
下调miR-122对非酒精性脂肪性肝炎小鼠的炎症发生及肝细胞凋亡的影响
2020-08-26 12:14谢景军魏从光张建峰孙常青王怀志
胃肠病学和肝病学杂志 2020年8期
谢景军,魏从光,张建峰,孙常青,王怀志
武警河南总队医院1.普通外科;2.内二科消化内科病区;3.卫勤处,河南 郑州 450002
非酒精性脂肪性肝病(non-alcoholic fatty liver disease, NAFLD)是由高水平的循环游离脂肪酸(free fatty acids, FFA)引起的病理状况,导致脂质沉积在肝细胞中,从而引发脂肪变性。脂肪变性会发展为非酒精性脂肪性肝炎(non-alcoholic steatohepatitis, NASH)、肝纤维化、肝硬化和肝细胞癌,最终导致肝衰竭。NASH与肥胖症和代谢综合征(特别是糖尿病和全身胰岛素抵抗)密切相关[1-2]。然而,NASH的发病机理和进展尚不完全清楚,需要进一步研究。
最近的报道指出,微小RNA(microRNAs, miRNAs)在调节肝脂肪变性中起关键作用。miRNAs负调控脂代谢中涉及多种蛋白质的表达,因此转录后修饰脂肪分解和脂肪形成[3]。此外,miRNAs从肝细胞中输出,其在血清中的分布与肝病临床前模型中肝脏病理的潜在机制相关[4]。研究发现,miR-122在发育中的肝脏中表达,并在NASH患者血清中高表达[5]。Esau等[6]研究发现,miR-122对脂质代谢具有调节作用,能使高脂喂养的小鼠脂肪变性水平和血浆胆固醇水平显著降低。因此,miR-122可作为诊断和评估组织学病变严重性的一种新的生物学指标,同时有望成为一个潜在的治疗靶点。本研究旨在探讨miR-122在NASH发病机理和进展中的确切作用,寻找新的治疗靶点。
1 材料与方法
1.1 材料
1.1.1 动物:60只C57BL6/J雄性小鼠购自北京维通利华实验动物技术有限公司,体质量(20±2)g。实验动物设计计划严格经过我院伦理委员会审查通过。将实验动物饲养于动物房,温度为(22±2)℃,湿度为(60±5)%,光照/黑暗周期为12 h。适应环境2周,正常饮水饮食。
1.1.2 试剂:MCD饲料购自美国Research Diet公司。AAV2-U6-gRNA-CAG-EGFP-hGH polyA(4.28×1012vg/ml)和AAV2-inhibitor-rno-miR-122-CAG-EGFP(4.10×1012vg/ml)由上海吉凯基因化学技术有限公司合成。Trizol试剂、cDNA逆转录试剂盒和SYBR Premix EX TaqTM试剂盒购自日本TaKaRa公司。HE试剂盒、DAB显色盒购自北京索莱宝生物科技有限公司。油红O染色试剂盒和BCA试剂盒购自美国Sigma公司。TG试剂盒、ALT试剂盒、AST试剂盒、TNF-α试剂盒、IL-1β试剂盒和IL-6试剂盒购自美国Abcam公司。TUNEL试剂盒和RIPA裂解液购自南京凯基生物科技有限公司。兔抗Bax抗体、兔抗Bcl-2抗体、兔抗GAPDH抗体和山羊抗兔IgG二抗购自Cell Signaling Technology公司。
1.2 方法
1.2.1 实验分组及给药:60只C57BL6/J雄性小鼠随机分为4组:control组、MCD组、MCD+miR-NC组和MCD+miR-122 inhibitor组。MCD+miR-NC组小鼠经尾静脉注射AAV2-U6-gRNA-CAG-EGFP-hGH polyA(4.28×1012vg/ml)的腺病毒100 μl,MCD+miR-122 inhibitor组小鼠经尾静脉注射AAV2-inhibitor-rno-miR-122-CAG-EGFP(4.10×1012vg/ml)的腺病毒100 μl,连续5 d;control组和MCD组小鼠通过尾静脉注射相应体积的无菌生理盐水,连续5 d。之后,MCD组、MCD+miR-NC组和MCD+miR-122 inhibitor组小鼠喂养MCD饲料,control组小鼠喂养正常饲料,连续3周,实验过程中无小鼠死亡。实验结束后,造模组小鼠的肝组织进行HE染色,若肝脏组织显示出典型的脂肪性肝炎特征,则造模成功。
1.2.2 qRT-PCR:用Trizol试剂提取肝组织的总RNA。使用紫外分光光度计检测总RNA的纯度和含量,并将每个配对的样品调节至相同浓度。然后按照PrimeScriptTM1st Strand cDNA Synthesis试剂盒将总RNA反转录成cDNA。使用SYBR Premix EX TaqTM试剂盒进行qRT-PCR。使用FTC-3000p实时PCR系统完成实验后,采用2-△△Ct法分析数据。其中使用的引物如表1所示。
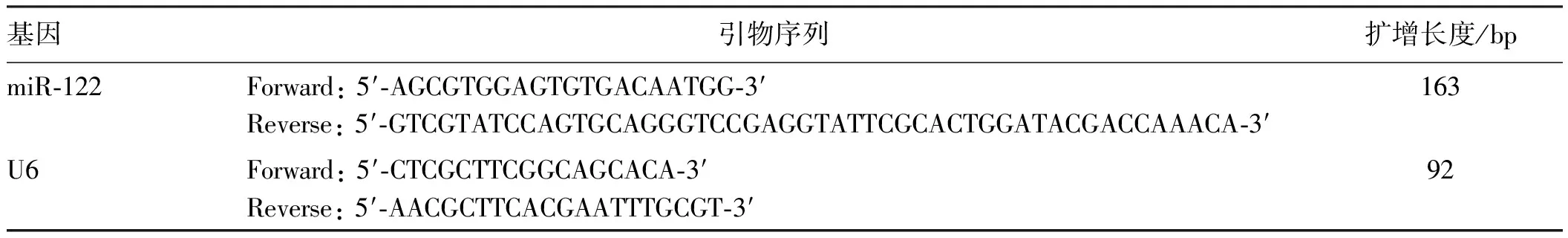
表1 引物序列
1.2.3 HE染色:处死小鼠后切取小块新鲜肝脏组织,经固定、常规脱水、透明、浸蜡、石蜡包埋,并制成5 μm的石蜡切片。切片放入苏木素染液中5 min,自来水清洗后,置于1%的盐酸酒精分化数秒,并用流水冲洗。切片放入伊红染液中染色 3 min,流水稍洗。之后切片置于梯度酒精(70%、80%、90%、100%)中脱水,各5 min,再放入二甲苯Ⅰ、Ⅱ中透明,分别5 min。将切片取出,用滤纸轻轻拭干切片上残余的二甲苯,在组织上滴加中性树胶封片,使用莱卡显微镜进行拍摄。
1.2.4 油红O染色:处死小鼠后,切取小块新鲜的肝脏组织,进行OCT包埋,并制作成6 μm厚的冰冻切片。取各组小鼠肝组织冰冻切片复温1 h后,将切片置于60%异丙醇中浸泡1 min,PBS清洗3次。切片置于油红溶液中染色30 min,37 ℃温浴,PBS清洗3次。苏木素染色30~60 s,1%盐酸酒精分化,1%氨水返蓝。PBS清洗3次后,用滤纸吸干残余水分,用甘油明胶封片,使用莱卡显微镜拍摄照片。
1.2.5 ELISA:按照ELISA试剂盒说明书进行操作,用酶联仪在450 nm波长依序测定各孔的光密度(OD)值,根据OD值所绘制的标准曲线分析血清中TG、ALT、AST、TNF-α、IL-1β和IL-6的表达。
1.2.6 TUNEL染色:取各组小鼠肝脏组织冰冻切片,复温1 h,37 ℃烤箱30 min,之后用PBS清洗3次。切片滴加蛋白酶K工作液(20 μg/ml),37 ℃温育10~30 min,PBS清洗3次。接着切片在4%的多聚甲醛溶液中室温固定5 min,PBS清洗3次。然后切片上滴加DNse I 反应液,37 ℃温育 30 min,PBS清洗3次。用吸水纸小心吸去样本区域周围的多余液体,滴加TdT酶反应液于样本区域上,于37 ℃湿盒中孵育60 min(避光),PBS清洗3次。用吸水纸小心吸去样本周围的液体,每个样本上滴加Streptavidin-Fluorescein标记液,放入温盒中,37 ℃反应30 min(避光),PBS清洗3次。最后DAPI染色液复染细胞核,室温避光反应10 min,PBS洗涤,加适量封片剂,荧光显微镜观察并拍照。
1.2.7 Western blotting检测:各组小鼠的肝组织(100 mg)用RIPA裂解提取总蛋白,并通过BCA方法进行定量,样品经变性后进行上样。按照实验步骤进行电泳-转膜-封闭(质量浓度为50 g/L的脱脂奶粉)-一抗孵育[兔抗Bax抗体(1∶2 000)、兔抗Bcl-2抗体(1∶2 000)和兔抗GAPDH抗体(1∶2 000),4 ℃过夜]-二抗孵育[山羊抗兔IgG抗体(1∶2 000)室温1 h]-显影曝光。Image-Pro Plus图像分析系统对蛋白条带进行分析。

2 结果
2.1 各组小鼠肝组织中miR-122 mRNA的相对表达
qRT-PCR结果表明,与control组比较,MCD组和MCD+miR-NC组小鼠肝组织中miR-122 mRNA的表达水平明显上调(P均<0.01);与MCD组和MCD+miR-NC组相比,MCD+miR-122 inhibitor组小鼠肝组织中miR-122 mRNA的表达水平显著下调(均P<0.001);且MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见表2)。

表2 各组细胞肝组织中miR-122的表达水平
2.2 各组小鼠肝组织病理损伤情况HE染色结果表明,与control组比较,MCD组和MCD+miR-NC组小鼠的肝脏显示出典型的脂肪性肝炎特征,出现中度至重度肝细胞空泡化和明显的肝细胞肥大;与MCD组和MCD+miR-NC组相比,MCD+miR-122 inhibitor组小鼠的肝脏组织表现出轻度至中度的肝细胞空泡,并且细胞肥大程度明显降低(见图1)。

图1 HE染色观察各组小鼠肝组织病理损伤情况(放大200倍)
2.3 各组小鼠肝组织脂肪堆积情况油红O染色表明,与control组比较,MCD组和MCD+miR-NC组肝脏中出现大量的脂质滴(P均<0.001);与MCD组和MCD+miR-NC组相比,MCD+miR-122 inhibitor组肝脏组织中脂质滴减少(P均<0.05);且MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见图2、表3)。
表3 各组小鼠肝组织脂肪堆积情况

图2 油红O染色观察各组小鼠肝组织脂肪堆积情况(放大200倍)
2.4 各组小鼠血清中TG、ALT和AST表达水平ELISA结果表明,与control组比较,MCD组和MCD+miR-NC组小鼠血清中TG、ALT、AST表达水平明显上调(P<0.01);与MCD组和MCD+miR-NC组比较,MCD+miR-122 inhibitor组小鼠血清中TG、ALT和AST表达水平下调(P<0.05);且MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见表4)。
表4 各组小鼠血清中TG、ALT、AST表达水平
2.5 各组小鼠血清中TNF-α、IL-1β和IL-6表达水平ELISA结果显示,与control组比较,MCD组和MCD+miR-NC组小鼠血清中TNF-α、IL-1β和IL-6表达水平明显上调(P<0.01);与MCD组和MCD+miR-NC组比较,MCD+miR-122 inhibitor组小鼠血清中TNF-α、IL-1β和IL-6表达水平下调(P<0.05);且MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见表5)。

表5 各组小鼠血清中TNF-α、IL-1β和IL-6表达水平
2.6 各组小鼠肝细胞凋亡情况TUNEL荧光染色结果表明,与control组比较,MCD组和MCD+miR-NC组肝细胞凋亡指数显著升高(P均<0.001);与MCD组和MCD+miR-NC组比较,MCD+miR-122 inhibitor组肝细胞凋亡指数下降(P均<0.05);MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见图3、表6)。
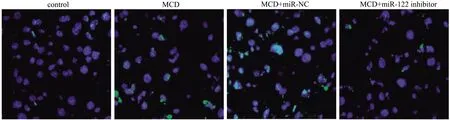
图3 TUNEL染色观察各组小鼠肝细胞凋亡情况(放大400倍)
表6 各组小鼠肝细胞凋亡情况
2.7 各组小鼠肝组织中Bax、Bcl-2蛋白表达水平Western blotting结果表明,与control组比较,MCD组和MCD+miR-NC组小鼠肝组织中Bax蛋白表达水平明显上调(P<0.01),Bcl-2蛋白表达显著下调(P均<0.001);与MCD组和MCD+miR-NC组比较,MCD+miR-122 inhibitor组小鼠肝组织中Bax蛋白表达水平下调(P均<0.05),Bcl-2蛋白表达上调(P均<0.05);且MCD组与MCD+miR-NC组之间差异无统计学意义(P>0.05)(见图4、表7)。

图4 Western blotting检测各组小鼠肝组织中Bax、Bcl-2蛋白表达水平
表7 各组小鼠肝组织中Bax、Bcl-2蛋白表达水平
3 讨论
NASH是最常见的慢性肝病之一。它的范围从肝脂肪变性到消化道脂肪性肝炎和肝纤维化,是多种代谢疾病的风险因素,包括肥胖、2型糖尿病和血脂异常[7]。NASH的主要治疗方法不仅涉及逆转肝细胞TG、ALT或AST的积累,而且还可以有效遏制肝炎,从而防止脂肪变性发展为NASH和肝纤维化[8]。然而,检查药理化合物如何预防NASH综合征并了解NASH发病机理至关重要。
MCD饮食的动物模型比其他饮食模型更好地模仿了人类NASH的病理学发现[9]。与高脂小鼠相比,炎症、纤维化和肝细胞凋亡的发展更为严重和迅速[9]。MCD饮食还可以更好地用于研究与人类NASH发病机理或治疗测试有关的机制。
miRNAs是高度保守的内源性非编码RNA分子,通过与靶mRNA的3′非翻译区(3′UTR)结合而沉默蛋白质翻译[10]。miRNAs的表达水平失调可能会引起肿瘤、代谢性疾病和心血管疾病等。miR-122是miRNAs家族成员之一,可促进胆固醇和脂肪酸合成、抑制胆汁酸和脂肪酸氧化等[11]。因此,miR-122 在肝脏脂质代谢调控中发挥重要作用。
NASH的病理机制复杂,其与肝脂质代谢紊乱、肝脂质过氧化增加等因素相关。研究发现,肝细胞凋亡是促进NASH发生、发展的重要环节[12],肝细胞凋亡也与肝脏炎症、肝脏纤维化的联系非常紧密,其肝脏炎性反应越重,肝细胞凋亡率越高[13]。因此,肝脏的炎性反应和肝细胞在NASH中发挥着至关重要的作用。
本实验研究发现,miR-122在NASH小鼠肝组织中表达上调,与前面的文献报道一致。且低表达miR-122后,NASH小鼠肝组织中miR-122的表达显著下调。HE染色和油红O染色结果表明,NASH小鼠肝脏显示出典型的脂肪性肝炎特征,出现中度至重度肝细胞空泡化、明显的肝细胞肥大和大量脂质滴,而低表达miR-122可发现小鼠的肝脏组织表现出轻度至中度的肝细胞空泡,并且细胞肥大程度明显降低,脂质滴明显减少,提示,低表达miR-122可减轻NASH的发生、发展。接下来我们对小鼠炎症因子和一些相关的酶进行了检测,结果表明,NASH小鼠血清中TG、ALT、AST、TNF-α、IL-1β和IL-6表达水平明显上调,而低表达miR-122可降低TG、ALT、AST、TNF-α、IL-1β和IL-6的含量,因此,低表达miR-122可抑制炎症因子和相关酶的表达,从而改善NASH的症状。最后,我们对肝细胞的凋亡情况进行了研究,发现NASH小鼠肝细胞的凋亡指数明显上升、Bax蛋白表达上调和Bcl-2蛋白表达下调,而低表达miR-122可逆转这一现象,提示,低表达miR-122可抑制肝细胞凋亡。
综上所述,我们的研究揭示了低表达miR-122在小鼠模型中对NASH的减弱作用,从而可能发挥其治疗作用。与这一过程相关的潜在机制可能涉及抑制炎症反应和肝细胞凋亡,以及减少肝实质脂质代谢。提示,miR-122可能是治疗NASH的潜在靶点。
猜你喜欢
浙江大学学报(农业与生命科学版)(2022年4期)2022-09-07
中国典型病例大全(2022年13期)2022-05-10
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
临床肝胆病杂志(2021年6期)2021-12-24
现代临床医学(2021年6期)2021-11-20
昆明医科大学学报(2021年3期)2021-07-22
昆明医科大学学报(2021年1期)2021-02-07
科学与财富(2020年15期)2020-07-04
移动通信(2020年4期)2020-05-07
