
DNA条码技术在上海辰山植物园10种灯诱夜蛾科昆虫种类鉴定中的应用
2020-08-19 11:33陈东旭上海市绿化管理指导站上海市黄浦区200020上海城市树木生态应用工程技术研究中心上海市黄浦区200020
上海农业科技 2020年4期
陈东旭 (上海市绿化管理指导站,上海市黄浦区 200020;上海城市树木生态应用工程技术研究中心,上海市黄浦区 200020)
防治有害生物是园林植物养护过程中的一个重要环节,防治效果的好坏直接影响园林景观和植物长势。有效地预测预报园林植物病虫害发生情况,可以较好地为养护人员提供预警信息,以便园林养护人员及时对病虫害进行人为干预,降低病虫害对景观的影响,从而达到病虫害绿色防控的目的。在病虫预测预报工作中常使用自动虫情测报灯(自动虫情测报灯可以自动启动关闭,转换接虫器,自动排出雨水,且每天诱捕到的标本被装入一个幼虫袋,测报员一周进行一次收虫工作即可,可大大减少工作量)进行虫害监测,其原理主要是利用黑光灯诱集鳞翅目、鞘翅目等有趋光性的成虫[1-2]。但在害虫测报与综合治理工作中,测报员主要通过捕捉到昆虫的翅膀纹路和病虫图谱进行初步比对,缺少了对雄性外生殖器及外部形态学特征进行分类这一专业环节,鉴定结果的准确度难以保证[3-4];同时,自动虫情测报灯收集到的很多鳞翅目、鞘翅目昆虫的形态学特征相似,标本局部损坏易导致成虫区分特征不明显,这也为昆虫种类鉴定带来了很大难度。而利用DNA条码技术作为昆虫种类鉴定的辅助手段,可提高昆虫种类鉴定结果的准确度[5-7]。
DNA条码技术的原理是提取样本基因组DNA,扩增其线粒体细胞色素C氧化酶亚基I(COI)基因序列片段并在数据库内进行比对分析,进而对物种进行快速、准确的鉴定和分类[8]。目前,基于COI的DNA条形码辅助物种分类技术已大量应用于害虫的测报工作,对鳞翅目昆虫分类鉴定的案例也已有很多报道[9-10]。但是,鉴于鳞翅目是昆虫纲中第2个大目,已知种类超过20万种,夜蛾科又是鳞翅目中最大的一科,具有2万多个种类,且其中有很多种类是为害农作物的种类,也是林业生产中非常重要的害虫[7]。故笔者拟对DNA条码技术在鳞翅目夜蛾科昆虫种类鉴定中的应用效果进行检验,以期建立快速准确的夜蛾科昆虫鉴定方法,从而为夜蛾科昆虫生态多样性评估及综合治理提供重要途径。
1 材料与方法
1.1 样本采集及形态鉴定
本研究选取的样本为夜蛾科昆虫标本。供试昆虫于2016年5月至2017年9月采自上海辰山植物园(植物园内生物多样性丰富,能为动植物群落种类、生态评估、水体等方面的研究提供样本)的三座自动虫情测报灯,分别位于园内北、南、东三个方向。设备均为佳多牌自动虫情测报灯,型号为JDB1-Ⅲ型,黑光灯管主波长为365 nm,诱虫半径为120 m。样本收集后分类整理,选取肢体较为完整的43个样本进行展翅,并保存于上海市绿化管理指导站实验室,待形态学鉴定。
1.2 基因组DNA提取、PCR扩增和测序
基因组DNA提取:研磨43个样本的昆虫腿部肌肉组织,提取基因组DNA,然后置于-20 ℃冰箱保存备用。
PCR扩增:合成鳞翅目COI基因通用引物,PCR扩增所用引物为LCO1490(5’-GGTCAACAAATCATAAAGATATTGG-3’)和HCO2198(5’-TAAACTTCAGGGTGACCAAAAAATCA-3’)。EX-Taq DNA聚合酶、dNTP等PCR试剂均购置于TakaRa公司。
PCR反应体系(50μL) : 10倍PCR缓冲液5 μL,正向引物(20μmol/L)2.5μL,反向引物(20μmol/L)2.5μL,dNTP(10 mmol/L)1.5μL,Ex-Taq 1μL,模板DNA 2μL,加无菌水至50μL。
EX-TaqPCR扩增条件:94 ℃变性300 s;94℃、40 s,54 ℃、40 s,72 ℃、60 s,反应50个循环;72 ℃延伸10 min。

表1 样本信息及COI基因序列的GenBank登陆序列号
扩增产物纯化后,送至生工生物工程(上海)股份有限公司测序。
1.3 序列比对分析、系统发育树构建及样本种类确定
采用Gene-Explorer软件对43条基因序列进行修饰,利用BLAST检索其相似性,所得序列均是线粒体COI基因片段。使用DNA Barcoding鉴定方法,用CLUSTAL软件进行多序列比对[11-12],导出FASTA格式文件。采用MEGA10软件分析序列的保守位点(conserved sites,C)数、变异位点(variable sites,V)数、简约信息位点(parsimin-formative sites,Pi)数、种间遗传距离、种内遗传距离、碱基偏倚性等信息。采用邻接法,构建系统发育树(Neighbor-joining tree,NJ树)[13-14],并将测序基因片段在GenBank数据库内进行检索比对及鉴定种类。查阅文献,根据样本雄性外生殖器及外部特征,确定DNA条码鉴定结果的准确度。
2 结果与分析
2.1 COI基因序列特征分析
以43个样本的基因组DNA为模板,使用鳞翅目COI基因片段通用引物进行扩增,克隆得到了43条687 bp左右的基因片段,经过BLAST比对分析,序列均为线粒体COI基因序列。相关信息见表1。
根据雄性外生殖器及外部形态特征,43个夜蛾科昆虫样本的鉴定结果为10个种。其中,2个种类无中文名对应,且未查到这2个种的形态描述,未做形态鉴定。利用MEGA10软件进行碱基含量分析,使用CLUSTAL软件进行序列比对,43个样本COI基因序列中均未发生缺失和插入情况,保守位点数、变异位点数、简约信息位点数、单突变位点数分别为520个、163个、146个、16个。各碱基平均含量分别为:A(30.0%)、T(40.6%)、C(14.7%)、G(14.7%),其中G+C的含量(29.4%)明显低于A+T的含量(70.6%),这与昆虫线粒体碱基含量特征一致[15]。
由表2可知,10种夜蛾科昆虫COI基因的碱基颠换率(Transversional Pairs)和转换率(Transitionsal Pairs)分别为33%和19%,偏倚率R值(si/sv)为0.6。碱基的转换以TC(84.2%)为主,少数为AG(15.8%)间的转换;颠换多发生于TA碱基间(90.6%),CA碱基间颠换占总数的9.4%。鉴于物种间的亲缘关系越近碱基转换率越高,物种间亲缘关系越远则碱基替换率越高[8],故不同夜蛾科昆虫间亲缘关系可以通过COI基因片段多序列分析结果推断出。

表2 10种夜蛾科昆虫COI基因片段碱基替换
2.2 种内及种间遗传距离
基于p-distance模型,计算出43个样本种内及种间的遗传距离,并利用bootstrap(1 000次)进行检验[8]。由表3可知,种间遗传距离在0.069 8~0.116 3之间,平均值为0.092 9。其中,粉纹夜蛾和白条夜蛾的遗传距离是0.069 8,大耳纹夜蛾和中赫夜蛾的遗传距离是0.070 8;乏夜蛾和粉纹夜蛾之间遗传距离值最大(0.116 3)。43个样本种内遗传距离在0~0.001 5之间,平均值为0.000 5。种间遗传距离远大于种内遗传距离,且种内遗传距离最大值和种间遗传距离最小值并未重叠,符合DNA条形码检验的标准(种间差异是种内差异10倍)[16]。
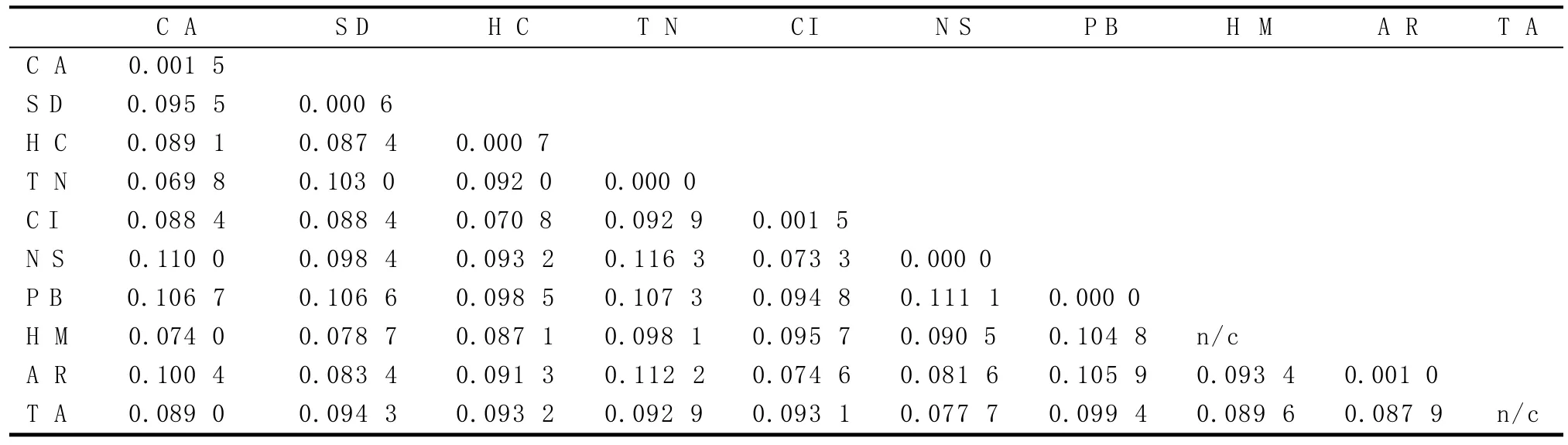
表3 夜蛾科10种不同害虫的种间和种内遗传距离
2.3 系统发育树构建分析
以43个样本的COI基因序列作为靶标,采用MEGA10软件进行数据分析,使用ClustalW功能进行序列比对,依据Kinura-2-Paramter 模型,采用邻接法(Neighbour-Joining)构建系统进化树,并进行重复1 000次的自举检验。由图1可知,不同种类样本形成了独立的进化分支。同时,聚类结果也反映了物种间的亲缘关系,如淡剑纹灰翅夜蛾、梨剑纹夜蛾是近缘种,这两个物种进化分支聚集在一起。因此,夜蛾科昆虫COI基因序列具有足够的遗传变异性和分化度,既可区分种间种类,同时种内相对保守。
3 结论与讨论
园林植物是城市园林绿化专类园和绿地的重要组成部分,虫害的发生会对园林植物的健康生长造成巨大威胁,所以虫害绿色防控是园林植保工作的重要环节,且只有在虫害发生后及时判别害虫的种类和发生规律,才能采取有效措施控制其为害;同时,对一个城市来说,一个区域的虫害发生情况,对该城市的整体绿化养管也具有一定的警示作用。因此,需要对害虫的消长动向进行预测预报,以便及时发出预警,从而更有效地对虫害进行绿色防控。但是,园林害虫种类繁多、形态各异,几乎所有的园林植物会受害,且在虫害大发生前较难被发现。而自动虫情测报灯的应用,可以准确地对害虫未来发生危害的时间和严重程度等情况进行预测预报,从而减少监测员的劳动强度。
自动虫情测报灯虽然能监测目标植物群落内活动昆虫种类的消长规律,且诱集到的昆虫种类多样,但诱集的鳞翅目昆虫成虫肢体和翅膀易破碎或粘成一团,较难准确区分和鉴定昆生种类[17],再加上植保技术人员和监测员在昆虫分类学专业性知识上相对欠缺,导致很难使用传统分类特征来区分昆生种类。同时,鳞翅目昆虫属于完全变态昆虫,其在不同发育阶段的形态特征差异较大,而鉴定往往要根据成虫特征来进行判断,但鳞翅目昆虫种类与数量庞大,且很多近似种成虫差异细微,这也给非专业鉴定人员带来了极大困难。因此,需在传统根据成虫进行鉴别分类的基础上,利用DNA条码技术对近似种、隐形种和新种进行鉴定,以缩短排查时间,提高工作效率,提升鉴定结果的精确性,这对于虫害综合治理意义重大[18]。
本研究结果表明,应用DNA条码技术辅助传统分类方法进行夜蛾科种类鉴定,具有较高的准确度;对43个样本基因序列进行比对分析,种间遗传距离远大于种内遗传距离,且种内遗传距离最大值和种间遗传距离最小值并未重叠,符合DNA条形码检验的标准;采用邻接法构建系统进化树的结果显示,不同种昆虫也会各自聚集到一个单系群。
DNA条码技术可辅助应用于自动虫情测报灯诱捕到的夜蛾科昆虫的分类鉴定与测报,其虽暂时不能取代传统分类工作,但将其作为辅助手段用于形态鉴定,可有效提高种类鉴定的效率和准确度。随着GenBANK等数据库的发展,DNA条码技术在夜蛾科昆虫种类鉴定工作中将得到更多的应用。
猜你喜欢
草业科学(2022年6期)2022-06-29
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
中学生理科应试(2021年10期)2021-12-07
云南农业(2021年11期)2021-11-12
今日农业(2021年9期)2021-07-28
今日农业(2020年17期)2020-12-15
今日农业(2020年13期)2020-12-15
今日农业(2020年16期)2020-12-14
科学与技术(2019年21期)2019-04-27
