
油菜素内酯对盐胁迫下水稻种子萌发的影响①
2020-08-31 07:10侯会云
热带农业科学 2020年7期
侯会云
(台前县农业局 河南濮阳457600)
土壤盐碱化是世界农业发展面临的一个重要难题。据不完全统计,世界盐碱地占总陆地面积的7.6%,大约有30多个国家遭受土壤盐碱化困扰,土壤盐碱化不仅破坏土壤结构、理化性质,造成土壤功能退化,还会导致植物渗透胁迫,阻碍植物对矿质养分的吸收,严重影响植物的生长发育[1]。因此,盐碱地的农业发展成为目前亟待解决的问题之一。
水稻是一种盐敏感作物,盐胁迫是影响水稻高产稳产的主要限制因子之一[2]。一般认为,低浓度的盐对种子的正常发芽没有影响,但随着盐浓度进一步提高,种子发芽受到限制,高浓度造成的盐胁迫导致水稻种子发芽率降低,根系活力受到抑制,从而影响营养物质的吸收和同化。李旭红等[3]研究表明,盐胁迫下的种子受到离子胁迫、渗透胁迫和化学抑制等不良影响,导致组织水分亏缺、营养失衡,严重影响水稻的正常生长;闫先喜等[4]研究认为,在种子吸胀过程中盐胁迫会破坏细胞膜,使得细胞膜透性增大,从而导致胞液外渗,种子萌发受阻。因此,提高水稻对盐胁迫的抵抗能力是当前应对盐胁迫的有效方法。油菜素内酯(BR)是广泛存在于植物体中的天然激素,是一种广谱性植物生长调剂剂,能够充分激发植物潜能,促进作物生长,提高作物耐冷、抗病、抗盐碱能力[5]。吴杨等[6]研究结果表明,在盐碱胁迫下,外源施加BR 可维护细胞结构的完整性,增强大豆幼苗耐盐碱能力。栗露露等[7]研究表明,BR 能够调控水稻的耐盐性。魏湜等[8]认为,外源BR 能够缓解盐胁迫对玉米幼苗的生理损伤。目前油菜素内酯主要用于提高蔬菜抗逆能力,对于盐胁迫下BR对水稻种子萌发的影响研究较少。本研究采用不同浓度BR 对水稻种子进行处理,研究种子萌发过程生理生化变化特征,探讨BR 对水稻种子萌发的影响,为水稻种子萌发理论研究和实际生产提供参考。
1 材料与方法
1.1 材料
供试品种为日本晴,由宁夏农林科学院提供,油菜素内酯由上海威敌生物公司提供。
1.2 方法
1.2.1 试验设计
试验于2018年5月在河套学院农学实验室进行。所用培养皿直径为9 cm,其溶液中含100 mmol/L NaCl 和0、0.1、0.5、1.0、1.5、2.0 mg/L BR(CK、B1、B2、B3、B4、B5)。选择颗粒饱满、尺寸一致的水稻种子,用10%次氯酸钠浸泡10 min;接着用无菌水反复冲洗5次;再用滤纸擦干种子表面水分,将其均匀摆放在铺有2 层滤纸的培养皿中,每处理设置5 次重复,每个培养皿内放30 粒种子;浸种12 h后放置于30 ℃恒温生化培养箱内进行催芽;取其中一部分露白种子进行盆栽试验,试验用土NaCl 含量为0.43%,试验用盆直径为30 cm,高26.5 cm,每个处理4盆,每盆播种5粒,于一心一叶期统计成苗数并计算成苗率。
1.2.2 指标测定
1.2.2.1 发芽率和发芽势的测定
以芽长2 mm 为发芽标准,测定发芽势、发芽率、发芽指数。发芽势=第三天发芽种子数/供试种子总数×100%;发芽率=全部发芽种子数/供试种子总数×100%;种子发芽指数(GI)=Σ(Gt/Dt)(Gt为在t日的发芽种子数,Dt为发芽天数)。
1.2.2.2 胚根细胞膜伤害率的测定
参考李秧秧等[9]的方法,用电导率法测定细胞膜伤害程度,用伤害率(%)表示。
伤害率=(Rd-R0)/(1-R0)×100%
式中,Rd 为受胁迫C1/C2,R0 为未受胁迫C1/C2,C1 和C2 分别为样品受胁迫前后外渗液的电导率值。
1.2.2.3 胚根抗氧化酶活性及丙二醛含量测定
种子萌发7 d 后,取胚根,参考高俊凤[10]的方法分别测定超氧化物歧化酶Superoxide dismutase(SOD)、过氧化物酶Peroxidase(POD)、过氧化氢酶Catalase(CAT)、抗坏血酸过氧化物酶Ascorbate peroxidase(APX) 活 性 和 丙 二 醛Malondialdehyde(MDA)含量。
1.2.2.4 成苗率的测定
于一心一叶期统计成苗数,成苗率=成苗数/播种数量
1.2.3 数据处理
采用Excel 2010 和SPSS 24.0 进行数据整理、分析,Duncan’s 法进行显著性检验;采用Excel 2010作图。
2 结果与分析
2.1 不同浓度BR对盐胁迫下水稻种子萌发的影响
由表1 能够看出,随着BR 浓度的升高,水稻种子发芽率逐渐升高,其中B3 处理发芽率最高,之后随着BR 浓度增加发芽率不再增加,B1、B2、B3、 B4、 B5 分 别 比CK 增 加12.83%、 21.78%、34.62%、32.06%、29.48%。B3、B4、B5 发芽势也随着BR 浓度的升高逐渐升高;B1 和CK 差异不显著,B2、B3、B4、B5 显著高于CK,其中B5 处理发芽势达到最大值,B3、B4、B5 间差异不显著。发芽指数也是随BR 浓度的升高而升高,BR 处理显著高于CK,其中B5 处理时最高,B3、B4、B5 间差异不显著。
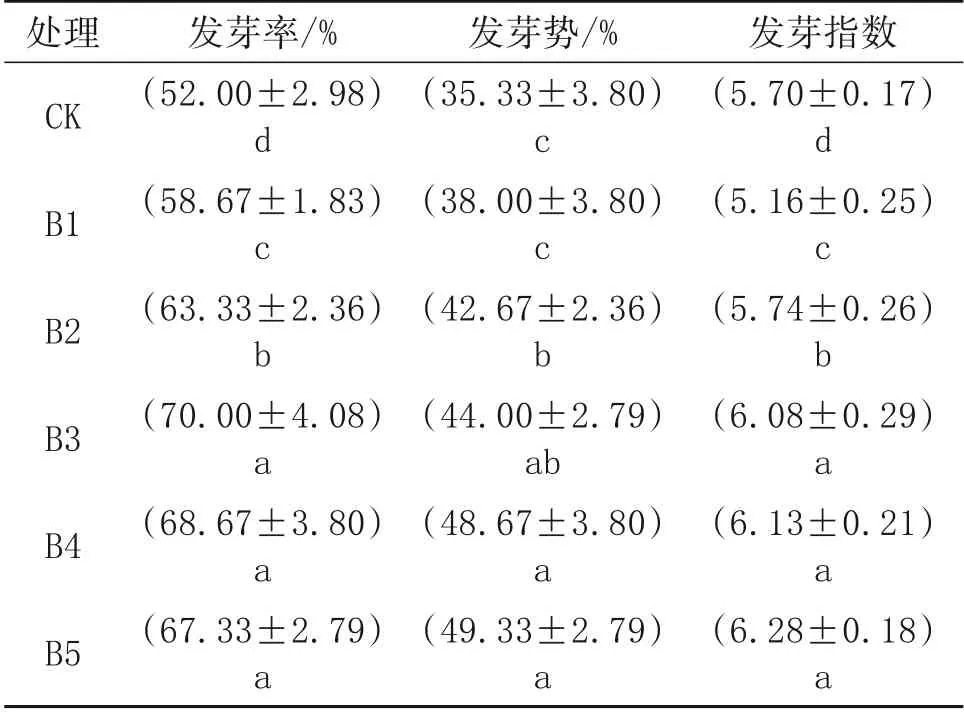
表1 不同浓度BR对盐胁迫下水稻种子萌发的影响
2.2 不同浓度BR 对盐胁迫下水稻胚根伤害率和MDA的影响
伤害率能够直接反映盐胁迫对水稻种子的伤害程度,MDA 是膜脂过氧化的重要产物,能够间接反映膜系统受损程度。由图1可看出,水稻胚根伤害率随着BR浓度的增加呈现先降低后上升的趋势,BR 处理的种子胚根伤害率均低于CK,B3 处理伤害率最低。B1、B2 和B3 伤害率下降速率较快,B4 与B5 差异不显著。B1、B2、B3、B4、B5 分别比CK 低6.20%、19.65%、40.09%、29.59%、26.49%。MDA反映了逆境下植物细胞膜脂过氧化作用和受伤害程度,也可间接反映植物组织抗氧化能力的强弱。结果显示,BR 能够减少MDA 含量,随BR 浓度的增加MDA 含量呈逐渐减少的趋势;B1 处理与CK 差异不显著,其他处理与CK 差异显著;B1、B2、B3、B4、B5 分别比CK 降低了8.31%、22.75%、26.57%、27.55%、30.98%,B2、B3、B4 和B5 间 差 异 不显著。
2.3 不同浓度BR 对盐胁迫下水稻胚根保护酶活的影响
SOD 是清除生物自由基的主要物质,POD 能够将植物体内过氧化氢催化为水和氧气,CAT、APX是清除植物内过氧化氢的关键酶。由图2 可看出,SOD、POD 活性随BR 浓度的升高逐渐升高,各处理均显著高于对照处理;SOD活性在B5处理达到最大值,比CK 高出27.35%;POD 活性也是在B5 处理达到最大值,比CK 高出62.89%。CAT 活性随BR 浓度的升高呈现先升高后降低的趋势,各处理均高于对照处理,B3 处理CAT 活性最大;B1、B2、B3、B4、B5 分 别 比 高 出41.46%、44.30%、57.54%、52.80%、48.64%。APX活性随BR浓度的升高呈现先升高后降低的趋势,B1 处理与CK 差异不显著,其他处理均显著高于CK,B1、B2、B3、B4、B5 分别比 对 照 高17.28%、 39.95%、 55.26%、 51.66%、43.97%。结果表明,高浓度的BR 对SOD、POD 活性具有促进作用;适宜BR 浓度下CAT 和APX 活性较高,但BR 浓度过高时CAT 和APX 活性受到抑制。
2.4 不同浓度BR 对盐胁迫下水稻种子成苗率的影响
由图3 可看出,幼苗成活率随BR 浓度的升高呈先升高后降低的趋势,除了B1,其他各处理均显著高于对照处理。成活率在B3处理达到最大值,比CK高28.78%。
3 讨论
3.1 BR 对盐胁迫下水稻种子发芽率、发芽势和发芽指数的影响
植物生长过程中,Na+和Cl-是必需的营养物质,但植物对Na+和Cl-需要的量极少,过高浓度的Na+和Cl-会对植物体产生离子毒害[11],影响植物的代谢功能,抑制植物细胞内酶的活性,进而影响植物的正常生长发育,植物在遭受盐胁迫后,体内对S 和Mg 的吸收有所增加,抑制了对K+、Ca2+的吸收,引起离子吸收失衡,代谢失调[12]。BR 对逆境有重要的调节作用。种子发芽率、发芽势和发芽指数反映种子的发芽速率和整齐程度。
本试验结果表明,BR 能够显著促进种子萌发,发芽率随着BR 浓度的增加呈现先增加后降低的趋势,发芽势和发芽指数均随着BR 浓度的增加呈现逐渐增加的趋势,发芽率增加了11.36%~25.71%,发芽势增加了7.02%~28.39%,发芽指数增加了8.91%~25.16%,但B3、B4、B5 间差异不显著,说明BR 浓度在达到一定值时对种子萌发的促进作用减缓。吴雪霞等[13]研究表明,外源BR显著缓解盐胁迫对茄子种子的伤害,使种子发芽率提高了8.23%,发芽势提高15.91%,发芽指数提高了17.23%,与本试验研究结果一致,说明BR 可以缓解盐胁迫对水稻种子的伤害,增强水稻种子的耐盐能力。本研究中,低浓度BR 能够提高盐胁迫下水稻种子的发芽率、发芽势和发芽指数,高浓度BR 处理对水稻种子萌发的促进作用不明显。因此,BR 对盐胁迫下水稻种子萌发的促进作用具有浓度效应。
3.2 BR 对盐胁迫下水稻胚根伤害率、MDA 和保护的影响
盐胁迫首先导致细胞膜结构和功能受损,伤害率能够反映胁迫对细胞膜伤害程度[14]。MDA是膜脂过氧化的重要产物,能够间接反映膜系统受损程度以及植物抗逆性[15]。本试验结果表明,水稻种子伤害率随BR 浓度增加呈现先降低后上升的趋势,MDA含量随BR浓度增加逐渐减少,说明一定浓度BR 能够减小盐胁迫对水稻种子的伤害,抑制膜脂过氧化, 防止离子渗漏,使细胞膜维持正常的结构,确保植物体各种代谢循环的正常进行。陆晓民等[16]研究发现,BR 降低了盐胁迫下黄瓜丙二醛含量和细胞膜透性,促进幼苗生长,有效缓解了盐胁迫造成的伤害。
超氧自由基阴离子和过氧化氢同属植物中的活性氧系统[17],植物在遭受逆境胁迫时会产生大量的氧自由基,自由基参与细胞膜脂过氧化。细胞受到逆境胁迫时,酶的活性降低,自由基则会逐渐增加,加速膜脂过氧化反应,破坏膜系统,导致细胞代谢出现异常,对机体造成伤害,从而破坏质膜,最终可能导致细胞死亡[18]。为抵抗逆境对植物造成的伤害,植物体内保护酶系统SOD、POD、CAT和APX会启动,其活性提高,但是随着受胁迫程度增加,保护酶系统清除自由基能力受到限制。外源BR 则能够进一步增强SOD、POD、CAT和APX活性,减少活性氧的产生,进而增强植物耐盐性。本研究结果表明,SOD 和POD 活性随BR 浓度增加而提高,CAT 和APX 活性随BR 浓度增加先提高后降低,可能是在盐胁迫下,SOD 和POD 对BR 的适应性较好,过高浓度的BR 限制了CAT 和APX 活性。朱学玮[19]研究结果表明,在干旱条件下,BR 通过提高小麦和玉米保护酶活性来提高其对干旱的抵抗力,从而提高其产量。王舒甜等[20]研究表明,适宜浓度的BR 可提高盐胁迫下香樟叶片抗氧化能力,清除叶片中的活性氧,保护膜结构,缓解盐胁迫对香樟的伤害,从而提高香樟的抗盐性,与本研究结果一致。本研究中,幼苗的成苗率在B3处理时达到最高,BR 和水稻种子发芽率、胚根伤害率、胚根保护酶活性有显著相关关系,说明油菜素内酯不仅能够影响种子的发生,还有利于幼苗的生长发育。
综上所述,水稻种子发芽率的提高主要是因为BR可以有效提高种子萌发过程中保护酶的活性,减少有害物质的积累,从而减小了盐胁迫对水稻种子萌发的伤害,增加了其发芽率和发芽指数。本试验结果表明,BR 浓度为1.0 mg/L 时,水稻种子发芽率最高、胚根伤害率最低和保护酶活性较高,幼苗成苗率最高。下一步将研究BR 对盐胁迫下水稻种子萌发及植株生长的影响,为今后相关研究提供理论依据。
猜你喜欢
安徽农业科学(2021年14期)2021-09-03
山西农业科学(2021年8期)2021-08-17
心肺血管病杂志(2020年5期)2021-01-14
分析化学(2018年11期)2018-01-16
现代农业研究(2017年3期)2017-04-17
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年11期)2016-01-27
科学中国人(2015年13期)2015-02-28
江苏农业科学(2014年9期)2014-11-15
安徽农学通报(2014年16期)2014-09-18
