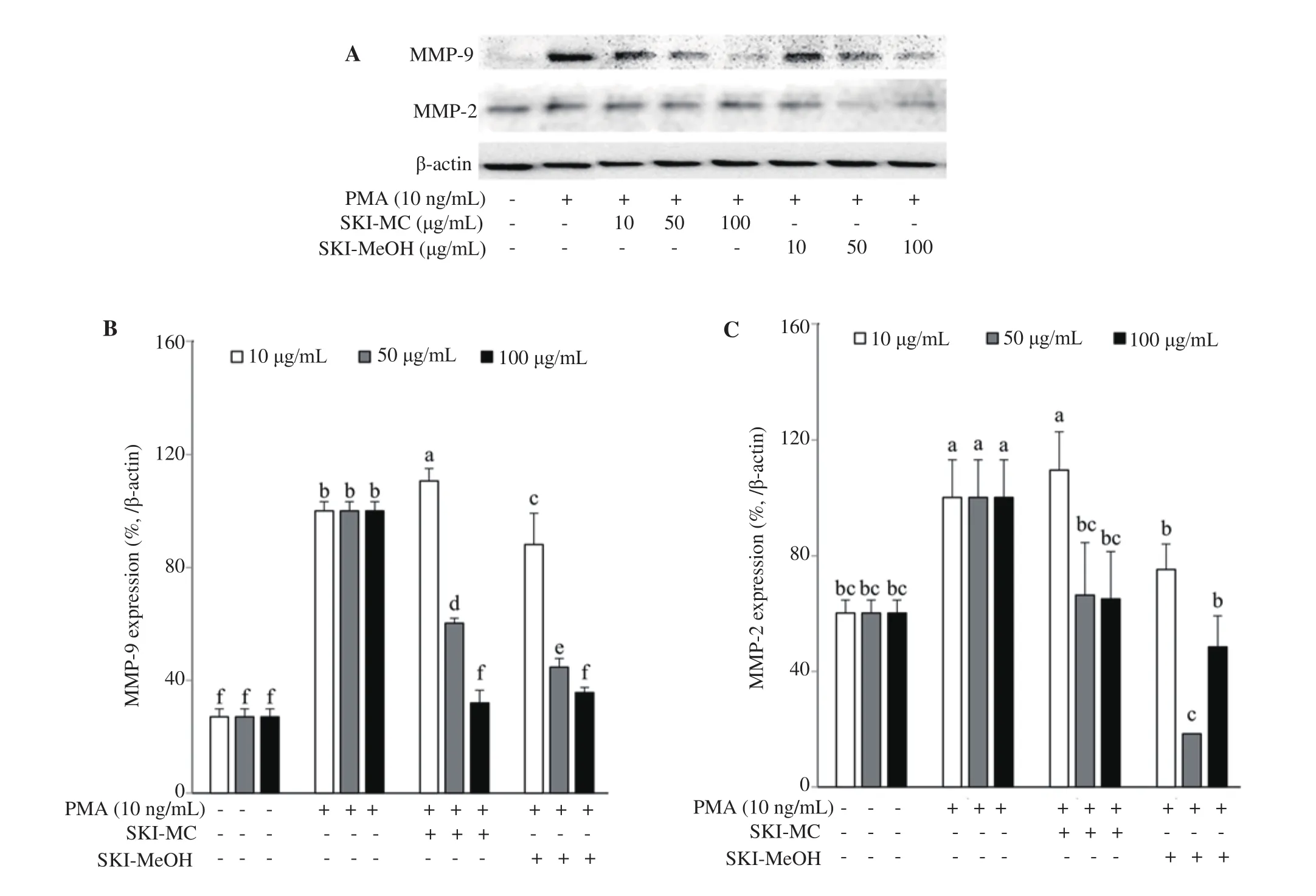
Figure 3. Effect of SKI-MC and SKI-MeOH fractions on protein levels of the MMP-2 and MMP-9 analyzed by Western blotting (A). Protein levels were quantified by densitometry and normalized against β-actin (B&C). Effect of fractions on protein levels was given as the percentage of the phorbol 12-myristate 13-acetate (PMA)-stimulated untreated control cells. a-fMeans with different letters are significantly different at P<0.05 level. SKI-MC: Methylene chloride (CH2Cl2) extract of Salsola komarovii crude extracts, SKI-MeOH: Methanol extract of Salsola komarovii crude extracts.
3.4. Effect of SKI fractions on cell viability
Both SKI-MC and SKI-MeOH showed notable MMP inhibiting activities against MMP-2 and MMP-9 suggesting that the SKI contained different types of compounds that could act as inhibitors.Both crude extracts were then combined and fractionated by different solvents into SKI n-hexane, 85% aq. MeOH, n-BuOH, and H2O fractions. Effects of SKI fractions on MMP-2 and MMP-9 activity,expression and release were analyzed and compared regarding the difference of chemical constituents of each fraction.
Cellular toxicities of SKI fractions were investigated by MTT assay.At tested concentrations (10, 50 and 100 μg/mL), all SKI fractions were shown not to drop viability of HT1080 cells under 80% of the untreated control group except 100 μg/mL SKI 85% aq. MeOH fraction treatment that decreased cell viability to 26% (Figure 4A).Hence further analysis of SKI fractions was performed using only 10 and 50 μg/mL concentrations.
3.5. Effect of SKI fractions on MMP-2 and MMP-9 activity by gelatin zymography assay
Activities of MMP-2 and MMP-9 after treatment with 10 and 50 μg/mL SKI fractions were analyzed using gelatin zymography.Among fractions, 85% aq. MeOH was the most active one in decreasing the activity of both MMP-2 and MMP-9. Depicted from the clear zones created by the enzymatic gelatinase activity of MMPs, 85% aq. MeOH fraction was able to decrease the MMP-9 activity to 1% of the untreated control cells, a level even lower than that of unstimulated untreated blank group (Figure 4B). n-Hexane fraction followed the 85% aq. MeOH fraction in terms of enzyme inhibiting efficiency.
3.6. Effect of SKI fractions on MMP-2 and MMP-9 expression and release by ELISA, RT-PCR, and Western blotting assay
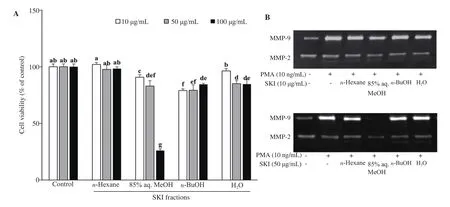
Figure 4. Effect of SKI solvent fractions (n-Hexane, 85% aq. MeOH, n-BuOH, and H2O) on the viability of HT1080 human fibrosarcoma cells analyzed by MTT assay (A). Cell viability was given as the percentage of untreated control cells (Control). a-gMeans with different letters are significantly different at P<0.05 level. Gelatin zymography (B) showing the inhibitory effects of SKI solvent fractions on enzymatic activity of MMP-2 and MMP-9 from phorbol 12-myristate 13-acetate (PMA)-stimulated HT1080 cells. SKI: Salsola komarovii crude extracts.
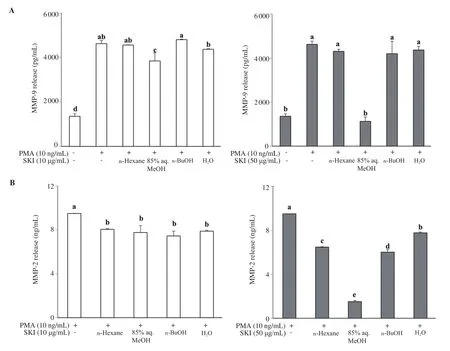
Figure 5. Effect of SKI solvent fractions (n-Hexane, 85% aq. MeOH, n-BuOH, and H2O) on the extracellular secretion levels of MMP-9 (A) and MMP-2 (B).Cells were treated with fractions at indicated concentrations and the culture medium was investigated by an ELISA kit. a-eMeans with different letters are significantly different at P<0.05 level. SKI: Salsola komarovii crude extracts.
Similar patterns to that of gelatin zymography results were obtained from MMP-2 and MMP-9 release analysis of stimulated cells treated with SKI fractions. Cells treated with 85% aq. MeOH showed the lowest amount of MMP release compared to stimulated untreated control cells. At the concentration of 50 μg/mL, 85% aq.MeOH fraction was able to decrease the MMP-9 release to 1 125 pg/mL from the 4 629 pg/mL value of control cells (Figure 5). The MMP-2 amount was also decreased to 1.52 ng/mL from 9.50 ng/mL.To confirm the significant MMP inhibitory effect of 85% aq. MeOH fraction, effects of SKI fractions on the levels of mRNA and protein expression of MMP-2 and MMP-9 were analyzed by RT-PCR and Western blotting, respectively. At the 50 μg/mL concentration, the n-hexane fraction was more effective in suppressing MMP-2 mRNA expression than other fractions by decreasing the levels to 53.77%of untreated control compared to 75.26% of 85% aq. MeOH.However, 85% aq. MeOH strongly suppressed the MMP-9 mRNA levels to 58.28% of the untreated control, being the most active fraction (Figure 6A). Different results were obtained from protein levels of MMP-2 and MMP-9 following SKI fraction treatment.Treatment with 50 μg/mL 85% aq. MeOH fraction decreased total MMP-9 protein amount to 88.82% of the untreated control group while lowering the MMP-2 protein levels to 48.80% (Figure 6B).On the other hand, n-hexane fraction was the most effective fraction that decreased total MMP-9 (20.84%) and MMP-2 (43.74%) levels compared to the untreated control group (Figure 6B).
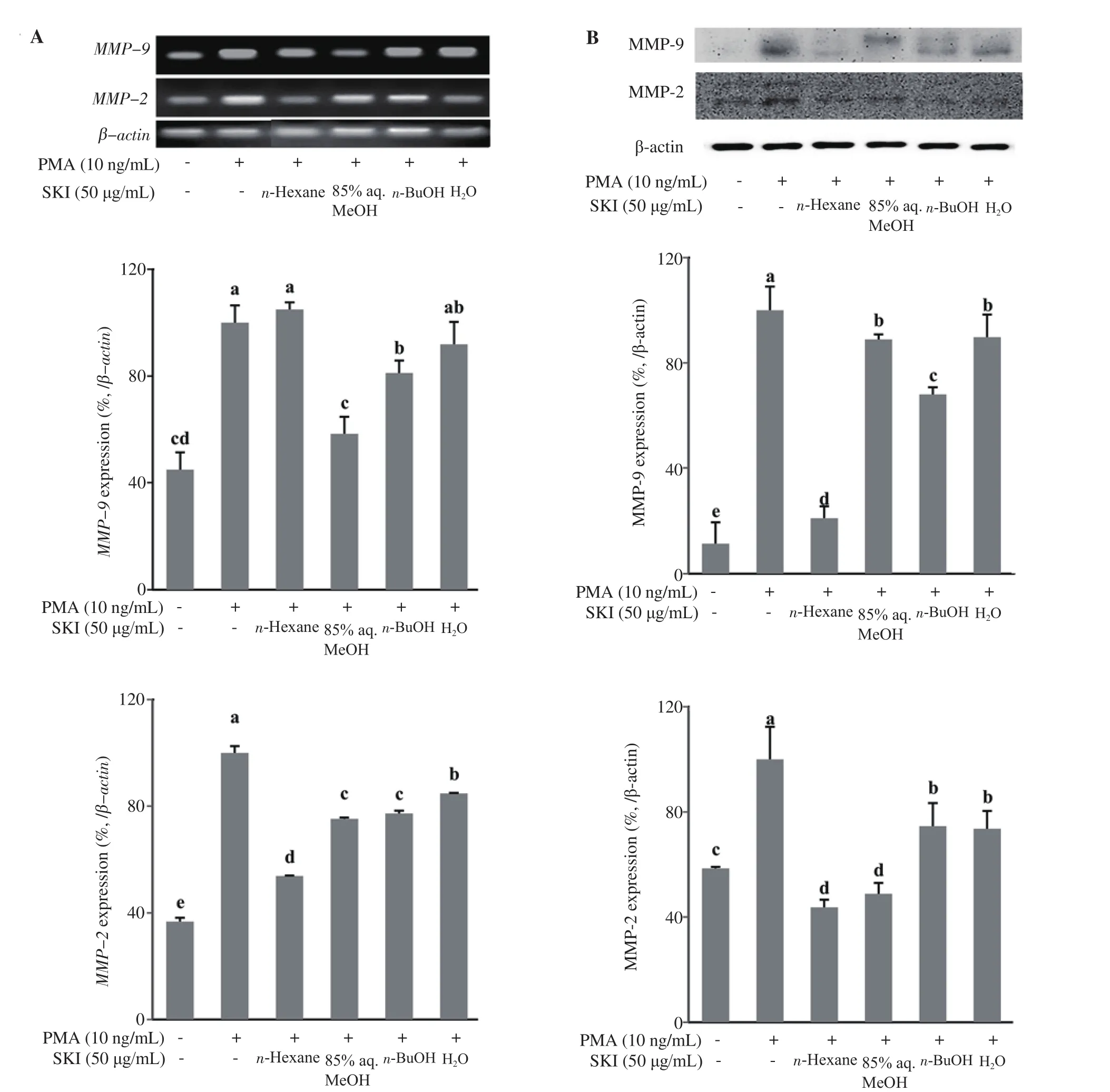
Figure 6. Effect of SKI solvent fractions (n-Hexane, 85% aq. MeOH, n-BuOH, and H2O) on mRNA expression (A) and protein (B) levels of the MMP-2 and MMP-9 analyzed by RT-PCR and Western blotting assays, respectively. Levels of expression were quantified by densitometry and normalized against β-actin. Effect of fractions on mRNA and protein levels was given as the percentage of the PMA-stimulated untreated control cells. a-eMeans with different letters are significantly different at P<0.05 level. SKI: Salsola komarovii crude extracts.
4. Discussion
Overexpression of MMPs is linked with several healththreatening complications varying from tumor growth and metastasis to inflammation and asthma[5]. Regulating the MMP activity and expression could decrease the harmful outcomes of these complications and in some cases prevent and/or treat the symptoms including cancer progression[22]. Natural products provide diverse sources of MMP inhibitors, thus the interest in natural pharmaceuticals is on the rise. In this context, halophytes, plants of salty marshes, were studied and several species were found to contain MMP inhibiting compounds due to unique and challenging environments that they inhabit[23,24]. S. komarovii Iljin is a halophyte found on the shores of Korea and has roles in the daily diet and traditional medicine of Korea, Japan, China, and Eastern Russia.Studies on Salsola species reported that this genus has the potential to be used for their health beneficial effects mainly due to their flavonoid and coumarin contents[25]. However, there is few studies on S. komarovii and none of them reported its effect against MMPs.Its effect against two types of MMP, MMP-2 and MMP-9, which play pivotal roles in tumor metastasis, was analyzed in the current study.
Results suggested that SKI contained MMP inhibitory compounds with different chemical backbone and speculatively different action mechanisms. 85% aq. MeOH fraction was significantly effective in inhibiting the enzymatic activity of MMP-2 and MMP-9 according to gelatin zymography assay. Also, the expression of mRNA and protein levels were downregulated by the n-hexane fraction, slightly more effective than 85% aq. MeOH. Overall, n-hexane was the second most active fraction to suppress the MMP-2 and MMP-9 levels followed by n-BuOH fraction. Among all tested fractions,H2O fraction was ranked last in terms of inhibiting MMP-2 and MMP-9 activity, release, and expression. Considering the previously isolated compounds from S. komarovii extracts and their chemical structures[21], it was suggested that polyphenolic and flavonoid ingredients of SKI, which were suggested to be abundant in 85%aq. MeOH fraction of halophyte extracts[26,27], were responsible for direct enzymatic inhibition of MMP-2 and MMP-9. On the other hand, coumarin and quercetin derivatives, which were reported to be isolated from both hexane and methanol extracts[28], were suggested to be in n-hexane fraction and responsible for the downregulation of intracellular MMP expression and release. To the best of our knowledge, this is the first study reporting the MMP inhibitory effect of S. komarovii Iljin. In conclusion, the current study suggested that S. komarovii Iljin is a potential source of inhibitors against MMP-2 and MMP-9, which are critical enzymes in cancer metastasis and chronic inflammation. However, further studies to elucidate bioactive constituents from active extracts and characterize their mechanisms of action are needed and would provide valuable insights towards the utilization of S. komarovii as a natural product.
Conflict of interest statement
We declare that there is no conflict of interest.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (No. 2016R1D1A1B03932769) and by the Ministry of Science and ICT (No. 2019R1F1A1059325).
Authors’ contributions
JHK, JIL, CSK and YS conceived the idea and designed the study.JHK, FK, GHY, HK and JHO carried out the in vitro assays and interpreted the data. JK, JHO and JIL carried out the extraction and fractionation. FK wrote the manuscript. JHK, and JHO performed the necessary revisions. CSK and YS provided the resources and supervised the study.
 Asian Pacific Journal of Tropical Biomedicine2020年10期
Asian Pacific Journal of Tropical Biomedicine2020年10期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Anti-diabetic properties and bioactive compounds of Teucrium polium L.
- Chemical characterization, docking studies, anti-arthritic activity and acute oral toxicity of Convolvulus arvensis L. leaves
- Mucus from different fish species alleviates carrageenan-induced inflammatory paw edema in rats
- Rice bran hydrolysates induce immunomodulatory effects by suppression of chemotaxis, and modulation of cytokine release and cell-mediated cytotoxicity