
基于相对迁移率的节节麦醇溶蛋白组成分析
2020-08-17 06:33叶发慧察艳艳冯美玲刘瑞娟刘宝龙陈文杰张怀刚
西北农业学报 2020年6期
叶发慧 ,察艳艳,冯美玲,李 响,, ,刘瑞娟,,曹 东, ,张 波, ,刘宝龙,,陈文杰,,张怀刚
(1.中国科学院高原生物适应与进化重点实验室,中国科学院 西北高原生物研究所,中国科学院 种子创新研究院,西宁 810008;2. 中国科学院大学,北京 100049;3. 青海省作物分子育种重点实验室,西宁 810008;4.青海大学,西宁 810016;5.青海师范大学,西宁 810008)
普通小麦(TriticumaestivumL,AABBDD,2n=6x=42)是世界广布的重要粮食作物,为人类提供30%粮食[1],小麦面粉品质与人类的营养状况直接相关。小麦面粉品质由小麦胚乳中的贮藏蛋白决定。麦醇溶蛋白(Gliadin,Gli)是小麦储藏蛋白的主要成分之一,在胚乳蛋白中高达40%,可通过二硫键与麦谷蛋白结合成稳定的网状结构,从而赋予面筋一定的可塑性、粘性和延展性[2]。麦醇溶蛋白由小麦第1、第6部分同源群染色体短臂上的Gli-1和Gli-2位点编码[3]。Gli-1位点包括Gli-A1、Gli-B1和Gli-D1;Gli-2位点包括Gli-A2、Gli-B2和Gli-D2[4]。麦醇溶蛋白的位点都表现出广泛的等位基因变异,到目前为止,在普通小麦的Gli-1和Gli-2位点上已鉴定出多于130个等位变异[5]。根据醇溶蛋白在酸性(pH3.1)聚丙烯酰胺凝胶电泳(Acidic polyacrylamide gel electrophoresis,A-PAGE)中迁移率的不同,进一步将醇溶蛋白细分为α,β,γ和ω醇溶蛋白(迁移率大小为ω<γ<β<α)[6]。麦醇溶蛋白谱带组成几乎不受环境影响,完全由基因控制[7]。因此,它也可作为特殊的生化和分子标记,用于揭示不同材料间的遗传差异[8]。少数骨干亲本杂交等传统小麦育种手段,导致现代小麦品种的遗传背景日趋狭窄,严重限制了小麦面粉品质的进一步提高。从具有更广泛遗传变异的普通小麦野生近缘物种中,发掘具有优质基因的种质资源并加以利用,是普通小麦遗传改良的有效途径[9]。
节节麦(Aegilopstauschii,DD,2n=2x=14)是普通小麦D染色体组的二倍体供体物种[10],广泛分布于西起中东各国,东至中国的广阔地域,具有丰富的遗传多样性[11],拥有大量现代普通小麦品种所不具备的有益基因[12-14]。节节麦中醇溶蛋白位点的结构和作用机制与普通小麦相似[15-16],Gli-D1和Gli-D2位点具有多个等位基因的特征,这使得区分系、遗传、变种和亚种,研究种群结构和确定它们的系统发育关系成为可能[17]。节节麦D染色体组与普通小麦D染色体组完全同源,其优良基因可通过远缘杂交直接转移到普通小麦中, 培育新的普通小麦品种[18-20]。
本研究通过对212份节节麦材料的醇溶蛋白进行酸性聚丙烯酰胺凝胶电泳(A-PAGE)检测,分析其遗传多样性,旨在了解这些节节麦种质资源的醇溶蛋白多样性状况,为后续远缘杂交亲本的选择和进一步的小麦品种改良提供参考依据。
1 材料与方法
1.1 材 料
试验于2019年春季在中国科学院西北高原生物研究所青海省作物分子育种重点实验室 进行。
供试材料为笔者前期已收集的212份节节麦种质资源(表1、表2),其中15份来自中国,58份地理位置不详。
普通小麦‘中国春’作为对照材料,用以确定212份节节麦的醇溶蛋白组成类型。

表1 供试材料来源地Table 1 Origin of test materials
1.2 方 法
1.2.1 醇溶蛋白的提取 醇溶蛋白的提取参照傅宾孝等[21]的方法并稍做改动。(研磨籽粒的量由1 粒改为半粒,蛋白提取液由100 μL改为 50 μL)。
1.2.2 溶液配制 溶液的配制参照王祖华等[22]的方法。
1.2.3 电泳分离 用酸性聚丙烯酰胺凝胶电泳分离醇溶蛋白(A-PAGE)。将两块玻璃板清洗干净后用体积分数为70%的酒精再次清洗,涂上含量为2% 的剥离硅烷。将装好的玻璃板倾斜放置在泡沫板上,取5~10 mL凝胶液按体积比1∶1比例加入体积分数为1.2%的过氧化氢,迅速摇匀,沿长玻璃板内表面倒入底部,10 min后封底。玻璃板底部和泡沫板之间放 2 个10 μL规格的枪头,便于底部凝胶倒吸。取30 mL凝胶原液,加入体积比1∶1的体积分数为1%的过氧化氢,迅速摇匀,沿长玻璃板内表面倒入玻璃板夹缝中,直至溢出,迅速插入样品梳,静止3 h,慢慢垂直拔出样品梳。每个样品槽内加入样品8 μL,两端加入‘中国春’作对照。反向连接电路,设置额定电压350 V,电流30 mA,时间2 h。
1.2.4 染色 电泳结束后取下玻璃板,留下黏附凝胶的玻璃板,将凝胶和玻璃板一起放入考马斯亮蓝染色液中,染色1 h。将胶和玻璃板一同取出,清水冲去表面染料,无需脱色,连同玻璃板一起拍照保存。
1.2.5 命名 参照Autran等[23]的命名方法,以‘中国春’的电泳图谱为标准,‘中国春’的第1条谱带(迁移率最小的条带)作为标准带,其他谱带的迁移率与它的迁移率比值作为相对迁移率来确定各蛋白质条带的位置,‘中国春’第1条谱带的相对迁移率记为1。
1.3 数据处理
利用Gis软件获得迁移率;按Bushuk 等[24]的计算系统,以‘中国春’为对照,计算节节麦醇溶蛋白的相对迁移率。根据相对迁移率大小将电泳谱带分为α、β、γ 和 ω 4 个区。按照条带的有无将结果计为 1(有)和 0(无), 利用NTsys软件计算品种间相似系数(GS),用 GS值按不加权成对群算术平均法(U-PGMA)进行聚类分析。统计分析在 Microsoft Excel 2013中进行。
2 结果与分析
2.1 醇溶蛋白条带多态性分析
212份材料共分离出2 143 条谱带(部分材料电泳图见图1),单个材料变幅为7~16条,平均为10.11条,大部分有9~12条,占总条带的 73.11%(表2);18份材料(8.49%)分离出12条以上的谱带,39份材料(18.40%)分离出9条以下的谱带。来自巴基斯坦的388号分离出的谱带最多,为16条;其次是来自阿塞拜疆的346号材料,分离出15条谱带;19份材料(8.96%)均分离出7条谱带,主要来自阿塞拜疆和乌兹别克斯坦。来自中国的15份材料分离出13、12、11、10、9、8条谱带,分别占6.67%、20.0%、13.3%、 13.3%、20.0%和 26.67%。212份材料共分离出422种不同相对迁移率的谱带(表3),谱带相对迁移率的变幅为0.27~5.82;第一条谱带相对迁移率最小的是37号材料,这份材料两条谱带的相对迁移率为0.72和0.89,均小于1;最后一条谱带的相对迁移率为5.82,是44号材料和321号材料,其中44号材料分离出的最小谱带的相对迁移率为0.99。谱带频率变幅为0.24%~ 7.35%,44号材料相对迁移率出现次数最多,为 7.35%,其次是58号;有94种相对迁移率只出现1 次。对照‘中国春’共分离出17条谱带,迁移率为1~5.15,在所有材料中,有28份材料谱带的相对迁移率小于1,其中6份材料的2条带均小于1;谱带相对迁移率大于 5.15的材料共有60份,18号、19号、24号、34号、41号、43号和44号7份材料均分离出12~13条谱带,第一条谱带的相对迁移率小于1,最后一条谱带的相对迁移率大于5.15。
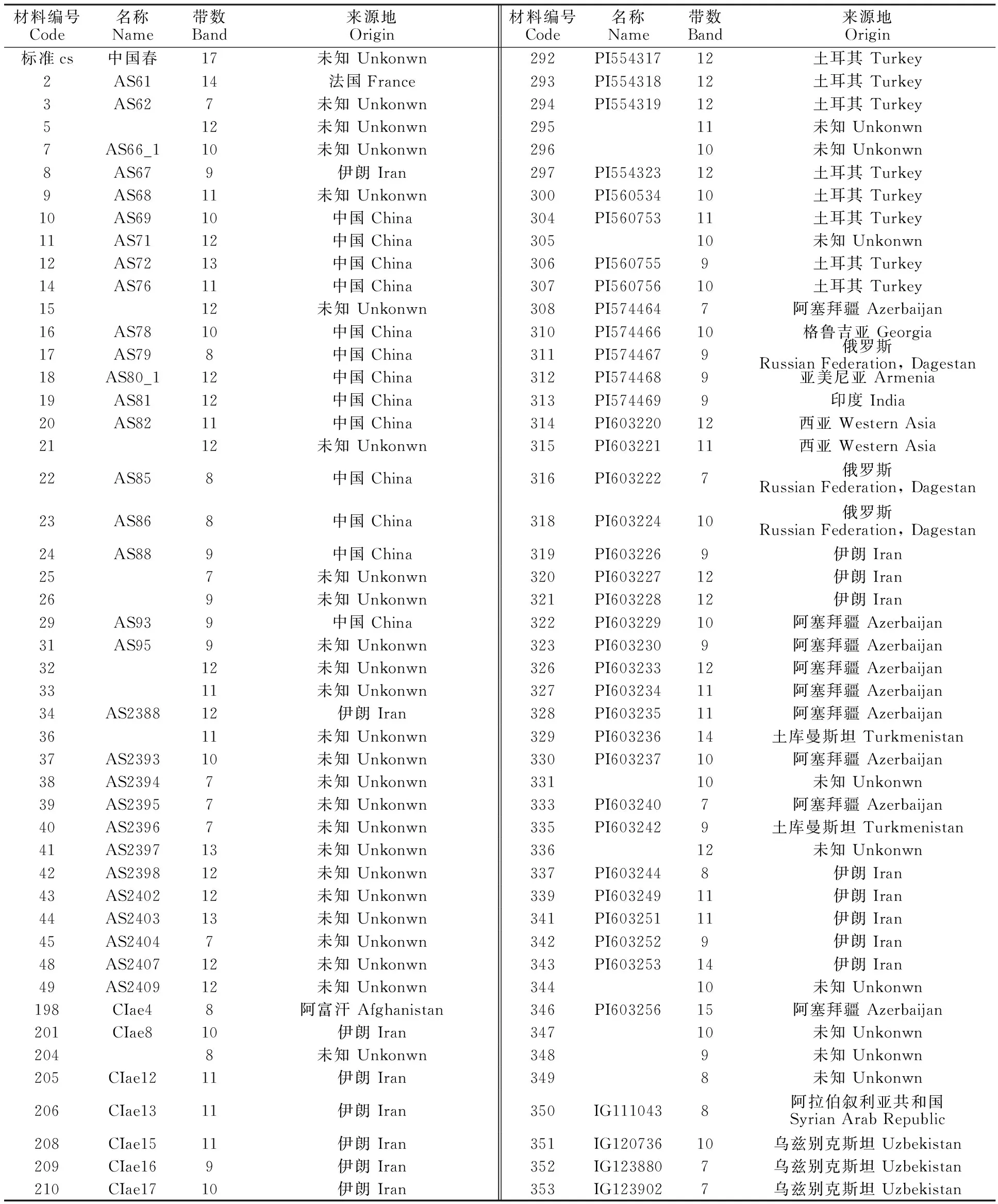
表2 供试材料编号及其醇溶蛋白带数Table 2 Number of test materials and the number of gliadin bands
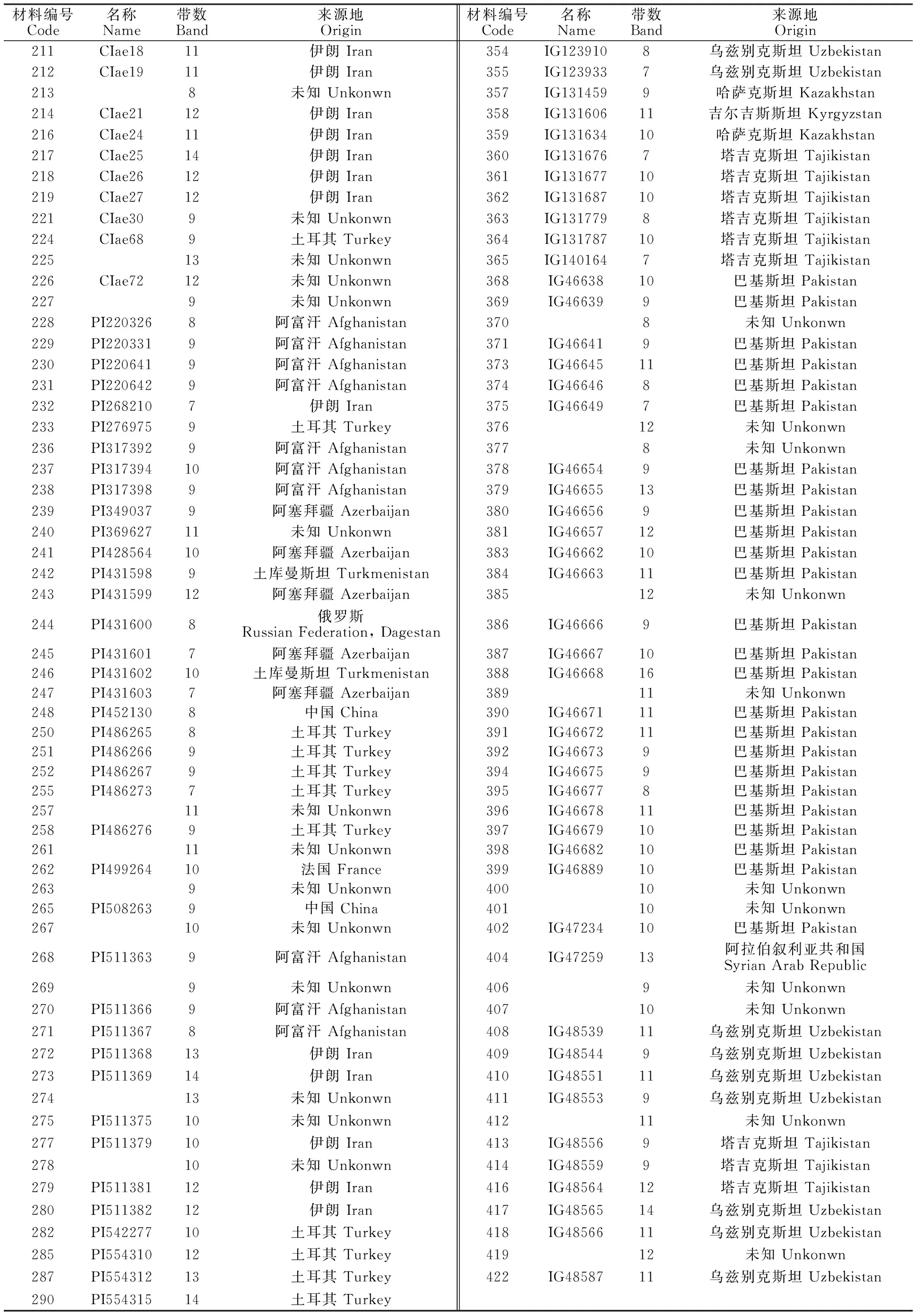
(续表2 Continued table 2)

箭头代表相对迁移率为1的主带
参照Bushuk 等[24]的方法,根据相对迁移率大小将422种谱带分为 α、β、γ 和 ω 4 个区(表4)。α区有362条谱带,占总数的16.89%,127种变异类型,占总数的30.09%;β区有470条谱带,占总数的21.93%,有60种变异类型,占总数的14.22%;γ区有483条谱带,占总数的 22.54%,有105种变异类型,占总数的24.88%;ω区有828条谱带,占总数的30.81%,有130种变异类型,占总数的30.81%。ω区谱带数最多,占总谱带数的38.64%,α区多态性条带最多,高达34.70%,谱带组合类型最多。
2.2 遗传相似系数分析
利用NTsys软件对212份材料进行遗传相似性分析,发现这些材料的遗传相似系数(GS)变幅为0~0.58,平均值为0.037。不存在GS值为1的现象,而有68.54%的GS值为0。来自中国的18号和19号材料的GS值最高(0.58),表明二者亲缘关系最近。
2.3 U-PGMA聚类分析
对212份材料进行基于UPGMA法的聚类分析(图2),212份材料在GS=0.032水平上可分为5个大类。第Ⅰ类包含8份材料,部分材料来自伊朗(3份)和乌兹别克斯坦(1份),可分为2个亚类,各包含4份材料;这一类群的遗传相似系数最小,分布面积较局限,并没有大面积出现。第Ⅱ类包含35份材料;部分材料来自伊朗(7份)、中国(7份)、塔吉克斯坦(4份)、土耳其(2份)、巴基斯坦(1份)、格鲁尼亚(1份)、阿塞拜疆(2份)和法国(2份),可分为2个亚类,分别包含6份和29份材料。这一类群中包含7份伊朗材料和7份中国材料,两者遗传相似系数高;中国的材料1份来自新疆(11号),3份来自河南(18号、19号和20号),3份来源地不详(10号、22号和23号)。第Ⅲ类包含23份材料,部分来自伊朗(6份)、中国(1份)、土库曼尼斯坦(1份)、乌兹别克斯坦(3份)、阿富汗(1份)、土耳其(1份)、巴基斯坦(3份)、塔吉克斯坦(1份)和阿塞拜疆(1份),可分为2个亚类,分别包含9份和14份材料。第Ⅳ类包含8份材料,部分来自伊朗(1份)和巴基斯坦(2份),可分为2个亚类,分别包含3份和5份材料。第Ⅴ类包含139份材料,部分来自伊朗(11份)、巴基斯坦(18份)、阿富汗(9份)、土耳其(15份)、乌兹别克斯坦(8份)、阿塞拜疆(8份)、中国(7份)、阿拉伯(2份)、哈萨克斯坦(2份)、吉尔吉斯斯坦(1份)、俄罗斯(4份)、塔吉克斯坦(4份)、土库曼尼斯坦(1份)、亚美尼亚(1份)和印度(1份),可分为3个亚类,分别包含43、58和37份材料;7份来自中国,1份新疆(12号),2份陕西(14号和248号),2份河南(16号和265号),2份产地不详(24号和29号);中国的材料与巴基斯坦、阿富汗的材料亲缘关系较近。笔者据此推测,伊朗地区至少存在5类节节麦居群,并具有5种可能的对外传播扩散路径(图3)。

表3 不同迁移率在212份材料中出现的频率Table 3 Frequency of different mobilities in 212 materials

(续表3 Continued table 3)

表4 4个区醇溶蛋白带型统计结果及多样性Table 4 Statistical results of gliadin in four zones and diversity of gliadin
3 讨 论
1959年Jones第一次将醇溶蛋白划分为α、β、γ 和 ω 4个区[7],1975年Gubareva利用加拿大栽培小麦的电泳图对其进行命名,称为“数字字母命名”系统[24]。其公式表示方法为:按迁移率的大小,将α、β、γ和ω 4 个区域分别分成 1~7、1~5、1~5 和 1~12 条谱带;染色强烈的谱带的数字添加下划线表示,染色弱的数字添加圆括号表示,而双重线则用数字上方的两个点表示。下标1表示2区移动快的临近波,下标2表示移动慢的临近波。同年,Autran等[25]利用生长在法国的春小麦和冬小麦的电泳图再次对4区进行规范,根据迁移率为65的γ-醇溶蛋白计算其余谱带的相对迁移率,并将相对迁移率为0.65的谱带定位主带。在后面研究中发现,法国命名系统并不能应用到加拿大栽培小麦上,1977年Bushuk等[24]发现这一问题,并将法国材料的主带定为相对迁移率为0.5的谱带。因为试验材料的地域等差异,本研究中的材料未找到其迁移率为0.5的主带,由此可见,麦醇溶蛋白现有的命名方法并不具备广泛的普适性。笔者经过多次重复试验证明,在保证时间、电压、电流等试验参数一致的前提下,‘中国春’的第1条谱带清晰易辨识,相对位置也比较稳定,可作为对照标准带。将其他谱带的迁移率和‘中国春’第1条谱带迁移率的比值作为相对迁移率来确定各醇溶蛋白条带的位置,可准确鉴定每一条醇溶蛋白谱带,且无需除‘中国春’以外的其他材料作为对照。


图2 212份种质材料醇溶蛋白聚类图Fig.2 Clusters of 212 accessions based on gliadin band patterns

图3 节节麦可能的传播路径Fig.3 Material propagation path
本研究从212份节节麦材料中共发现422种不同迁移率的谱带,谱带出现频率变幅为0.24%~ 7.35%。谱带相对迁移率变幅为0.27~5.82。根据相对迁移率的大小,可将谱带分为 α、β、γ 和 ω 4 个区,其中处于α区谱带的相对分子质量最小,相对迁移率最大,可能由于扩散效应等原因,在A-PAGE胶图上最不清晰。为提高该区谱带的辨识度,建议使用双向电泳等其他方法验证α区谱带组成。本研究中,谱带迁移范围最大的是44号材料,范围最小的是阿富汗的270号材料。小于对照‘中国春’第1条主带和大于对照‘中国春’最后一条谱带的材料分别有28份和60份。有7份材料谱带的最小相对迁移率小于1,最大相对迁移率大于5.15(谱带迁移范围大于对照‘中国春’)。这7份材料在聚类图(图2)中分布在Ⅱ类(4份)、Ⅲ类(1份)和Ⅴ类(2份),在第Ⅱ类的4份中,18号材料和19号材料来自中国,34号材料地理位置不详,但它和来自中国的材料亲缘关系最近;同样,位于第Ⅲ类群的44号材料也是材料地理位置不详,但它和来自中国的材料近缘关系最近。位于第Ⅴ类群中的24号材料也来自中国。由此可见,中国的节节麦居群中存在着具有较广迁移率范围醇溶蛋白谱带的材料。醇溶蛋白谱带迁移范围异常的材料是否会对小麦品质改良有积极作用呢?值得进一步研究。
结合参试材料的聚类结果(图2)和地理分布(图3),发现来自伊朗的材料遗传多样性最丰富,此结果支持节节麦的起源地是伊朗地区的观点[26]。中国境内的节节麦至少存在 3 种类型。一类(类型Ⅴ)遍布节节麦的自然分布区,而另两类(类型Ⅱ和Ⅲ)则是在西北南三个方向上仅存在于伊朗及其毗邻地区,唯独向东一路分布到中国。这两类材料是以伊朗→土库曼尼斯坦→塔吉克斯坦→中国的单线路径传播,并没有向四周扩散,而这一路径正好与古丝绸之路陆路中的一条不谋而合,由此可见,至少不能排除这类材料是经古丝绸之路等人为途径传播至中国的可能。从聚类结果来看,类型Ⅱ中,和伊朗原始材料亲缘关系最近的11号材料来自中国新疆,18、19和20号材料来自中国河南。来自新疆的12号材料、陕西的14号、248号材料和河南的265号材料同处于第Ⅴ类群中,亲缘关系较近,根据这些结果可以推测,中东地区节节麦从阿富汗和巴基斯坦传入中国新疆地区后(主要是巴基斯坦),很可能继续向东传播到黄河中下游地区。这与笔者之前新疆及黄河流域节节麦的高分子谷蛋白研究结果一致(未发表资料)。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
昆明医科大学学报(2021年1期)2021-02-07
湖北农业科学(2019年11期)2019-07-22
商情(2019年26期)2019-06-25
发明与创新·小学生(2019年5期)2019-06-14
小学生作文(中高年级适用)(2018年10期)2018-10-26
电子技术与软件工程(2018年20期)2018-02-28
中学物理·高中(2016年3期)2016-04-07
分析化学(2015年7期)2015-07-30
医学研究杂志(2015年7期)2015-06-22
