
南水北调工程对湖泊湿地土壤呼吸速率及碳排放的影响研究
2020-08-24 14:46陈永金陶宝先刘加珍郭雯雯闫胜文贾一灿尚婉滢吴昊杰
生态科学 2020年4期
陈永金, 陶宝先, 刘加珍, 郭雯雯, 闫胜文, 贾一灿, 尚婉滢, 吴昊杰
南水北调工程对湖泊湿地土壤呼吸速率及碳排放的影响研究
陈永金, 陶宝先, 刘加珍, 郭雯雯, 闫胜文, 贾一灿, 尚婉滢, 吴昊杰
聊城大学环境与规划学院, 山东, 聊城 252059
湿地是地球三大生态系统之一, 其碳汇功能对应对全球气候变化具有举足轻重的作用。为了阐明湖泊湿地土地利用变化条件下不同植物群落土壤呼吸速率变化及其与相关环境因子的关系, 运用LI-8100A开路式土壤碳通量测量系统, 测定了东平湖湿地典型植物群落(香蒲、芦苇、杨树林)以及应季作物的春季、夏季土壤呼吸速率, 并对大气温度、大气相对湿度、光照强度、土壤温度以及土壤水分等环境因子进行了监测。结果表明:东平湖湿地不同植物群落春夏土壤呼吸速率有明显不同, 夏季呼吸速率高于春季; 不同群落之间土壤呼吸速率也有较大差异, 应季作物土壤呼吸速率均值最大, 其次是香蒲群落和芦苇群落, 杨树林群落最小。从整体看, 南水北调东线工程的实施, 扩大了芦苇、香蒲群落和杨树林群落的面积, 减少了农作物的面积, 对减少碳排放起到了积极作用。为增强湖泊湿地的碳汇功能, 减少温室气体的排放, 应进一步增加湿地植物中木本植物的面积, 减少农作物种植面积; 不同土层温度对不同群落土壤呼吸速率的影响不同, 香蒲群落土壤呼吸速率与0—5cm土层温度呈正相关, 芦苇群落土壤呼吸速率与0—5 cm,5—10cm土层温度呈正相关, 杨树林群落土壤呼吸速率与0—5cm, 20—25cm土层温度呈正相关关系。
东平湖; 土壤呼吸; 湿地; 植物群落; 环境因子
0 前言
土壤呼吸是陆地生态系统向大气释放CO2 的主要过程[1], 也是陆生植物固定的CO2返回大气的主要途径[2,3], 土壤呼吸速率的变化会带来大气中CO2浓度的变化[3]。湿地与森林和海洋并称地球上三大生态系统, 湿地面积占全球陆地总面积的6%, 碳储量却占陆地土壤碳总储量的11%[4,5]。因此, 湿地既是陆地上重要的有机碳库, 又是多种温室气体的源和汇, 在全球碳循环中占有重要地位[6]。国内外都十分重视湿地土壤呼吸的研究, 美国科罗拉多高山沼泽湿地土壤呼吸速率为1.2—526 mmol.·CO-2m-2·d-1[7], 弗吉尼亚河流湿地土壤碳通量研究发现, 其呼吸速率为3.1 umol·m-2·s-1 [8]。温度是影响湿地土壤呼吸速率的主要环境因子, 科罗拉多高山沼泽湿地土壤呼吸速率与5 cm土层温度呈极显著相关[7], 太湖湿地土壤呼吸速率与土壤温度呈显著正相关, 与土壤湿度不相关[9], 黄河三角洲湿地土壤呼吸速率日变化与与气温关系密切[10-12]。土壤呼吸速率的影响因素还与植物群落类型有关[13], 天津永定河湿地碱蓬群落土壤呼吸速率与土壤温度和含水率具有显著相关性, 芦苇群落不具有相关性[14]; 青藏高原高寒泥炭型湿地土壤呼吸速率与地上生物量、枯落物累积量呈显著正相关[15]; 人类干扰对湿地土壤呼吸速率有着越来越显著的影响[16,17], 过量的氮元素的输入会干扰湿地森林生态系统的养分循环, 破坏其平衡[18]。不同土地利用方式, 土壤呼吸速率也有差异, 在东北三江平原湿地, 土地开垦为旱田后, 土壤呼吸速率增加, 碳汇功能减弱或丧失[13]。
湖泊湿地是湿地生态系统的重要组成部分, 但有关湖泊湿地土壤呼吸速率的研究却较少, 且集中在西北干旱区或青藏高原区[19-21 ], 位于我国东部季风区的湖泊, 受自然环境变化和人类活动影响深刻的我国东部湖泊湿地土壤呼吸的研究相对较少[22-24]。东平湖是山东省第二大淡水湖, 也是我国南水北调东线工程重要调蓄枢纽。2014年整个东线工程建成并通水, 东平湖水位由44 m以下提高到46 m, 5000 hm2土地被淹没, 或因地下水位太高而无法耕种, 农田变成了芦苇、香蒲湿地或人工杨树林。分析比较不同植物群落土壤呼吸速率特征, 探讨土地利用变化对土壤-大气碳循环的影响, 对区域生态环境保护, 应对全球变化都具有十分重要的意义。
1 研究区概况
东平湖位于黄河下游南岸泰安市东平县西部, 35°30′N—36°20′N, 116°00′E—116°30′E之间, 总面积627 km2。该地区属暖温带大陆性季风气候、四季分明、降水较为充沛。春季(3—5月)受极地大陆性气团控制, 温度回升快, 干燥少雨, 多大风, 常有春旱发生。夏季(6—8月)受海洋季风影响, 高温高湿、炎热多雨, 间有伏旱、内涝、大风及连阴雨等灾害性天气出现。秋季(9—11月)随着北方大陆高气压的重新建立与控制, 气温急速下降, 降水量明显减少, 天气稳定, 常有秋旱发生。冬季(12—2月)因受大陆冬季季风控制, 天气干冷, 雨雪稀少。年平均气温13.6 ℃, 气温年较差4.5 ℃, 无霜期200 d, 年日照时数2401.1 h, 平均风速2.8 m·s-1。东平湖湖区面积418 km2, 年降水量624.4 mm, 湖区平均水深2.5 m, 多年平均水文40.68 m[25]。
东平湖是我国南水北调东线工程的重要调蓄枢纽, 也是山东省西水东调的水源地, 地理位置和生态地位十分重要。东平湖地区属于温带大陆性半湿润季风气候, 四季分明。冬春两季多风少雨雪, 夏季高温多雨。东平湖湿地主要由国家级滨湖湿地公园和稻屯洼国家城市湿地公园组成, 湿地生物多样性高, 有野鸭、脊鸟、令鸟、白鹭、翠鸟和白天鹅等大型鸟类。
2 断面设置与监测方法
2.1 监测断面设置
为了监测东平湖湿地土壤呼吸速率, 在东平滨湖国家湿地公园设置3个监测断面, 分别是(1)老湖断面, 位于东平湖滨老湖镇。该断面以人工杨树林群落(6年龄杨树)和应季农作物为主; (2)展营断面, 位于大清河入东平湖处, 该断面主要有人工杨树林(2年龄稀疏林)和小麦、玉米等应季农作物, 还有稀疏芦苇; (3)稻屯洼断面, 稻屯洼原是东平湖的一部分, 随着水位的下降, 逐渐脱离的东平湖, 成为独立的城市湿地。该断面主要以香蒲和芦苇群落为主。
2.2 监测方法
利用LI-8100A(LI-COR,Lincoln,NE,USA)红外气体分析仪测定土壤呼吸速率, 为了比较生长期不同季节湿地土壤呼吸速率的差异, 呼吸速率监测与于2015年4月(春季)和7月(夏季)进行。土壤呼吸的日动态变化测量在6:00—18:00之间, 每隔1 h测定一次, 每次测量时间为5 min, 同一时段每个观测点重复测定3次, 分析时采用其均值。在测量土壤呼吸时, 利用LI-8100A附带的温湿度计在采样点附近同时测量土壤湿度; 用温度计和干湿度计测定近地面的大气温度和湿度, 用土壤温度计测定不同深度的土壤温度。
3 结果与分析
3.1 不同群落春夏土壤呼吸速率比较
3.1.1 春季
老湖监测断面春季土壤呼吸速率在1.0—2.8 μmol·(m2·s)-1之间(图1a), 杨树林群落土壤呼吸速率最低为1 μmol·(m2·s)-1, 最高为1.5 μmol·(m2·s)-1, 小麦和油菜群落的呼吸速率最低值为1.6 μmol·(m2·s)-1, 反映了不同土地利用方式对碳排放的巨大差异。从呼吸速率的日变化看, 小麦、油菜和杨树林三个群落都呈单峰曲线变化, 其中, 小麦和油菜群落表现出较高的一致性, 但小麦的呼吸速率的最高值出现在11:30, 而油菜的最高值出现在12:00, 最低值都出现在早晨8:00。杨树林群落呼吸速率的最高值出现在14:00, 最低值出现在18:00。

图1 东平湖湿地不同断面不同群落土壤呼吸速率日动态
Figure 1 Diurnal dynamics of soil respiration rate among different plant communities of different sections
展营断面小麦群落土壤呼吸速率从8:00到12:00变化很小, 14:00出现明显增加, 16:00达到最高值。然后逐渐下降, 16:00及其以后的呼吸速率明显高于16:00前。杨树林群落呼吸速率没有显著峰值, 14:00之前呼吸速率处于平稳且较低状态(图1b)。
香蒲与芦苇是稻屯洼湿地最典型的植被, 两群落呼吸速率在1.4—2.87 μmol·(m2·s)-1之间波动, 香蒲与芦苇群落土壤呼吸速率日变化趋势大致相同, 但芦苇群落明显高于香蒲群落。香蒲群落土壤呼吸速率日动态中在18:00达到最大值, 为2.39 μmol·(m2·s)-1; 芦苇群落的土壤呼吸速率最大值为2.87 μmol·(m2·s)-1, 出现在16:00(图1c)。
3.1.2 夏季
老湖断面2种植被7月份土壤呼吸速率日变化整体变化范围为3.9—6.8 μmol·(m2·s)-1之间, 且2种植被土壤呼吸日动态曲线都较为平缓。杨树群落的土壤呼吸曲线在一天之中呈现下降趋势, 最大值在早上8:00出现, 为4.43 μmol·(m2·s)-1, 最小值出现在18:00, 为3.94 μmol·(m2·s)-1。应季作物群落土壤呼吸最大值出现在上午10:00, 为6.81 μmol·(m2·s)-1, 最小值出现在下午18:00, 为5.49 μmol·(m2·s)-1(图1d)。
展营断面杨树群落土壤呼吸速率在2.0—3.0 μmol·(m2·s)-1之间, 远低于5.1—6.0 μmol·(m2·s)-1的玉米群落。杨树群落的土壤呼吸最大值与最小值分别为3.09 μmol·(m2·s)-1和2.41 μmol·(m2·s)-1, 分别出现在14:00和8:00; 而玉米群落的最大值与最小值分别为6.01 μmol·(m2·s)-1和5.35 μmol·(m2·s)-1, 分别出现在14:00和6:00(图1e)。
稻屯洼断面2种植被土壤呼吸速率在4–7 μmol·(m2·s)-1之间波动, 香蒲群落土壤呼吸呈先升后降的单峰曲线, 12:00达到最大值, 为6.88 μmol·(m2·s)-1; 最低值则出现在早上8:00, 为4.54 μmol·(m2·s)-1。芦苇群落的土壤呼吸速率变化也较为平缓, 最大值出现在下午16:00, 为5.52 μmol·(m2·s)-1, 最小值出现在早上8:00, 为4.1 μmol·(m2·s)-1(图1f)。
3.1.3 春夏土壤呼吸速率差异
利用SPSS软件中的方差分析比较不同群落以及同一群落不同季节土壤呼吸速率的差异性。老湖和展营断面春季杨树林群落和油菜、小麦群落的呼吸速率差异显著, 而农作物之间呼吸速率差异性不具有统计显著性(图2a,b); 稻屯洼断面香蒲和芦苇群落土壤呼吸速率差异性显著(图2c); 夏季老湖和展营断面的杨树林和小麦群落呼吸速率差异性显著, 稻屯洼断面香蒲和芦苇群落土壤呼吸速率的差异也达到统计显著性(图2d-f)。
图3是杨树林、芦苇和香蒲群落春夏土壤呼吸速率的比较, 可以看出, 三个群落的春夏土壤呼吸速率差异性都具有统计的显著性。
3.2 环境因子
3.2.1 春季
春季东平湖湿地各监测断面太阳照度日变化显著, 日出和日落时最低, 峰值出现在10:00—14:00。不同断面照度也有差异, 其中展营断面平均照度大于稻屯洼和老湖两个断面。不同植物群落照度也有明显差异, 老湖断面小麦和油菜群落照度是杨树林群落的2倍多, 展营断面的芦苇群落照度也差不多高出杨树林近1倍, 稻屯洼断面香蒲和芦苇群落照度则比较接近。相同植物群落在不同断面的照度也有非常显著的差异, 展营断面的杨树林群落照度远高于老湖断面, 稻屯洼断面芦苇的照度也低于展营断面, 这是因为展营断面杨树林是刚栽种不久的稀疏幼林, 而芦苇也长得比较稀疏, 因此, 照度最高。的相比较太阳照度, 相同断面不同群落之间气温和大气相对湿度相差较小(图4a-c)。
东平湖湿地气温的日变化比照度要平缓, 而且同一断面不同植物群落间气温差异不是很大, 但不同断面气温还是有差异的(图4d-f)。大气相对湿度的变化与气温呈反向变化(图4g-i)。
土壤温度表现出明显的分层结构, 其中0-5cm表层土壤温度日变化最大, 平均温度也最高, 越下下层温度越低, 变化幅度也越小(图5)。
3.2.2 夏季环境因子分析
相对于呼吸速率, 环境因子在不同断面和不同群落之间的差异性较小, 我们以展营断面为例, 分析照度、气温和大气相对湿度以及土壤温度的日变化特征。展营断面杨树林和玉米群落照度都呈单峰曲线变化, 最高值为52533 Lx, 出现在杨树林群落, 但照度均值则是玉米群落大于杨树林群落, 另外, 杨树林和玉米群落照度峰值出现的时间也不同, 杨树林为14:00, 玉米群落出现在12:00(图6)。两群落气温变化趋势大体相同, 都呈单峰曲线, 最高值都出现在14:00, 玉米群落平均气温高于杨树林群落。大气相对湿度变化与气温呈反向变化, 杨树林群落高于玉米群落。

图2 不同群落土壤呼吸速率的差异性
Figure 2 Difference of soil respiration rates between different communities

图3 典型群落春夏土壤呼吸速率对比
Figure 3 Comparison of soil respiration rates of typical communities between Spring and Summer
我们以稻屯洼断面为例, 比较环境因子的春夏变化。从图可以看出, 香蒲和芦苇群落照度和大气温度都是夏季高于春季, 而大气相对湿度则是春季高于夏季(图7a-c)。夏季气温高于春季与太阳高度角有关, 大气相对湿度夏季高于春季则是由于大气中水汽含量相对固定, 而夏季气温高, 饱和水汽压大, 因此, 相对湿度出现下降变化。
图8是稻屯洼断面香蒲和芦苇群落春夏土壤平均气温的变化, 从整体看, 春夏土壤平均温度变化趋势基本一致, 都是早晨6: 00温度最低, 然后逐渐升高, 在16: 00达到最高温度后逐渐下降。具体群落来说, 春季芦苇群落土壤平均温度略高于香蒲群落, 而夏季香蒲群落土壤均温又稍高于芦苇。香蒲群落春夏平均温差在8.7℃, 而芦苇群落春夏温差为7.1℃。
3.3 呼吸速率与环境因子关系
对土壤呼吸速率与大气温度、大气湿度、照度、土壤水分、土壤温度进行回归分析, 结果如表1、表2:
春季东平湖湿地土壤呼吸与大气温度的关系呈指数型变化, 夏季呈二次线性关系。4月份四种植被相关系数2按从大到小的顺序排列, 依次是:香蒲(0.854)>芦苇(0.85)>小麦(0.793)>杨树(0.319); 7月份三种植被相关系数R2按从大到小的顺序排列, 依次是:芦苇(0.81)>香蒲(0.555)>杨树(0.36)。
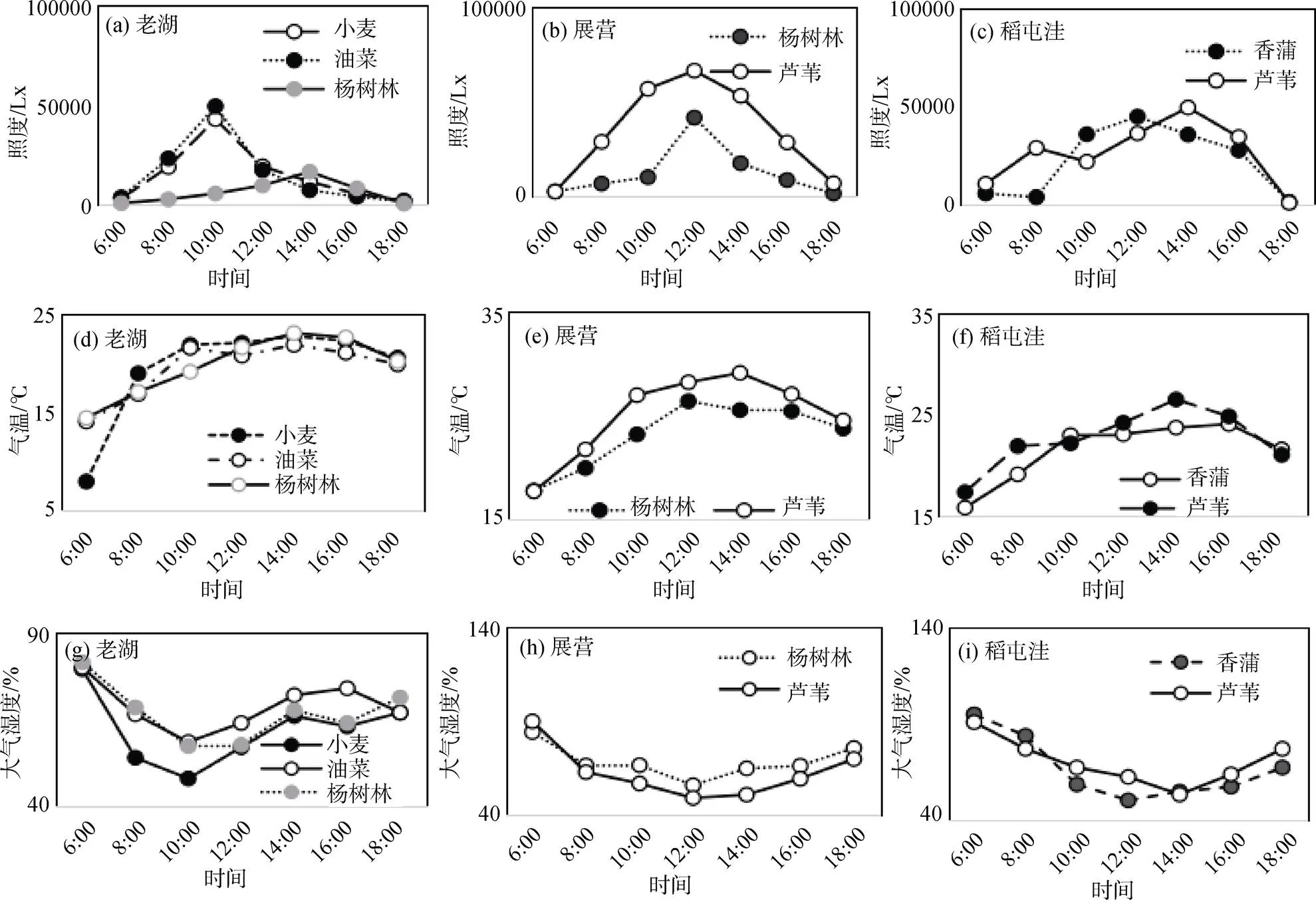
图4 春季环境因子日动态
Figure 4 Diurnal dynamics of environmental factors in Spring
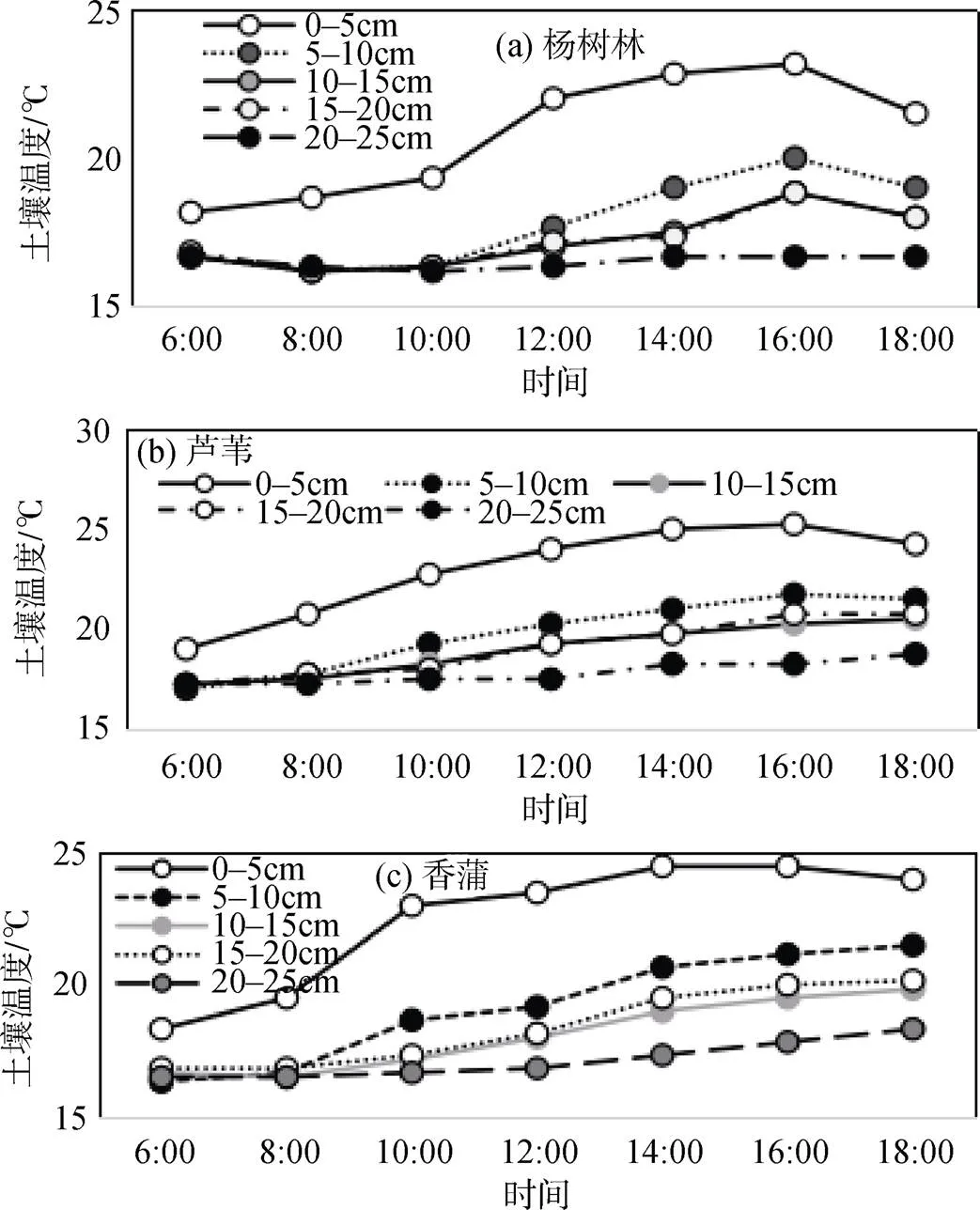
图5 典型群落春季土壤温度日动态
Figure 5 Diurnal dynamics of soil temperature of typical communities in Spring

图6 展营断面夏季环境因子日动态
Figure 6 Daily dynamics of environmental factors of Zhanying Section in Summer

图7 香蒲、芦苇群落春夏环境因子比较
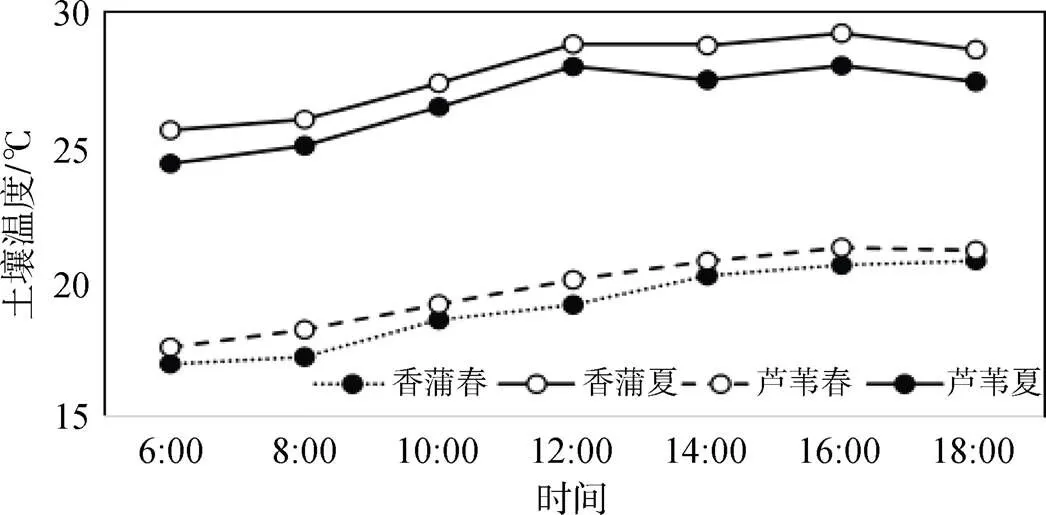
图8 稻屯洼断面香蒲和芦苇群落土壤剖面春夏平均温度日变化
Figure 8 Diurnal variation of average temperature in soil profile ofand reed communities at Daotunwa Section in Spring and Summer
大气湿度是与温度息息相关的环境因子, 根据4月与7月份与土壤呼吸的相关性曲线分析, 4月份相关系数R2按从大到小排序为:芦苇(0.866)>香蒲(0.716)>杨树(0.531)>小麦(0.432), 7月份 R2按从大到小排序为: 芦苇(0.805)>杨树(0.552)>香蒲(0.504)。该结论与大气温度相似, 芦苇群落与大气相对湿度的相关性高于香蒲、杨树林和小麦群落。不同植物群落由于受气象因子影响, 会有相当大的差异。近地面大气湿度会影响土壤微生物, 植物根系等呼吸作用的进行, 从而影响土壤呼吸速率。

表1 春季3种植物群落与各环境因子拟合曲线
植物群落与照度呈现较平缓的二次多项式关系, 此规律在4月份与7月份相一致。随着照度的变化, 各植物群落土壤呼吸速率变化趋势略有不同, 其中杨树群落的相关性最高, 相关系数最大(R2=0.738), 其次为小麦(0.445), 芦苇(0.404), 相关性最小的则是香蒲(0.317)。7月份, 香蒲群落与土壤呼吸的相关性是最高的, 相关系数为0.852, 其次为芦苇(0.612), 杨树最小(0.499), 这种现象有可能由于是季节变化, 大气湿度与温度均发生不同程度的改变, 各种环境因子发生微妙变化, 共同作用所引起的。

表2 夏季3种植被群落与各环境因子拟合曲线
春夏土壤呼吸速率与土壤水分拟合方程显示, 除香蒲的相关系数为0.9以外, 其余的相关系数均在0.1—0.3之间, 表明土壤呼吸速率与土壤水分的相关性并不高, 即土壤含水量不是影响土壤呼吸速率的限制性因子。
土壤呼吸与春夏土壤温度有很高的相关性, 而且同一植物群落土壤呼吸与春夏土壤温度的相关程度也有所差异, 例如春季杨树群落两种关系的相关系数为0.43, 明显小于夏季的0.74, 其他植物群落也是如此。春季, 不同植物群落土壤呼吸与土壤温度的相关性大小顺序为: 香蒲(0.82)>芦苇(0.8)>杨树(0.434)。详细分析, 杨树在10—15 cm处土壤呼吸与土壤温度的相关性最高(0.64), 香蒲则是在5—10 cm处最高(0.94), 而芦苇是在20—25 cm处相关性最高(0.9); 夏季3种植物群落的土壤呼吸速率与土壤温度的相关性都在0—5 cm处最高, 相关性依次为为杨树(0.95), 芦苇(0.86), 香蒲(0.77), 越往深处, 会有一些减小或增大的微小波动。整体的相关性从大到小依次为杨树(0.74)>芦苇(0.67)>香蒲(0.434), 与春季正好相反, 分析其可能原因为, 各种植物群落土壤呼吸与土壤温度的相关性对于季节的敏感性很高。
4 讨论
(1)东平湖湿地春夏土壤呼吸速率有显著差异, 夏季土壤呼吸速率比春季高1倍多。这与多数研究结论相似[26-29]。夏季植物生长到达旺季, 水热条件逐渐到达最佳状态, 植物根系呼吸与土壤微生物活动强烈, 使得呼吸速率达到最高水平[29-32]。本研究发现, 同一季节农作物群落的土壤呼吸速率高于杨树林、芦苇和香蒲群落, 杨树林群落的土壤呼吸速率低于自然湿地芦苇和香蒲群落。有学者在新疆艾比湖湿地的研究发现, 作为草本植物的芦苇群落土壤呼吸速率大于木本植物柽柳群落[19, 33], 与本研究结论具有一致性。土壤呼吸是土壤有机碳转化为无机碳的重要过程, 土壤有机碳库是土壤微生物分解的主要碳源, 其含量高低直接影响着土壤微生物的活性, 从而影响土壤碳排放[34]; Wagner等研究表明, 土壤CO2排放强度主要取决于土壤中碳氮底物的含量及矿化速率[35]; 还有研究认为, 土壤有机碳及生物量是造成土壤CO2排放差异的直接原因[20]。而在东平湖湿地杨树林, 芦苇和香蒲群落中,杨树林群落的土壤有机质明最高[22], 说明土壤有机质含量以及碳氮多少并不是唯一决定土壤呼吸速率的因素。经过进一步对比, 我们发现, 不论春季还是夏季, 都是杨树林群落的土壤温度和大气温度最低, 或许气温对土壤呼吸速率的影响很重要。当然, 杨树林是紧挨湖边的, 地下水较高, 长期淹水也是抑制土壤呼吸速率的重要因素。土壤呼吸速率高低是土壤碳库中各种形式的碳分解速率的直接体现, 农田生态系统由于每年有大量有机质及养分的输入, 其呼吸速率自然最高。芦苇、香蒲是湖泊湿地上自然生长的植物, 其较高的呼吸速率对湿地的碳源/汇功能有很大影响。在本研究中芦苇、香蒲群落呼吸速率显著高于杨树林群落, 这是由于芦苇香蒲都是一年生植物, 其生长迅速, 但分解也快。而杨树林作为木本植物, 其吸收的物质被转化成植物体的一部分固定起来了, 只有很少一部分以枯枝落叶形式返回土壤中参与碳循环。从整体看, 南水北调工程的实施, 增加了东平湖湿地香蒲芦苇和杨树林群落的面积, 减少了农作物的种植面积, 从碳排放的角度看, 减少了CO2排放, 对应对全球气候变暖起到了积极作用。从不同植物群落呼吸速率看, 广植木本植物是维护湖泊湿地土壤碳汇功能, 进一步减少温室气体排放的重要途径之一。
(2)东平湖湿地土壤呼吸呼吸速率日动态随季节与植物群落变化而有显著变化。香蒲群落春夏季节土壤呼吸速率日动态都表现为明显单峰曲线; 芦苇群落春季土壤呼吸速率日动态表现为非对称性单峰曲线, 夏季则接近双峰曲线; 杨树林群落春季和夏季土壤呼吸速率最高值和最低值之差都只有0.3 μmol·(m2·s), 峰谷变化很小。展营断面, 夏季杨树林群落土壤呼吸速率日动态表现呈从早到晚几乎呈单调下降变化, 与前人在同一研究区研究结果有一定相似性[23],国内外其他地区的研究也有类似的结论[8,35-37]。温度是影响土壤呼吸强度最主要的环境因子之一, 并对土壤中植物根系生长、微生物代谢以及有机质的分级来调控土壤呼吸[36-39], 至于会出现某些群落下土壤呼吸速率不随气温变化发生显著变化的的原因, 有研究认为可能是由于土壤异养微生物竞争不充足的土壤有机质基质来源的结果[38]; 还有研究认为, 是由于土壤活性炭对温度的变化比较敏感, 而惰性碳部分对温度的变化不敏感[39]。
(3)土壤温度与土壤呼吸速率关系密切, 东平湖湿地不同植物群落土壤呼吸速率与不同土层温度表现出不同的关系: 香蒲群落土壤呼吸速率变化主要受0—5 cm土层温度变化的影响, 随着该层温度的升降而增减; 芦苇群落土壤呼吸速率变化与0—5和5—10 cm土层温度变化高度同步性, 人工杨树林群落土壤呼吸速率与0—5, 5—10, 以及20—25 cm土层温度同步变化。国内外有关土壤呼吸速率与不同深度土层关系的研究也较多: 美国科罗拉多高山沼泽湿地土壤呼吸速率与5 cm土层温度呈极显著相关[7]; 在淮南的大通湿地深度土壤温度是芦苇群落土壤呼吸强度主要受10 cm土层温度变化的影响[39]; 在崇明岛湿地, 林带土壤呼吸速率与土壤5 cm处温度的相关性最高[41]; 博斯腾湖芦苇湿地, 近地面温度和5 m土壤温度与CO2排放通量呈现极显著的正相关关系[20]; 安落叶松沼泽与5—15 cm土壤温度存在极显著相关性[26,29,42]。土壤呼吸主要是植物根部以及与根部共生的微生物的呼吸, 植物根系分布深度决定了土壤呼吸速率与不同土层温度的关系。香蒲是多年生草本植物, 其根系较浅, 故土壤呼吸速率仅决定于表层土壤温度; 芦苇也是多年生草本植物, 其根状茎分布深度大于香蒲, 因此, 土壤呼吸速率与0—5 cm和5—10 cm土层温度同步变化; 而杨树作为高大乔木, 其根系分布较深, 因此其呼吸速率与最深土层的关系密切。
5 结论
(1) 东平湖湿地不同植物群落土壤呼吸速率差异显著, 人工杨树林群落呼吸速率低于农作物和芦苇、香蒲群落的土壤呼吸速率。南水北调东线工程的实施, 增加了东平湖湿地整体面积, 芦苇香蒲群落和杨树林群落面积增加, 农田面积减少, 整体上减少了CO2的排放, 进一步增加木本植物的面积是维护湖泊湿地碳汇功能, 缓解全球温室效应的途径之一;
(2) 相同植物群落春夏呼吸速率差异明显, 香蒲群落土壤呼吸速率春夏日变化都呈明显单峰曲线变化, 芦苇群落夏季单峰曲线不明显, 杨树群落春夏日变化都很小, 展营断面杨树林呈单调小幅增加变化, 而老湖断面杨树林群落夏季土壤呼吸速率呈单调下降变化;
(3) 土壤呼吸速率与温度关系密切, 杨树林群落土壤呼吸速率与15—20 cm深度土层温度呈极显著正相关, 芦苇与5—10 cm土层温度呈极显著正相关, 香蒲群落与0—5 cm土层温度呈极显著正相关。由此可见, 植物根系分布深度决定了呼吸速率与不同土层温度的关系。
[1] SCHIMEL D, STEPHENS B B, FISHER J B. Effect of increasing CO2on the terrestrial carbon cycle[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(2): 436–441.
[2] HASIBEDER R, FUCHSLUEGER L, RICHTER A,et al. Summer drought alters carbon allocation to roots and root respiration in mountain grassland[J]. New Phytologist, 2015, 205(3): 1117–1127.
[3] ZHOU Z , CHAO G , HE M , et al. Temperature Sensitivity and Basal Rate of Soil Respiration and Their Determinants in Temperate Forests of North China[J]. PLos One, 2013, 8(12): 1–9.
[4] MATTHERWS E, FUNG I. Methane emission from natural wetlands: Global distribution, area and environmental characteristics of sources[J]. Global Biogeochemical Cycles, 1987, 1(1): 61–86.
[5] FRANZEN L G. Can earth afford to lose the wetlands in the battle against the increasing greenhouse effect[C]// International Peat society Proceedings of International Peat Congress. Uppsala, 1992: 1–18.
[6] 朱慧森, 李刚, 董宽虎, 等. 放牧对賴草草地土壤呼吸日、季动态的影响[J]. 植物学报, 2015, 50(5): 605–613.
[7] Wichland K P, Striegl R G, Mast M A, et al. Carbon gas exchange at a southern Rocky Mountain wetland, 1996-1998[J]. Global Biogeochemical Cycles, 2001, 15(2): 321–335.
[8] BATSON J, NOE G B, HUPP C R, et al. Soil greenhouse gas emissions and carbon budgeting in a short-hydroperiod floodplain wetland[J]. Journal of Geophysical Research: Biogeosciences, 2015, 120(1): 77–95.
[9] 冯育青. 太湖湖滨带土壤呼吸与微生物生物量研究[D]. 南京:南京林业大学, 2010.
[10] 王玲玲, 孙志高, 牟晓杰, 等. 黄河口滨岸潮滩湿地CO2、CH4和N2O通量特征初步研究[J]. 草业学报. 2011, 20(3): 51–61.
[11] 陈亮, 刘子亭, 韩广轩, 等. 环境因子和生物因子对黄河三角洲滨海湿地土壤呼吸的影响[J]. 应用生态学报. 2016, 27(6): 1795–1803.
[12] LUO X X, XING Z Q. Comparative study on characteristics and influencing factors of soil respiration of reed wetlands in Yellow River Estuary and Liaohe River Estuary[J]. Procedia Environmental Sciences, 2010, 2: 888–895.
[13] 王毅勇, 宋长春, 闫百兴, 王德宣, 赵志春, 娄彦景, 宋新山. 等. 三江平原不同利用方式下湿地土壤CO2通量研究[J]. 湿地科学. 2003, 1(2): 111–114.
[14] ZHANG Q, MENG W Q, LI H Y. Research of the diurnal soil respiration dynamic in two typical vegetation communities in Tianjing estuarine wetland [C]//IOP Conference Series: Earth and Environmental Science. IOP Publishing, 2016, 41(1): 12–16.
[15] 汪浩, 于凌飞, 陈立同, 等. 青藏高原海北高寒湿地土壤呼吸对水位降低和氮添加的响应[J]. 植物生态学报, 2014, 38(6): 619–625.
[16] 朱敏, 张振华, 于君宝, 等. 氮沉降对黄河三角洲芦苇湿地土壤呼吸的影响[J]. 植物生态学报,2013, 37 (6): 517–529.
[17] 杨利琼, 韩广轩, 于君宝, 等. 黄河三角洲芦苇湿地生长季净生态系统CO2交换及其环境调控机制[J]. 应用生态学报. 2013, 24(9): 2415–2422.
[18] 张徐源, 闫文德, 郑威, 赵亮生. 氮沉降对湿地松林土壤呼吸的影响[J]. 中国农业通报. 2012, 28 (22): 5–10.
[19] 赵明亮. 艾比湖高盐湖泊湿地典型植物群落下土壤呼出版地吸规律研究[D]. 乌鲁木齐: 新疆师范大学, 2017.
[20] 王金龙, 李艳红, 李发东. 博斯腾湖人工和天然芦苇湿地土壤CO2、CH4和N2O排放通量[J]. 生态学报, 2018, 38(2): 668–677.
[21] 杨龙. 温度和水分对青海湖湿地不同植物群落土壤呼吸的影响-以小湖泊为例[D]. 西宁: 青海师范大学, 2012.
[22] 路亚坤, 刘加珍, 陈永金, 路明, 肖兆伟, 东平湖湖滨带不同植被类型下春季土壤CO2通量研究[J]. 中国农学通报, 2012, 28(28): 8–14.
[23] 路亚坤刘加珍陈永金, 等. 东平湖湖滨带农田土壤碳通量的动态变化[J]. 贵州农业科学, 2013, 5: 104–108.
[24] 路亚坤, 刘加珍, 陈永金, 等. 温带湖泊周边湿地原生草地与人工林土壤碳释放差异性分析[J]. 生态环境学报, 2013, 4: 605–610.
[25] 靖淑慧, 刘加珍, 陈永金, 等. 氢氧稳定同位素对东平湖枯水期水环境的指示作用[J]. 南水北调与水利科技, 2019, 17(1): 120–129
[26] BRIX H. Gas exchange through the soil-atmosphere in terphase and through dead culms of Phragmites australis in a constructed reed bed receiving domestic sewage[J]. Water Research, 1990, 24(2): 259–266.
[27] SONG C C, WANG Y Y, WANG Y S, et al. Character of the greenhouse gas emission in the freshwater mire under human activities[J]. Scientia Geographica Sinica, 2006, 26(1): 82–86.
[28] BUBIER J L, GAYTRI B, MOOR T R, et al. Spatial and temporal variability in growing-season net ecosystem carbon dioxide exchange at a large peatland in Ontario, Canada[J]. Ecosystem, 2003, 6(1): 353–367.
[29] YANG J S, LI J S, WANG J D, et al. Emissions of CH4emissions from wetland soils in the Sanjiang plain[J]. Journal of Agro-Environment Science, 2004, 23(5): 846–851.
[30] MU C C, SHI L Y, SUN X X. Fluxes and controls of CO2, CH4 AND N2O in amarsh wetland of Xiaocing’an mountains, northeastern China[J]. Chinese Journal of Plant Ecology. 2009, 33(3): 617–623.
[31] MU C C, CHENG W, SUN X X. et al. Seasonal variation of emission fluxes of CO2, N2O and CH4from Larix gemlinii wamps Soils in Xiaoxing’an Mountains of China[J]. Scientia Silvae Sinicae, 2010, 46(7): 7–15.
[32] 宋长春, 阎百兴, 王跃思, 王毅勇, 娄彦景, 赵志春. 三江平原沼泽湿地CO2和CH4通量及影响因子[J]. 科学通报, 2003, 48(24): 2749–2753.
[33] 杨建军, 吕光辉, 张燕, 塔西甫拉提·特依拜. 艾比湖流域不同植物群落土壤呼吸研究[J]. 环境科学研究, 2009, 22(3): 362–370.
[34] WAN SQ, NORBY R J, LEDFORD J, WELTZIN J F 2007. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland[J]. Global Change Biology, 13, 2411–2424.
[35] WAGNER D, PFEIFFER E M. Two temperature optima of methane production in a typical soil of the Elbe river marshland[J]. FEMS Microbiology Ecology, 1997, 22(2): 145–153.
[36] 杨晶, 黄建辉, 詹学明, 等. 农牧交错区不同植物群落土壤呼吸的日动态观测与测定方法比较[J]. 植物生态学报, 1994, 28(3): 318–325.
[37] 唐燕飞、王国兵, 阮宏华. 土壤呼吸对温度的敏感性研究综述[J]. 南京林业大学学报(自然科学版), 2008, 32(1): 124–128.
[38]GIARDINA, C P, RYAN M G. Evidence that decomposition rates of organic carbon in mineral soil to not vary with temperature [J]. Nature, 2000. 404: 858–861.
[39] THORNLEY J H M, CANNELL M G R. Soil carbon storage response to temperature: a hypothesis [J], Annual Botany, 2001, 87: 591–598.
[40] 赵魁, 姚多喜, 张治国, 杨清, 安世凯, 张文影. 大通芦苇生态湿地土壤呼吸特征及其影响因子[J]. 中国农学通报, 2013, 29(11): 126–131.
[41] 任文玲, 侯颖, 杨淑慧, 等. 崇明岛新围垦区不同土地利用条件下的土壤呼吸研究[J]. 生态环境学报, 2011, 29(1): 97–101.
[42] 刘霞, 胡海清, 李为海, 孙成坤, 黄超等, 寒温带岛状林沼泽土壤呼吸速率和季节变化[J]. 生态学报, 2014, 4(24): 7356–7364.
Analysis on effects of South-to-North Water Transfer Project on soil respiration rate and carbon emission in lacustrine wetlands
Chen Yongjin, TAO Baoxian, Liu Jiazhen, GUO Wenwen, YAN Shengwen, JIA Yican, SHANG Wanying, WU Haojie
School of Environment and Planning, Liaocheng University, Liaocheng, 252059, China
Wetland is one of the three major ecosystems on the Earth, and its carbon sink function plays an important role in global climate change. In order to clarify the change of soil respiration rate of different plant communities in the wetland of lake and its relationship with related environmental factors, the LI-8100A open-circuit soil carbon flux measurement system was used to determine typical plant communities (,and) in the wetland of Dongping Lake. And the environmental factors were monitored ad collected.The results indicated that the soil respiration rates of different plant communities were significantly different in spring and summer, and higher in summer than in spring. There were also significant differences in soil respiration rates among different communities. The mean values of soil respiration rates of crops in season were the highest, followed byandcommunities, and the lowest incommunities. Overall, the implementation of the East Route Project of South-to-North Water Transfer has expanded the area of,andcommunities, and reduced the area of crops, playing a positive role in reducing carbon emissions. In order to enhance the carbon sink function of lake wetlands and reduce greenhouse gas emissions, the area of woody plants in wetlands should be further increased and crop planting area should be reduced. Temperature exerted different effects on soil respiration rate of different communities. Soil respiration rate ofcommunity was positively correlated with soil temperature of 0-5 cm layer, and that ofcommunity was positively correlated with soil temperature of both0-5cm and 5-10 cm layer, and that ofcommunity was positively correlated with the temperature of 0-5 cm and 20-25 cm layer.
Dongping Lake; soil respiration; wetland, plant community; environmental factor.
10.14108/j.cnki.1008-8873.2020.04.026
陈永金, 陶宝先, 刘加珍, 等. 南水北调工程对湖泊湿地土壤呼吸速率及碳排放的影响研究[J]. 生态科学, 2020, 39(4): 207–217.
Chen Yongjin, Zhang Baohua, Liu Jiazhen, et al. Analysis on effects of South-to-North Water Transfer Project on soil respiration rate and carbon emission in lacustrine wetlands[J]. Ecological Science, 2020, 39(4): 207–217.
Q148
A
1008-8873(2020)04-207-11
2019-12-20;
2020-06-06
国家自然科学基金(40901276, 40871239);国家科技支撑计划项目(2014BAC15B02);聊城大学社科平台项目(321021915,32102916)资助
陈永金(1968—), 男, 山东临沂人, 博士, 副教授, 主要从事湿地生态水文过程研究, E-mail:chenyongjin@lcu.edu.cn
刘加珍, 主要从事生态恢复与保护研究, Email: liujiazhen@lcu.edu.cn
猜你喜欢
——东平湖增殖放流活动实施
科学养鱼(2021年5期)2021-11-30
皮革制作与环保科技(2021年14期)2021-11-12
皮革制作与环保科技(2021年8期)2021-11-11
开卷有益·求医问药(2021年8期)2021-08-24
家教世界·创新阅读(2021年8期)2021-08-23
东坡赤壁诗词(2021年1期)2021-03-24
鸭绿江(2020年15期)2020-11-12
核桃源(2019年3期)2019-11-14
自然保护地(2019年1期)2019-03-18
前卫文学(2017年5期)2017-11-09
