
基于配合力和遗传距离的甜高粱杂种优势预测
2020-08-15 02:41王黎明严洪冬焦少杰姜艳喜苏德峰孙广全
中国农业科学 2020年14期
王黎明,严洪冬,焦少杰,姜艳喜,苏德峰,孙广全
基于配合力和遗传距离的甜高粱杂种优势预测
王黎明,严洪冬,焦少杰,姜艳喜,苏德峰,孙广全
(黑龙江省农业科学院作物资源研究所,哈尔滨 150086)
【】对甜高粱主要农艺性状进行杂种优势、一般配合力及特殊配合力分析,同时,分析配合力、表型遗传距离以及分子遗传距离用于杂种优势预测的可行性,为甜高粱的种质创新和杂交种选育提供理论参考。采用不完全双列杂交设计,以8个甜高粱不育系为母本及8个甜高粱恢复系为父本配制64个杂交组合。对亲本及杂交后代进行2年的性状调查,包括:出苗至开花日数、生育期、株高、穗长、茎粗、分蘖、单穗粒重、千粒重、籽粒产量、单株重、生物产量和含糖量。分析不同性状的杂种优势、一般配合力、特殊配合力、表型遗传距离、分子遗传距离以及配合力、遗传距离与杂种优势的相关性。各性状的中亲优势由强到弱分别为单株重、籽粒产量、单穗粒重、生物产量、株高、穗长、千粒重、茎粗、生育期、至开花日数、分蘖和含糖量,其中,生育期、至开花日数、分蘖和含糖量等性状为负优势。不同性状的中亲优势和超亲优势由强到弱的顺序基本相同。配合力分析表明,每个性状中,不同亲本的一般配合力相差较大,且不同组合的特殊配合力也有很大差异。大多数特殊配合力高的组合,其亲本的一般配合力也较高。杂种优势与配合力和遗传距离的相关性为单株重、籽粒产量、单穗粒重、生物产量、穗长、千粒重、分蘖以及含糖量等性状的杂种优势与其亲本的一般配合力和特殊配合力均为极显著正相关。生育期的杂种优势与特殊配合力为极显著正相关,至开花日数与特殊配合力为显著正相关。亲本间的表型遗传距离为2.86—6.82,分子遗传距离为0.50—0.96。单株重、籽粒产量、单穗粒重、生物产量、株高、穗长、茎粗及含糖量等性状的杂种优势与分子遗传距离的相关性大于表型遗传距离,其中,生物产量、单株重、穗长和茎粗的杂种优势与分子遗传距离为极显著正相关。所有性状中,与产量相关性状的杂种优势较高,而含糖量和分蘖的杂种优势较低。在杂种优势预测上,利用亲本的配合力可有效预测杂种优势,预测效果优于遗传距离。与表型遗传距离相比,分子遗传距离对杂种优势的预测更有效。
甜高粱;杂种优势;配合力;遗传距离;预测
0 引言
1 材料与方法
1.1 试验材料
分别利用在农艺性状上差异较大的8个甜高粱不育系为母本及8个甜高粱恢复系为父本配制杂交组合,其中,307A和8431为对照,454A、4190A和314A为生产上常用的亲本材料(电子附表1)。
1.2 性状测定方法
采用不完全双列杂交设计(NCⅡ设计),以8个不育系和8个恢复系为亲本共配制64个杂交组合。于2012年和2013年种植在黑龙江省农业科学院(哈尔滨)试验基地,随机区组排列,行长5 m,5行区,垄距65 cm,3次重复。对亲本及杂交组合进行2年的主要农艺性状调查,调查项目包括出苗至开花日数、生育期、株高、穗长、茎粗、分蘖、单穗粒重、千粒重、籽粒产量、单株重、生物产量和含糖量等性状。所有性状的调查标准参照陆平[25]的方法。其中,含糖量的测定为田间采样后,整株榨汁,用测糖仪进行测定,以锤度计算。选择中间3行测定籽粒产量和生物产量。
1.3 分子遗传距离测定
利用筛选出的41对多态性高、扩增稳定且分布于所有连锁群上的SSR引物用于PCR扩增(电子附表2)。DNA提取及PCR扩增的具体方法见文献[26]。
1.4 数据分析
运用SAS软件对杂种优势、一般配合力、特殊配合力、表型遗传距离和分子遗传距离进行分析。其中,杂种优势分析包括中亲优势和超亲优势,计算公式为:
中亲优势(%)=[1-(1+2)/2]/(1+2)×100;
超亲优势(%)=(1-)/×100;
式中,1为杂交后代值,1为母本值,2为父本值,为高亲值。
参照孔繁玲等[27]方法计算一般配合力、特殊配合力和表型遗传距离。其中,表型遗传距离采用欧氏距离。
大部分焊接工艺都要求直接或间接地对纳米线结进行加热熔融焊接,但是在一些特定的环境中难以获得纳米尺寸的热源,很少有方法可以连接两个目标纳米线,并保持周围纳米物体的完整性,而冷焊接则是一种不需要任何热源的焊接方法[33],从而使其在纳米尺度上成为有吸引力的解决方案.
按照Nei等[28]公式计算亲本间的遗传相似系数GS=2Nxy/(Nx+Ny),其中,GS为亲本间的相似系数,Nxy代表2个亲本共有的等位基因,Nx和Ny分别代表亲本x和y的等位基因数,遗传距离GD=1-GS。
2 结果
2.1 主要农艺性状的杂种优势分析
2.1.1 中亲优势分析 各性状的中亲优势分析结果(表1)表明,不同性状的中亲优势差别很大,由强到弱分别为单株重、籽粒产量、单穗粒重、生物产量、株高、穗长、千粒重、茎粗、生育期、至开花日数、分蘖和含糖量。产量性状的中亲优势强于其他性状,其中,单株重的中亲优势最高,平均为89.17,且所有组合均为正值。与产量性状相比,株高的中亲优势略低,均为正优势,且最高值较高,为633.33。另外,生育期、至开花日数、分蘖和含糖量的平均中亲优势均为负值,其中,含糖量的中亲优势最低。至开花日数和生育期的中亲优势组合间最高值和最低值相差较小。分蘖的中亲优势平均值虽然为负值,但后代的杂种优势差异很大,最高值达766.67,最低值为-100.00,可利用组合间的差异性选择目标性状。各性状中,至开花日数、生育期、茎粗、分蘖、单穗粒重和含糖量等性状中,超过半数杂交组合的中亲优势高于对照组合,而株高、穗长、千粒重、籽粒产量、生物产量和单株重等性状中,只有少数组合的中亲优势高于对照组合。
2.1.2 超亲优势分析 从各性状的超亲优势结果(表2)可以看出,杂交后代在各性状中的超亲优势差别很大。超亲优势由强到弱分别为单株重、籽粒产量、单穗粒重、生物产量、株高、穗长、千粒重、茎粗、生育期、至开花日数、含糖量和分蘖。各性状中,产量性状的超亲优势强于其他性状,其中单株重的超亲优势最高,平均为61.70。超亲优势略低的性状为株高、穗长和千粒重。而茎粗、生育期、至开花日数、含糖量和分蘖的超亲优势均为负值。其中,分蘖的超亲优势最低,但后代中的超亲优势差异很大。各性状中,至开花日数、生育期、茎粗、分蘖、单穗粒重和含糖量等性状中,超过半数杂交组合的超亲优势高于对照组合,而株高、穗长、千粒重、籽粒产量、生物产量和单株重等性状中,只有少数组合的超亲优势高于对照组合。
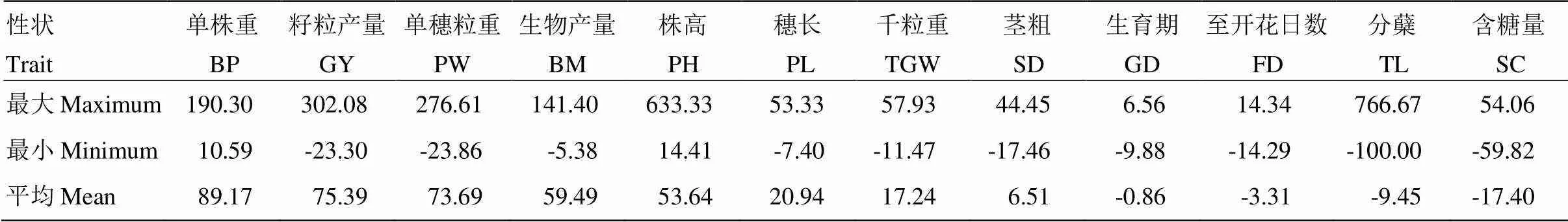
表1 主要农艺性状的中亲优势分析
FD:至开花日数;GD:生育期;SD:茎粗;TL:分蘖;PH:株高;PL:穗长;BM:生物产量;BP:单株重;GY:籽粒产量;PW:单穗粒重;TGW:千粒重;SC:含糖量。下同
FD: Days to flowering; GD: Growth duration; SD: Stem diameter; TL: Tillers; PH: Plant height; PL: Panicle length; BM: Biomass; BP: Biomass/plant; GY: Grain yield; PW: Panicle weight; TGW: 1000-grain weight; SC: Sugar content. The same as below

表2 主要农艺性状的超亲优势分析
通过对比各性状的中亲优势和超亲优势可以看出,各性状的中亲优势和超亲优势由强到弱的顺序基本相同,只有含糖量和分蘖略有不同。
2.2 杂种优势与配合力的相关性分析
2.2.1 一般配合力分析 通过对不育系和恢复系在所有性状中的一般配合力效应进行分析(表3),结果显示,每个性状中,不同亲本的一般配合力效应相差较大。同时,同一亲本在不同性状的配合力表现上也有很大差异。各性状中,一般配合力最高的亲本分别为至开花日数、籽粒产量、生物产量以及单株重为A3;生育期为R6;株高为R2;穗长以及单穗粒重为R4;茎粗为R5;分蘖为A5;千粒重为A8以及含糖量为R1。以上亲本在多个性状中的一般配合力高于对照。
2.2.2 特殊配合力分析 在所测定的杂交组合中,不同亲本间各性状的特殊配合力效应差异较大(表4)。不同组合在同一性状间以及同一组合在不同性状上的特殊配合力表现均有较大差异。其中,大多数性状中特殊配合力最高的组合中至少有1个亲本的一般配合力最高或较高,如株高A3/R2、穗长A3/R4、分蘖A5/R8、籽粒产量A3/R1、单穗粒重A4/R4、千粒重A8/R5、单株重A2/R4、含糖量A1/R4以及至开花日数A5/R6。而生育期、茎粗和生物产量中特殊配合力最高的组合,其亲本的一般配合力均不高,但这几个性状中特殊配合力较高的组合中,其亲本的一般配合力也较高。与对照相比,至开花日数、生育期、茎粗和分蘖等性状中一半以上组合的特殊配合力高于对照,但其他性状中高于对照的杂交组合较少。
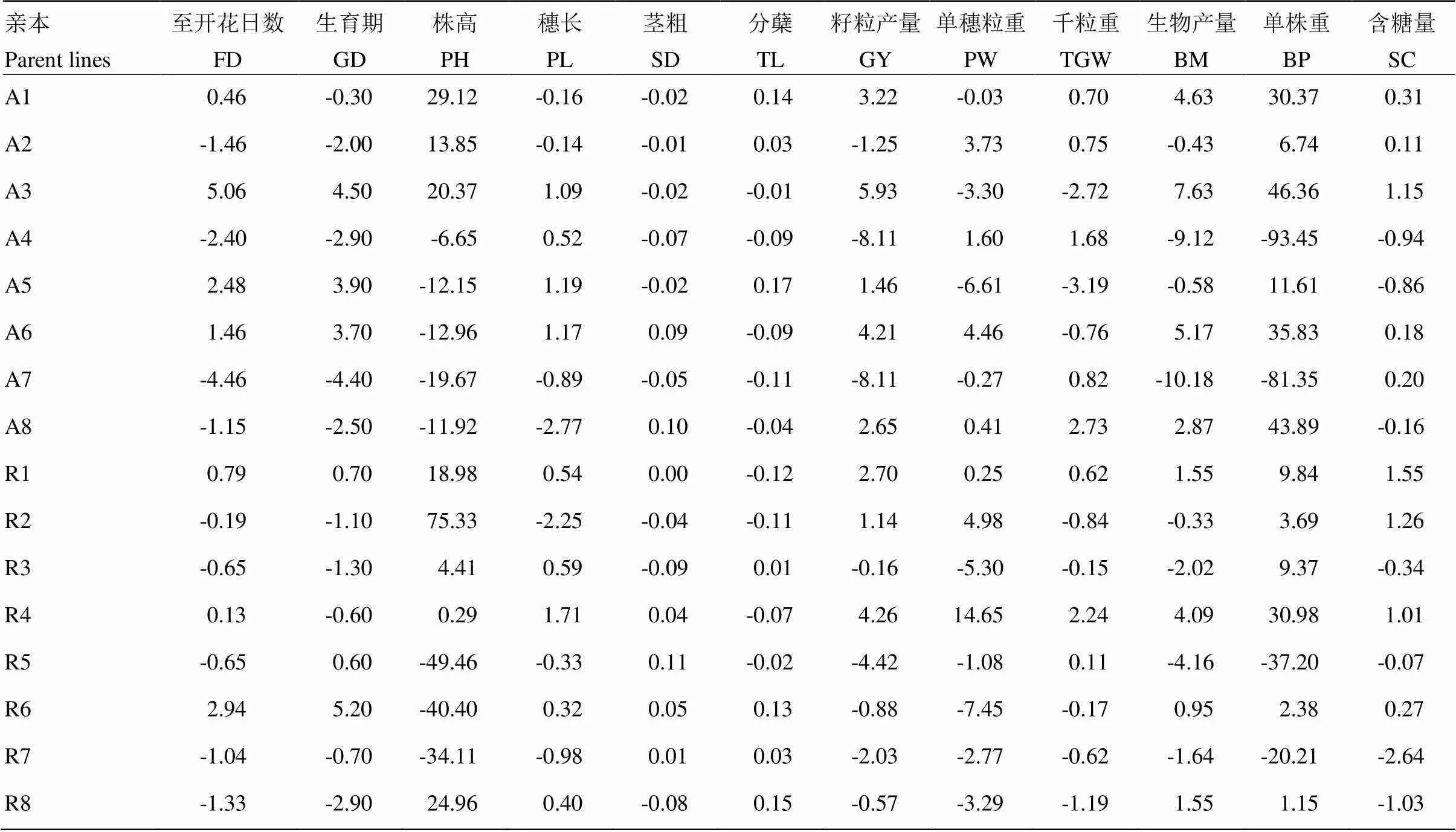
表3 亲本主要农艺性状的一般配合力效应

表4 亲本主要农艺性状的特殊配合力效应
2.2.3 杂种优势与配合力的相关性 通过对配合力和中亲优势的相关性进行研究(表5),大多数性状的杂种优势与其亲本的一般配合力和特殊配合力相关显著或极显著。其中,单株重、籽粒产量、单穗粒重、生物产量、穗长、千粒重、分蘖以及含糖量的杂种优势与其亲本的一般配合力和特殊配合力均为极显著正相关。并且,含糖量的杂种优势与一般配合力和特殊配合力的相关程度较高,相关系数均在70以上,同时,籽粒产量与一般配合力的相关系数也达到70以上。在所有相关系数达极显著的性状中,除籽粒产量的杂种优势与一般配合力的相关性显著高于特殊配合力外,其余性状的一般配合力和特殊配合力之间的相关系数差异较小。生育期的杂种优势与特殊配合力为极显著正相关,至开花日数与特殊配合力为显著正相关。而株高和茎粗的杂种优势与各配合力相关均不显著。
2.3 杂种优势与遗传距离的相关性分析
2.3.1 表型遗传距离分析 亲本间的表型遗传距离平均为4.73(表6),其中,最高值为6.82(A7和R6之间),最低值为2.86(A3的R2之间),不同亲本的表型性状差异较大。
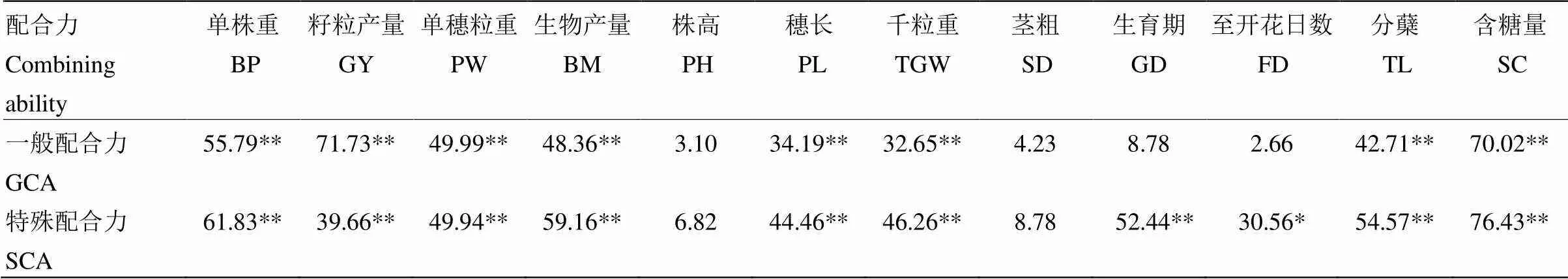
表5 杂种优势与配合力的相关性
GCA:一般配合力;SCA:特殊配合力;*:<0.05水平相关性差异显著;**:<0.01水平相关性差异显著。下同
GCA: General combining ability; SCA: Special combining ability; *: Significance at<0.05; **: Significance at<0.01. The same as below

表6 亲本间的表型遗传距离
2.3.2 分子遗传距离分析 利用SSR引物扩增后,每个引物检测到的等位基因为2—11个,平均4.6个。引物的多样性指数介于0.094—0.870,平均为0.586。
分子遗传距离结果(表7)表明,亲本间的分子遗传距离平均为0.74,其中,最高值为0.96(A3和R6之间),最低值为0.50(A5和R5之间),说明不同亲本在遗传水平上差异较大。
2.3.3 杂种优势与遗传距离的相关性 分别对表型遗传距离和分子遗传距离与中亲优势的相关性进行分析(表8)。结果表明,单株重、籽粒产量、单穗粒重、生物产量、株高、穗长、茎粗及含糖量等性状的杂种优势与分子遗传距离的相关性大于表型遗传距离。所有性状的杂种优势与表型遗传距离相关性均不显著,但单株重、生物产量、穗长和茎粗的杂种优势与分子遗传距离为极显著正相关。而籽粒产量、单穗粒重、株高和含糖量等性状的杂种优势与其分子遗传距离虽然相关系数较高,但没达到显著水平。另外,籽粒产量、单穗粒重、株高、千粒重、茎粗、生育期、至开花日数和分蘖的杂种优势与表型遗传距离的相关性较高,也没有达到显著水平。以上结果表明,可利用分子遗传距离对甜高粱部分性状的杂种优势进行预测。
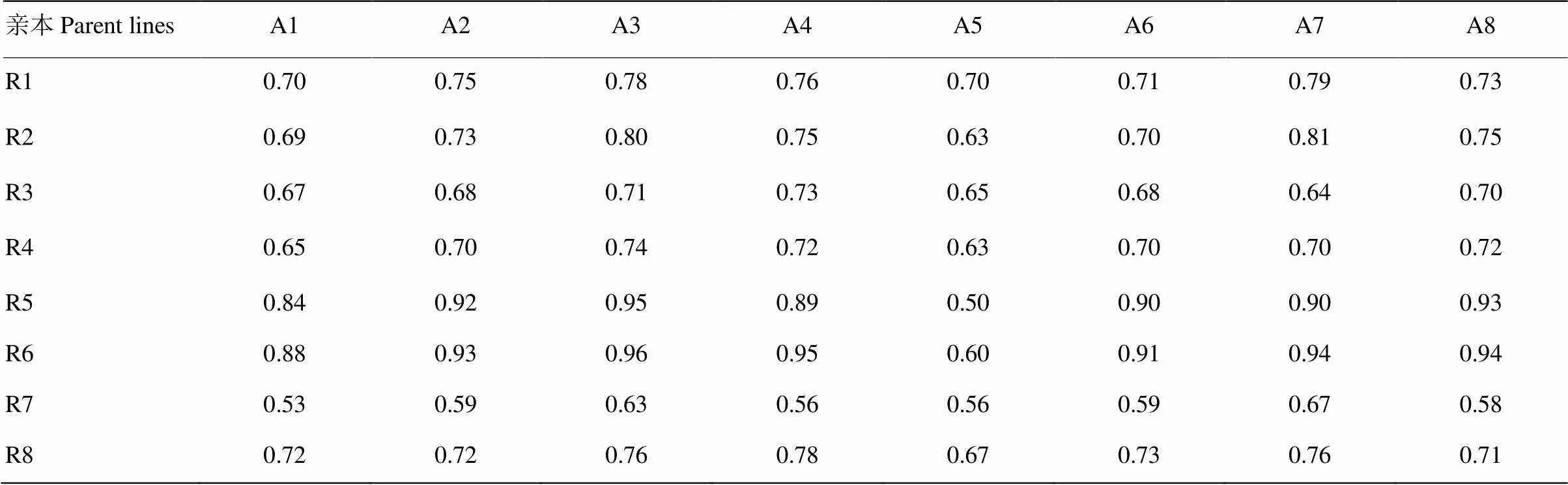
表7 亲本间的分子遗传距离
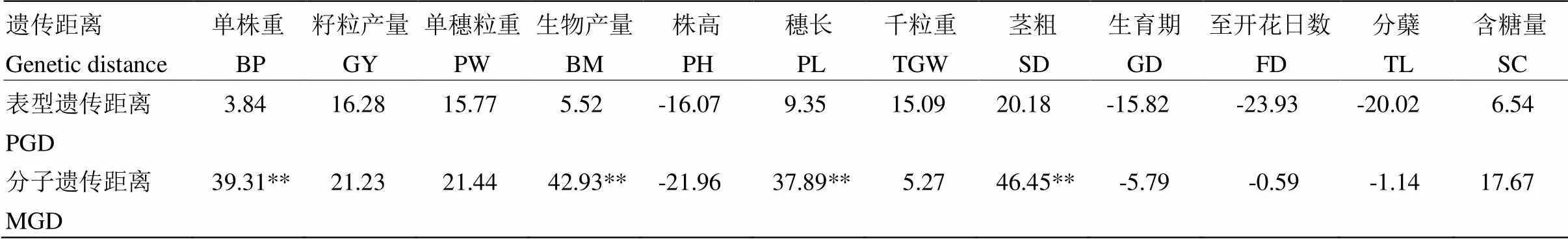
表8 杂种优势和遗传距离的相关性
PGD:表型遗传距离;MGD:分子遗传距离PGD: Phenotypic genetic distance; MGD: Molecular genetic distance
3 讨论
3.1 甜高粱杂种优势的利用
杂交后代各性状的中亲优势和超亲优势由强到弱的顺序基本相同。在所有性状中,单株重、籽粒产量、单穗粒重及生物产量等产量性状的杂种优势高,其次为株高,说明利用杂种优势易获得植株高大且籽粒产量和生物产量均高的杂交后代。穗长、千粒重和茎粗的杂种优势为中等,杂交后代的优势一般。至开花日数、生育期、分蘖和含糖量的杂种优势均为负优势,说明杂交后代的生育期容易变早,且不易获得分蘖多以及含糖量高的杂交后代。BUNPHAN等[18]研究也发现甜高粱的生物产量和籽粒产量的杂种优势较高,但含糖量的杂种优势低,研究结果与本研究相近。
从配合力分析结果来看,亲本在不同性状中的一般配合力和特殊配合力效应相差较大。因此,根据不同育种目标,对亲本的一般配合力和特殊配合力可有针对地利用。同时,特殊配合力较高的组合,其亲本的一般配合力往往也较高。以往研究也表明,虽然在一些性状上特殊配合力比一般配合力重要[12,18-19],但在亲本选择时不能只依靠特殊配合力效应,应将特殊配合力和一般配合力相结合,在一般配合力高的基础上,再选择特殊配合力高的亲本[20-21],以增加创造优异组合的机率。
此外,在选育甜高粱杂交种时,应根据杂交种的具体用途对不同性状重点选育。当用于提取乙醇或制糖时,要重点对生物产量和含糖量进行选育,而籽粒产量及其他性状则不是选育的重点[19,29]。当用于青贮饲料时,则要求杂交种的生物产量和籽粒产量都高,但含糖量可以不必太高[30]。此外,研究表明,黑龙江、吉林、内蒙古以及辽宁等省/自治区是中国利用甜高粱提取乙醇最具生产潜力的省份[31],为适应这些地区机械化水平相对较高的特点,可通过降低株高、增加种植密度来增加群体生物产量和糖产量的方式,选育适宜机械化种植的甜高粱品种[6,32],以有效降低生产成本,大力发展甜高粱种植。
3.2 甜高粱的杂种优势预测
在甜高粱的杂交育种中,含糖量及产量性状很难根据亲本的表型性状来预测杂交种的性状表现[33]。为此,育种者也在探讨更有效的预测方法。随着分子标记的不断发展,利用分子标记进行杂种优势的预测已在多种作物上进行了研究[34-36]。
本研究中,通过对比甜高粱亲本的表型遗传距离和分子遗传距离与杂种优势的相关性发现,大多数性状的杂种优势与分子遗传距离的相关性大于表型遗传距离,其中,单株重、生物产量、穗长和茎粗的杂种优势与亲本的分子遗传距离为极显著正相关。由此可见,与表型遗传距离相比,分子遗传距离对杂种优势的预测更有效。王瑞等[24]分析了粒用高粱的杂种优势与亲本遗传距离的相关性,也认为高粱亲本的选配应充分考虑遗传距离,为获得杂种优势强的后代,应选择遗传距离大的亲本杂交。同时,WEGARY等[15]采用欧氏距离的表型遗传距离和SSR分子遗传距离分析与玉米杂种优势的相关性,结果表明,分子遗传距离和籽粒产量为极显著正相关,和至开花期和抽丝期为极显著负相关,认为分子遗传距离对杂交种产量及一些农艺性状的预测比表型遗传距离更有效,研究结果与本研究相近。在水稻[37]、棉花[16]等作物中也有相似的研究结果。另外,本研究对甜高粱配合力与杂种优势的相关性研究显示,包括单株重、籽粒产量、单穗粒重、生物产量、穗长、千粒重、分蘖以及含糖量等大多数性状的杂种优势与其亲本的一般配合力和特殊配合力均为极显著正相关,亲本的配合力在甜高粱杂种优势的预测上与遗传距离相比,可对更多性状进行有效预测。AMELEWORK等[23]和JORDAN等[38]对粒用高粱的研究表明,表型遗传距离和分子遗传距离与籽粒产量杂种优势的相关性虽然不显著,但和一般配合力效应有关,认为在杂种优势的预测上,基于一般配合力的预测方法比基于分子标记的预测方法更有效。
在不同种质和不同物种上遗传距离用于杂种优势的预测效果不同,可能有以下几种原因。首先,研究结果因所选择材料群体的数量、亲本的多样性以及所研究的性状而不同,且不同类型的农艺性状受环境影响程度也不同。另外,缺乏和测定性状连锁的控制基因,所选择的分子标记和数量具有随机性,使标记在染色体上的分布不均衡以及显性基因的多样性效应等[39-40]。利用分子多样性来预测杂交种表现的一个基础假设是由于在性状QTL和标记等位基因间存在高水平的配子阶段连锁不平衡[34,38],因此,当用于计算遗传距离的所选标记比例超过50%以上与目标性状和杂种优势的QTL相关时,则利用分子标记预测杂种优势是可行的[39-40]。
4 结论
在甜高粱杂种优势预测上,利用亲本的一般配合力和特殊配合力可对单株重、籽粒产量、单穗粒重、生物产量、穗长、千粒重、分蘖、含糖量、至开花日数及生育期等性状的杂种优势进行预测,预测效果优于表型遗传距离和分子遗传距离。与表型遗传距离相比,分子遗传距离对杂种优势的预测更有效,尤其在单株重、生物产量、穗长和茎粗等性状的杂种优势预测上。
[1] BERENJI J, DAHLBERG J. Perspectives of sorghum in Europe., 2004, 190: 332-338.
[2] APPIAH-NKANSAH N B, LI J, ROONEY W, WANG D. A review of sweet sorghum as a viable renewable bioenergy crop and its techno-economic analysis., 2019, 143: 1121-1132.
[3] 邹剑秋, 王艳秋. 我国甜高粱育种方向及高效育种技术. 杂粮作物, 2007, 27(6): 403-404.
ZOU J Q, WANG Y Q. Sweet sorghum breeding objective and efficient breeding technology in China., 2007, 27(6): 403-404. (in Chinese)
[4] MISHRA J S, KUMAR R, RAO S S. Performance of sweet sorghum () cultivars as a source of green fodder under varying levels of nitrogen in semi-arid tropical India., 2017, 19(5): 532-538.
[5] TAKAKI M, TAN L, MURAKAMI T, TANG Y Q, SUN Z Y, MORIMURA S, KIDA K. Production of biofuels from sweet sorghum juice via ethanol-methane two-stage fermentation., 2015, 63: 329-336.
[6] LIU H H, REN L T, SPIERTZ H, ZHU Y B, XIE G H. An economic analysis of sweet sorghum cultivation for ethanol production in North China., 2015, 7: 1176-1184.
[7] TAZOE Y, SAZUKA T, YAMAGUCHI M, SAITO C, IKEUCHI M, KANNO K, KOJIMA S, HIRANO K, KITANO H, KASUGA S, ENDO T, FUKUDA H, MAKINO A. Growth properties and biomass production in the hybrid C4crop., 2016, 57(5): 944-952.
[8] Pfeiffer T W, Bitzer M J, Toy J J, Pedersen J F. Heterosis in sweet sorghum and selection of a new sweet sorghum hybrid for use in syrup production in appalachia., 2010, 50: 1788-1794.
[9] MINDAYE T T, MACE E S, GODWIN I D, JORDAN D R. Heterosis in locally adapted sorghum genotypes and potential of hybrids for increased productivity in contrasting environments in Ethiopia.,2016, 4: 479-489.
[10] 张福耀, 平俊爱, 赵威军. 中国酿造高粱品质遗传改良研究进展. 农学学报, 2019, 9(3): 21-25.
ZHANG F Y, PING J A, ZHAO W J. Genetic quality improvement of brewing sorghum in China: research progress., 2019, 9(3): 21-25. (in Chinese)
[11] GALICIA-JUAREZ M, MENDOZA-ONOFRE L E, GONZALEZ- HERNANDEZ V A, CISNEROS-LOPEZ M E, BENITEZ- RIQUELME I, CORDOVA-TELLEZ L. Heterosis and combining ability of seed physiological quality traits of single cross vs. three-way sorghum hybrids., 2017, 39(2): 175-181.
[12] PACKER D J, ROONEY W L. High-parent heterosis for biomass yield in photoperiod-sensitive sorghum hybrids., 2014, 167: 153-158.
[13] BIRCHLER J A, YAO H, CHUDALAYANDI S, VAIMAN D, VEITIA R A. Heterosis., 2010, 22: 2105-2112.
[14] REDDY B V S, RAMESH S, REDDY P S, RAMAIAH B. Combining ability and heterosis as influenced by male-sterility inducing cytoplasms in sorghum [(L.) Moench]., 2007, 154: 153-164.
[15] WEGARY D, VIVEK B, LABUSCHAGNE M. Association of parental genetic distance with heterosis and specific combining ability in quality protein maize., 2013, 191: 205-216.
[16] 曲玉杰, 孙君灵, 耿晓丽, 王骁, SARFRAZ Z, 贾银华, 潘兆娥, 何守朴, 龚文芳, 王立如, 庞保印, 杜雄明. 陆地棉亲本间遗传距离与杂种优势的相关性研究. 中国农业科学, 2019, 52(9): 1488-1500.
QU Y J, SUN J L, GENG X L, WANG X, SARFRAZ Z, JIA Y H, PAN Z E, HE S P, GONG W F, WANG L R, PANG B Y, DU X M. Correlation between genetic distance of parents and heterosis in upland cotton., 2019, 52(9): 1488-1500. (in Chinese)
[17] 韩东倩, 韩立朴, 薛帅, 尤明山, 谢光辉. 基于能源利用的高粱配合力和杂种优势分析. 中国农业大学学报, 2012, 17(1): 26-32.
HAN D Q, HAN L P, XUE S, YOU M S, XIE G H. Combining ability and heterosis of sorghum for biomass energy., 2012, 17(1): 26-32. (in Chinese)
[18] BUNPHAN D, JAISIL P, SANITCHON J, KNOLL J E, ANDERSON W F. Heterosis and combining ability of F1hybrid sweet sorghum in Thailand., 2015, 55: 178-187.
[19] UMAKANTH A V, PATIL J V, RANI C, GADAKH S R, KUMAR S S, RAO S S, KOTASTHANE T V. Combining ability and heterosis over environments for stalk and sugar related traits in sweet sorghum ((L.) Moench)., 2012, 14(3): 237-246.
[20] MAKANDA I, TONGOONA P, DERERA J, SIBIYA J, FATO P. Combining ability and cultivar superiority of sorghum germplasm for grain yield across tropical low- and mid-altitude environments., 2010, 116: 75-85.
[21] MAKANDA I, TONGOONA P, DERERA J. Combining ability and heterosis of sorghum germplasm for stem sugar traits under off-season conditions in tropical lowland environments., 2009, 114: 272-279.
[22] 侯荷亭, 杜志宏, 赵根弟. 高粱亲本遗传距离与杂种优势和特殊配合力的关系. 遗传, 1995, 17(1): 30-33.
HOU H T, DU Z H, ZHAO G D. Studies on the relationships of genetic distance of sorghum parental lines with heterosis and specific combining ability., 1995, 17(1): 30-33. (in Chinese)
[23] AMELEWORK B, SHIMELIS H, LAING M. Genetic variation in sorghum as revealed by phenotypic and SSR markers: implications for combining ability and heterosis for grain yield., 2016, 3: 1-13.
[24] 王瑞, 王金胜, 张福耀, 程庆军, 田承华, 凌亮. 1970s—2000s中国高粱杂交种亲本遗传距离演变的 SSR 分析. 中国农业科学, 2015, 48(3): 415-425.
WANG R, WANG J S, ZHANG F Y, CHENG Q J, TIAN C H, LING L. Evolution of genetic distance between parental lines of Chinese sorghum hybrids from1970s-2000s based on SSR analysis., 2015, 48(3): 415-425. (in Chinese)
[25] 陆平. 高粱种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006: 51-58.
LU P.[(L.) Moench]. Beijing: China Agricultural Press, 2006: 51-58. (in Chinese)
[26] WANG L M, JIAO S J, JIANG Y X, YAN H D, SU D F, SUN G Q, YAN X F, SUN L F. Genetic diversity analysis in parent lines of sweet sorghum based on agronomical traits and SSR markers., 2013, 149: 11-19.
[27] 孔繁玲. 植物数量遗传学. 北京: 中国农业大学出版社, 2006: 403-412.
KONG F L.. Beijing: China Agricultural University Press, 2006: 403-412. (in Chinese)
[28] NEI M, LI W. Mathematical model for studying genetic variation in terms of restriction endonucleases., 1979, 76(10): 5269-5273.
[29] 邹剑秋, 王艳秋, 张志鹏, 朱凯. A3型细胞质能源用甜高粱生物产量、茎秆含糖锤度和出汁率研究. 中国农业大学学报, 2011, 16(2): 8-13.
ZOU J Q, WANG Y Q, ZHANG Z P, ZHU K. Research on biomass, brix and juice extraction of A3-type cytoplasmic sweet sorghum for energy use., 2011, 16(2): 8-13. (in Chinese)
[30] 高士杰, 刘晓辉, 李玉发, 李继洪. 中国甜高粱资源与利用. 杂粮作物, 2006, 26(4): 273-274.
GAO S J, LIU X H, LI Y F, LI J H. Sweet sorghum resources and its utilization in China., 2006, 26(4): 273-274. (in Chinese)
[31] ZHANG C X, XIE G D, LI S M, GE L Q,HE T T. The productive potentials of sweet sorghum ethanol in China., 2010, 87: 2360-2368.
[32] SHUKLA S, FELDERHOFF T J, SABALLOS A, VERMERRIS W. The relationship between plant height and sugar accumulation in the stems of sweet sorghum ((L.) Moench)., 2017, 203: 181-191.
[33] KNOLL J E, ANDERSON W F, HARRIS-SHULTZ K R, NI X Z. The environment strongly affects estimates of heterosis in hybrid sweet sorghum., 2018, 20(3): 261-274.
[34] JAIKISHAN I, RAJENDRAKUMAR P, HARIPRASANNA K, BHAT B V. Gene expression analysis in sorghum hybrids and their parental lines at critical developmental stages in relation to grain yield heterosis by exploiting heterosis-related genes from major cereals., 2018, 36: 418-428.
[35] NDHLELA T, HERSELMAN L, SEMAGN K, MAGOROKOSHO C, MUTIMAAMBA C, LABUSCHAGNE M T. Relationships between heterosis, genetic distances and specific combining ability among CIMMYT and Zimbabwe developed maize inbred lines under stress and optimal conditions., 2015, 204: 635-647.
[36] PAVANI M, SUNDARAM R M, RAMESHA M S, KISHORE P B K, KEMPARAJU K B. Prediction of heterosis in rice based on divergence of morphological and molecular markers., 2018, 97(5): 1263-1279.
[37] 王林友, 张礼霞, 勾晓霞, 范宏环, 金庆生, 王建军. 利用InDel标记鉴定浙优系列杂交稻籼粳属性和预测杂种优势. 中国农业科学, 2014, 47(7): 1243-1255.
WANG L Y, ZHANG L X, GOU X X, FAN H H, JIN Q S, WANG J J. Identification ofattribute and prediction of heterosis of Zheyou hybrids rice using InDel molecular markers., 2014, 47(7): 1243-1255. (in Chinese)
[38] JORDAN D R, TAO Y, GODWIN I D, HENZELL R G, COOPER M, MCINTYRE C L. Prediction of hybrid performance in grain sorghum using RFLP markers., 2003, 106: 559-567.
[39] MELCHINGER A E, LEE M, LAMKEY K R, WOODMAN W L. Genetic diversity for restriction fragment length polymorphisms: relation to estimated genetic effects in maize inbreds., 1990, 30: 1033-1040.
[40] CHARCOSSET A, LEFORT-BUSEN M, GALLAIS A. Relationship between heterosis and heterozygosity at marker loci: a theoretical computation., 1991, 81: 571-575.
Heterosis Prediction of Sweet Sorghum Based on Combining Ability and Genetic Distance
WANG LiMing, YAN HongDong, JIAO ShaoJie, JIANG YanXi, SU DeFeng, SUN GuangQuan
(Crop Resource Institute of Heilongjiang Academy of Agricultural Sciences, Harbin 150086)
【】The heterosis, general combining ability (GCA) and special combining ability (SCA) of main agronomical traits of sweet sorghum were analyzed. Meanwhile, the feasibility of heterosis prediction based on combining ability, phenotypic genetic distance (PGD) and molecular genetic distance (MGD) was estimated. The result will provide theoretical reference for germplasm improvement and hybrid breeding of sweet sorghum.【】Sixty-four hybridized combinations were generated with 8 sterile lines as females and 8 restorer lines as males in accordance with a North Carolina Design II mating scheme. Twelve agronomical traits including days to flowering, growth duration, plant height, panicle length, stem diameter, tillers, panicle weight, 1000-grain weight, grain yield per plot, biomass per plant, biomass per plot and sugar content of hybrids and their parent lines were investigated in two years. The heterosis, GCA, SCA, PGD and MGD were analyzed, as well as the correlation between combining ability, genetic distance and heterosis. 【】Mid-parent heterosis (MPH) of traits from high to low was: biomass per plant, grain yield per plot, panicle weight, biomass per plot, plant height, panicle length, 1000-grain weight, stem diameter, growth duration, days to flowering, tillers and sugar content. Among which, heterosis of growth duration, days to flowering, tillers and sugar content was negative. The sequence of MPH for all traits was almost the same as that of better parent heterosis (BPH). Combining ability analysis showed that the GCA of different parents varied greatly in each trait. Meanwhile, the SCA of different combinations was also significantly different. Most combinations with high SCA also showed high GCA in their parent lines. Heterosis of biomass per plant, grain yield per plot, panicle weight, biomass per plot, panicle length, 1000-grain weight, tillers and sugar content was positively and highly significantly correlated with GCA and SCA of their parent lines simultaneously. Heterosis of growth duration was positively and highly significantly correlated with SCA. Meanwhile, heterosis of days to flowering was positively and significantly correlated with SCA. The PGD of parent lines was 2.86-6.82, and MGD was 0.50-0.96. The correlation between heterosis and MGD was greater than that of PGD in biomass per plant, grain yield per plot, panicle weight, biomass per plot, plant height, panicle length, stem diameter and sugar content. Among which, heterosis of biomass per plot, biomass per plant, panicle length and stem diameter was positively and highly significantly correlated with MGD. 【】Heterosis of yield related traits was higher, and that of sugar content and tillers was lower among all traits. In the prediction of heterosis, combining ability was more effective than genetic distance, therefore, can be used to predict heterosis. MGD was more effective in heterosis prediction compared with PGD.
sweet sorghum; heterosis; combining ability; genetic distance; prediction

10.3864/j.issn.0578-1752.2020.14.003
2019-07-09;
2019-09-20
国家现代农业产业技术体系(CARS-06-13.5-A1)、国家国际科技合作项目(2011DFR30840-10)
王黎明,Tel:0451-86668645;E-mail:dawnw@126.com。严洪冬,E-mail:hljcrop@163.com。王黎明和严洪冬为同等贡献作者
>
(责任编辑 李莉)
猜你喜欢
蔬菜(2021年7期)2021-11-27
养殖与饲料(2021年11期)2021-11-15
园艺与种苗(2021年8期)2021-09-23
北方果树(2016年1期)2016-12-17
食品与健康(2016年5期)2016-05-14
园艺与种苗(2015年8期)2015-12-28
科学大众(中学)(2015年11期)2015-12-09
江苏农业科学(2015年1期)2015-04-17
发明与创新·大科技(2009年5期)2009-05-31
商界(2000年7期)2000-06-14
