
不同糯性高粱胚乳淀粉形成与积累过程的酶学调控机制及显微结构变化
2020-08-15 02:49柯福来朱凯李志华石永顺邹剑秋王艳秋
中国农业科学 2020年14期
柯福来,朱凯,李志华,石永顺,邹剑秋,王艳秋
不同糯性高粱胚乳淀粉形成与积累过程的酶学调控机制及显微结构变化
柯福来,朱凯,李志华,石永顺,邹剑秋,王艳秋
(辽宁省农业科学院高粱研究所,沈阳 110161)
【】高粱是酿造白酒和食醋的主要原料,其淀粉组成和结构是影响高粱酿造品质的主要指标。通过研究淀粉积累过程中相关酶活性和淀粉粒超微结构的变化动态,解析高粱淀粉合成与积累的酶学调控机制,了解不同糯性高粱胚乳淀粉的超微结构特点,以期为酿造高粱的优质育种和栽培技术研究提供理论依据。以辽粘3号、辽杂19和辽杂10号3种不同糯性的高粱品种籽粒为研究对象,利用酶学及扫描电镜技术检测淀粉积累过程中相关酶活性的变化,观测淀粉积累过程中淀粉粒的形成过程及结构特征,利用相关分析研究淀粉合成过程中相关合成酶类的调控作用。淀粉合成过程中,不同胚乳类型高粱的尿苷二磷酸葡萄糖焦磷酸化酶(UDPG)、腺苷二磷酸葡萄糖焦磷酸化酶(ADPG)、可溶性淀粉合成酶(SSS)、结合态淀粉合成酶(GBSS)、淀粉分支酶(SBE)、淀粉去分支酶(DBE)活性均呈单峰曲线变化趋势;糯高粱结合态淀粉合成酶(GBSS)活性呈单峰曲线变化趋势,粳高粱、半粳半糯高粱则表现双峰曲线变化趋势。UDPG、ADPG、SSS活性与直链淀粉、支链淀粉积累速率显著正相关,SBE、DBE活性与支链淀粉积累速率呈显著正相关,GBSS活性与粳高粱、半粳半糯高粱直链淀粉积累速率呈显著正相关,与糯高粱直链淀粉积累速率表现正相关的趋势,但相关不显著。不同胚乳类型高粱淀粉粒的充实过程表现相似变化趋势,在开花后的14—35 d充实较快。糯高粱的淀粉粒较小,直径在10 µm以内,内部有圆孔型或楔形空洞。粳高粱淀粉粒较大,呈不规则球形,内部空洞极少。半粳半糯型高粱淀粉粒粒径分布较广,多数分布楔形或星形空洞,少部分无空洞。UDPG、ADPG、SSS是调控淀粉合成的关键酶,GBSS、SBE、DBE是调控淀粉组分比例的关键酶,SBE、DBE活性高是糯高粱支链淀粉含量高的主要原因,不同淀粉组分比例可能是影响淀粉粒结构的主要因素。
高粱;直链淀粉;支链淀粉;淀粉合成关键酶;显微结构
0 引言
【研究意义】高粱是中国主要的旱区作物,其籽粒主要用于酿造白酒和陈醋,尤其是名牌白酒和陈醋都是以高粱为原材料酿造而成。淀粉的含量及组分对于白酒和陈醋的品质和产量具有重要作用[1],因此,不同胚乳类型淀粉结构及调控研究将为高粱籽粒品质和产量提升提供理论基础。【前人研究进展】周宇飞等[2]以不同淀粉含量的高粱为研究对象,分析高粱淀粉形成过程中相关淀粉合成酶的调控作用,认为ADPG、GBSS、SBE和DBE活性的差异及共同作用对高粱淀粉积累差异具有重要生理调控作用。李栋梁等[3]采用扫描电镜、荧光显微镜、冰冻切片技术对高粱颖果发育过程中胚乳和淀粉体的形态进行了观察,发现高粱淀粉粒形成过程中存在不同于其他作物的发生中心结构。王玲玲等[4]采用光学显微镜和电子显微镜相结合的技术观察高粱胚乳细胞发育,重点研究了高粱胚乳细胞中淀粉体的发育情况,认为淀粉体是由质体通过充实淀粉形成的。闫松显等[5]以19份酿酒高粱为对象,对其果皮(果皮层和种皮层)厚度及淀粉粒进行显微结构研究,认为不同酿造高粱淀粉粒径差异显著。Huber等[6]通过对化学诱变剂诱变获得的消化率高、赖氨酸含量高的高粱淀粉进行扫描电镜观察,可以清楚地看到在一些淀粉颗粒表面有可观数量的孔。史春余等[7]、石德杨等[8]、杨勇等[9]在甘薯、玉米、水稻上的研究表明淀粉结构是影响作物品质的重要因素。【本研究切入点】与水稻、小麦、玉米等作物相比,关于高粱籽粒中淀粉合成机制的研究较少,对淀粉结构的相关研究更少,尤其是对高粱不同胚乳类型淀粉结构及调控的研究鲜见报道。【拟解决的关键问题】本研究以糯型、半粳半糯型和粳型3种不同类型的高粱品种为试材,研究不同类型高粱淀粉积累过程中相关酶类的变化,同时利用扫描电镜技术观察淀粉粒的形成、大小以及结构变化特点,揭示调控高粱籽粒淀粉积累及不同淀粉组分比率的内在因素,为高淀粉尤其是高支链淀粉高粱品种的选育和栽培提供理论基础。
1 材料与方法
1.1 供试材料
选择3种不同糯性的代表性高粱品种各1个(表1),其中,糯型品种为辽粘3号(辽粘A-2/辽粘R-2),半粳半糯型品种为辽杂19(7050A/0-01),粳型品种为辽杂10号(7050A/9198)。
1.2 试验方法
田间试验,每品种20行,行长3 m,行距0.6 m,3次重复,随机区组排列。2016年5月8日播种,开花期选择花期整齐一致的植株100株挂牌标记,用于取样测定。从开花后开始,每天取样一次,开花后7—42 d,每7天取样一次,每次取3株植株穗中部的籽粒混合,一部分样品150℃杀青30 min,75℃烘干至恒重;一部分籽粒用液氮迅速冷冻,-80℃保存;一部分籽粒用固定液固定后,利用酒精脱水保存,待全部样品取完后,统一测定。其中,参照赵俊晔等[10]方法测定尿苷二磷酸葡萄糖焦磷酸化酶(uridine diphosphateglucose pyrophosphorylase,UDPG)、腺苷二磷酸葡萄糖焦磷酸化酶(adenosine diphosphate glucosepyrophosphorylase,ADPG)、可溶性淀粉合成酶(soluble starch synthase,SSS)和结合态淀粉合成酶(granule- bound starch synthase,GBSS)的酶活性。按照Nakamura等[11]、Nakamura等[12]和赵法茂等[13]方法测定淀粉分支酶(starch branching enzyme,SBE)和淀粉去分支酶(starch-debranching enzymes,DBE)的酶活性。由上海釜诚生物技术有限公司利用扫描电镜测定淀粉粒结构,采用SPSS19.0进行数据分析。
2 结果
2.1 不同类型高粱籽粒淀粉积累过程中相关酶活性动态变化
开花后,籽粒开始灌浆。在籽粒灌浆过程中,UDPG、ADPG酶活性授粉后迅速上升,授粉后21 d达到最大值,其后UDPG酶活性略有下降,ADPG酶活性迅速下降(图1)。3个品种相比较,灌浆前期差异不大,灌浆中期差异显著,表现为辽杂10号>辽杂19>辽粘3号,灌浆后期UDPG酶活性差异仍达显著水平,ADPG酶活性差异不显著。
在高粱籽粒灌浆过程中,SSS酶活性表现和UDPG活性相似的变化趋势,均表现为授粉后迅速上升,而后略有下降,但SSS酶活性的峰值出现较早,在开花后15 d活性最高。整个灌浆过程中,辽粘3号SSS酶活性显著高于辽杂10号和辽杂19,辽杂10号与辽杂19间差异不显著。
SBE主要作用是水解α-1,4糖苷键,将切下的短链通过α-1,6糖苷键连接到受体上,从而形成分支淀粉链。不同类型高粱籽粒中SBE活性均呈单峰曲线变化,均在授粉后21 d达到峰值。整个灌浆过程中都表现为辽粘3号SBE活性显著高于辽杂19和辽杂10号,辽杂19和辽杂10号之间差异也达显著水平(图2)。
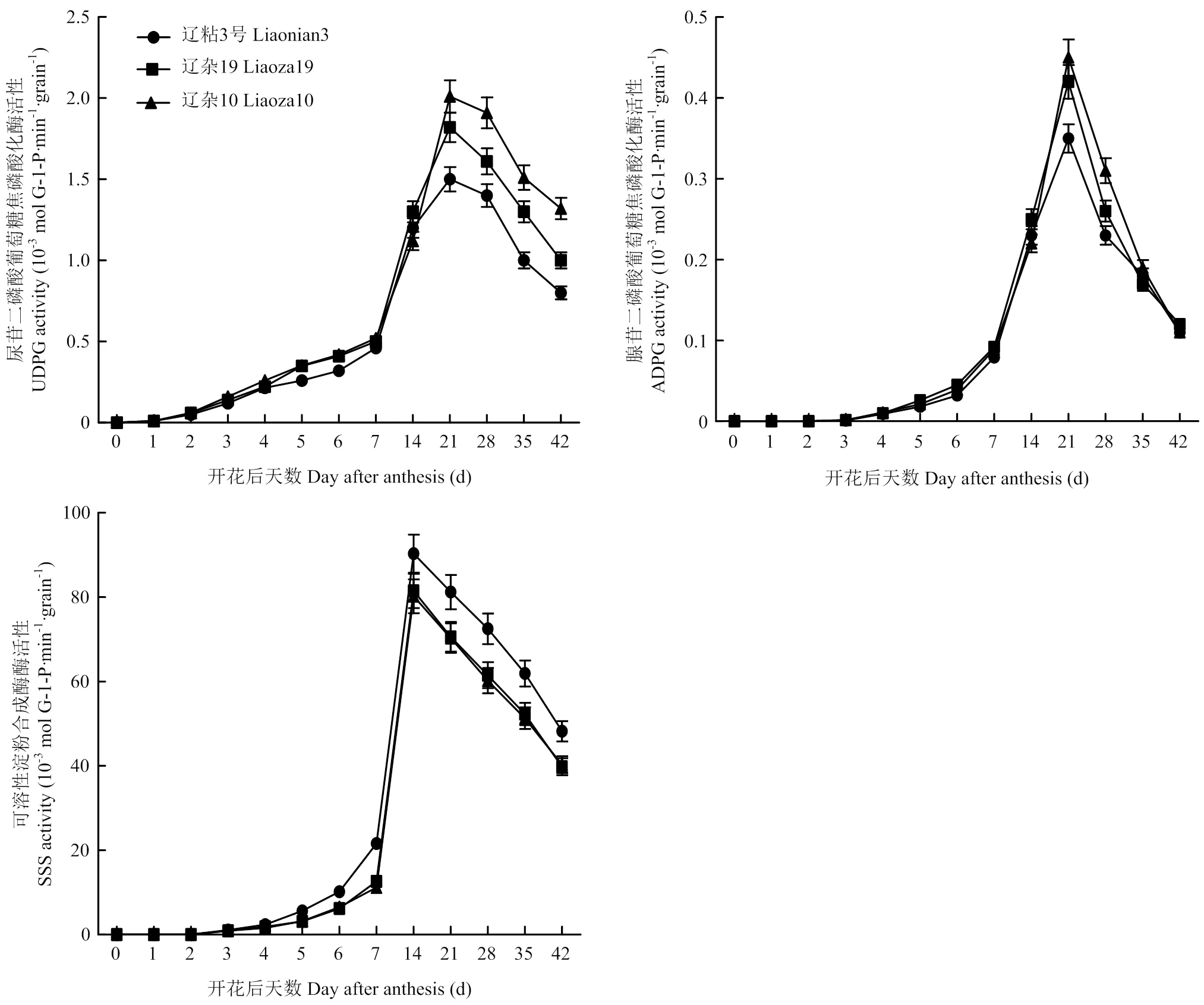
图1 UDPG、ADPG和SSS酶活性动态变化
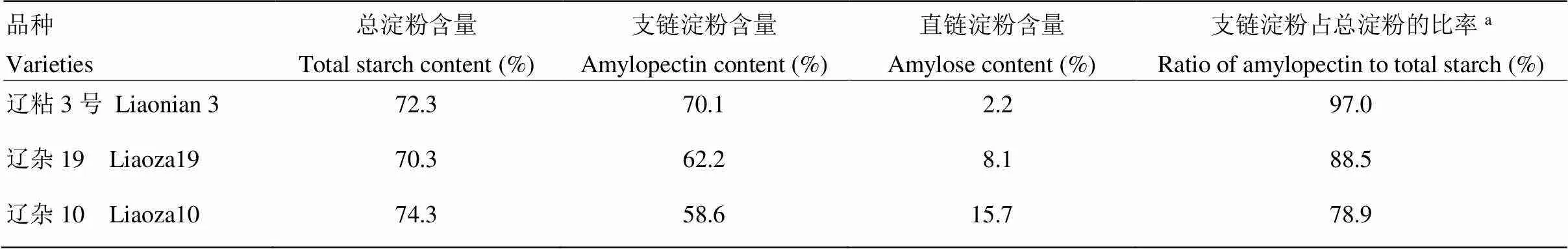
表1 参试品种籽粒的淀粉组成
a:国家高粱品种登记标准规定支链淀粉占总淀粉的比率超过90%(含90%)为糯高粱,低于80%(不含80%)为粳高粱
a: The standard of national sorghum variety registration stipulates that the ratio of amylopectin to total starch more than 90% (including 90%) is glutinous sorghum, less than 80% (excluding 80%) is nonwaxy sorghum
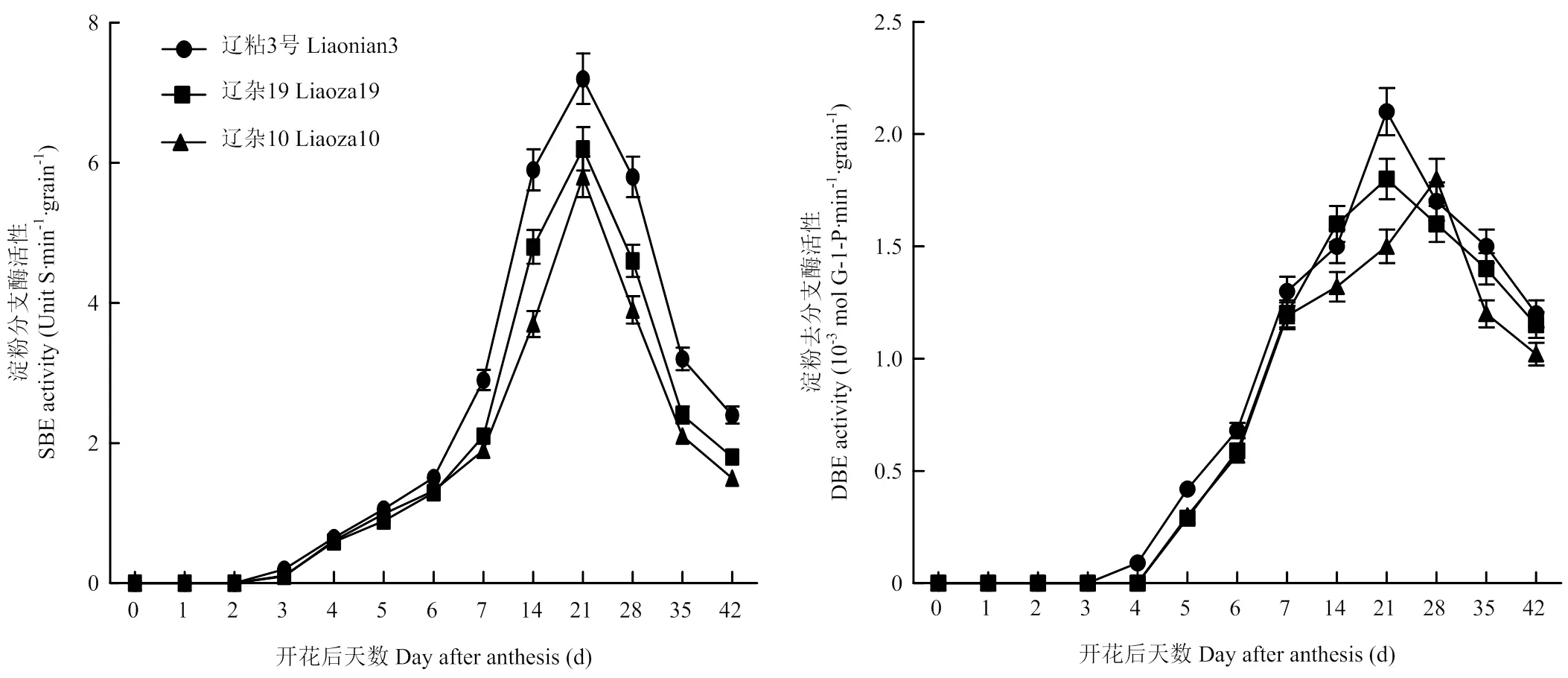
图2 SBE、DBE酶活性动态变化
DBE主要作用是水解a-1,6-糖苷键,对淀粉的最终结构进行修饰。由图2可见,不同类型高粱籽粒中DBE活性的变化也呈单峰曲线,糯高粱品种辽粘3号峰值最高,在授粉后21 d达到峰值;粳高粱和半粳半糯型高粱品种的峰值较低,粳型高粱达峰值的时间也较晚,辽杂10号在授粉后28 d籽粒中DBE活性达到峰值(图2)。
高粱籽粒灌浆过程中,GBSS酶活性波动较大,辽杂10号、辽杂19表现出明显的双峰曲线变化,峰值出现在授粉后15和35 d,且前期峰值较高;辽粘3号表现为单峰曲线,授粉后约20 d达到峰值。整个灌浆过程中,辽粘3号GBSS酶活性显著低于辽杂10号和辽杂19;辽杂10号与辽杂19间差异也达显著水平,除灌浆后期外,均表现为辽杂10号显著高于辽杂19(图3)。
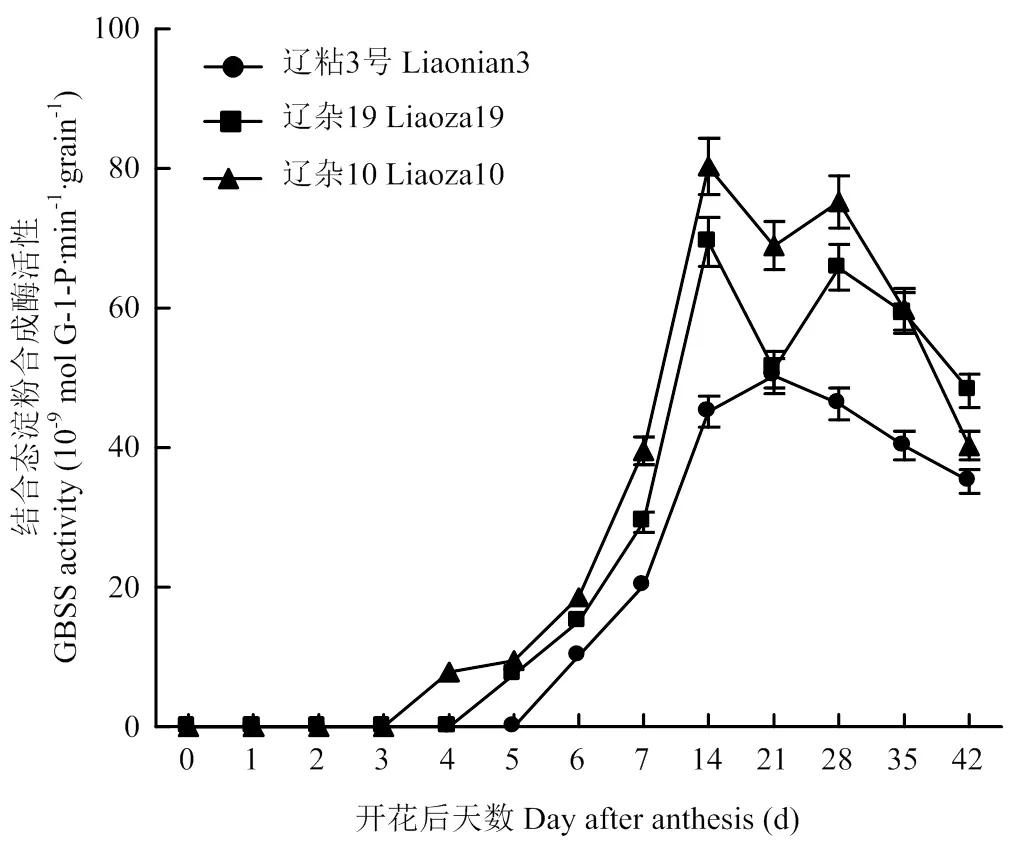
图3 不同类型高粱籽粒GBSS酶活性变化
2.2 淀粉合成相关酶活性与淀粉积累速率间相关分析
为了进一步明确淀粉合成相关酶类对淀粉合成的调控作用,对不同合成酶类与淀粉积累速率间的相关性进行了分析。结果(表2)表明,UDPG、ADPG和SSS酶活性和不同类型高粱直链淀粉、支链淀粉的积累速率均呈显著正相关。GBSS酶活性与辽杂19、辽杂10号直链淀粉积累速率显著正相关,与辽粘3号直链淀粉积累速率相关性不显著,与不同品种的支链淀粉积累速率相关性不显著。SBE、DBE活性与不同品种的支链淀粉积累速率显著正相关,与直链淀粉积累速率相关性不显著。

表2 不同类型高粱籽粒中淀粉合成相关酶活性与淀粉积累速率间相关分析
UDPG:尿苷二磷酸葡萄糖焦磷酸化酶;ADPG:腺苷二磷酸葡萄糖焦磷酸化酶;SSS:可溶性淀粉合成酶;GBSS:结合态淀粉合成酶;SBE:淀粉分支酶;DBE:淀粉去分支酶;*表示在0.05水平上相关显著
UDPG: Uridine diphosphate glucose pyrophosphorylase; ADPG: Adenosine diphosphate glucose pyrophosphorylase; SSS: Soluble starch synthase; GBSS: Granule-bound starch synthase; SBE: Starch branching enzyme; DBE: Starch-debranching enzymes; * mean significance of correlation coefficients at the 0.05 levels
2.3 淀粉超微结构的显微观察
2.3.1 胚乳细胞中淀粉粒的充实过程 粳高粱辽杂10号开花后7 d,胚乳细胞中已经出现淀粉粒积累,此时,淀粉粒个数较少,大多是圆形或卵圆形,个别细胞中已经出现淀粉粒合成中心(图4),在合成中心淀粉粒的体积较小,由合成中心向外,淀粉粒呈放射性分散,外围淀粉粒的体积逐渐增大。开花后14 d,部分胚乳细胞中淀粉粒充实较多,淀粉粒的积累速率加快,但仍有大部分细胞淀粉粒积累较少。开花后21 d,淀粉合成中心的数量显著增加,导致淀粉粒的积累速率明显增加,至花后28 d,大部分胚乳细胞中都可观察到淀粉粒积累,个别细胞中甚至可以观测到多个淀粉粒形成中心。开花后35 d,绝大部分细胞中都充满淀粉粒,淀粉粒之间可以观测到明显的蛋白体。开花后42 d,几乎所有细胞中都充满淀粉粒,淀粉粒形成中心的数量也显著下降。成熟期,由于淀粉粒的互相挤压,胚乳细胞中淀粉粒的外形由近圆形变为不规则的多边形,淀粉粒之间则充满蛋白体。
糯高粱辽粘3号胚乳细胞中淀粉粒的积累过程也表现相似的变化趋势,开花后7 d积累较少;开花后14—35 d淀粉粒的积累较多。淀粉粒最初的形状也呈近圆形,后期由于淀粉粒间互相挤压,也变为不规则的多边形。淀粉粒之间也充满蛋白体。
2.3.2 淀粉粒的形成中心 花后7 d在胚乳细胞中可观测到淀粉粒的分布较分散,只有少量淀粉粒,淀粉粒的直径以特定点为中心,呈现由大到小的变化趋势,在中心点存在点状呈簇的集合体,这就是淀粉粒形成中心的雏形。随着胚乳细胞的充实进程,淀粉粒形成中心逐渐显现,至开花后35 d,可以观测到淀粉粒以特定点为中心,呈现明显的轮状分布,内圈淀粉粒的直径较小,外圈淀粉粒的直径较大。花后45 d,形成中心处的淀粉粒互相堆叠挤压产生明显的形变(图5)。
2.3.3 淀粉粒的形态特征 不同胚乳类型高粱籽粒中,淀粉粒的形态特征表现为糯型淀粉粒相对较小,大部分均在10 μm以下,表面凹坑较多;粳型淀粉粒相对较大,大部分均在10 μm以上,不规则球形,表面比较光滑;半粳半糯型淀粉粒粒度分布较广,多为椭球形,表面凹坑较少(图6)。
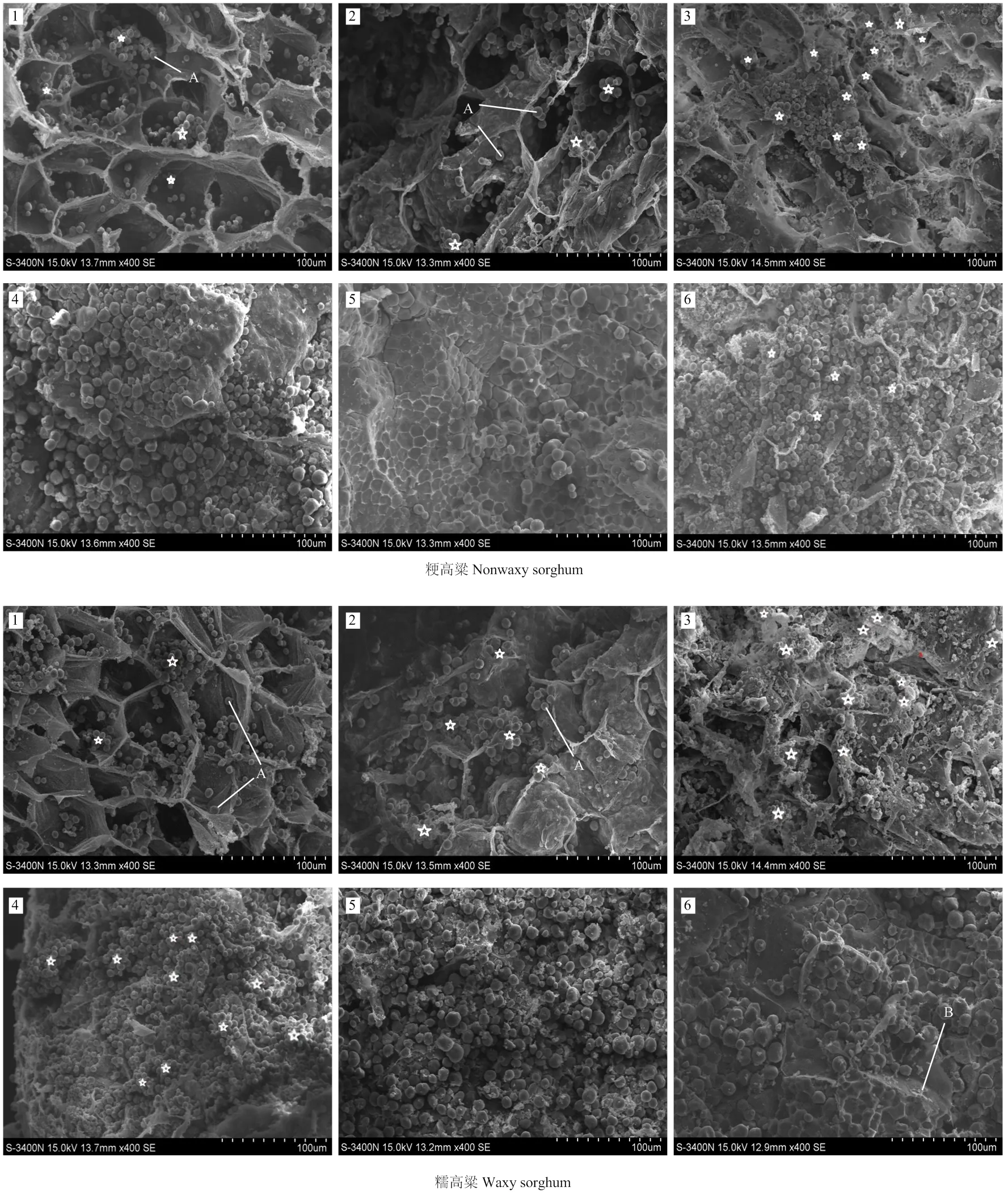
1:花后7 d;2:花后14 d;3:花后21 d;4:花后28 d;5:花后35 d;6:花后42 d;A:淀粉粒;☆:淀粉粒形成中心;B:蛋白体

1:35 d观测;2:7 d观测;3:42 d观测。A:淀粉粒;☆:淀粉粒形成中心;C:质体

1:糯高粱;2:半粳半糯高粱;3:粳型高粱 1: Glutinous sorghum; 2: Semi-glutinous sorghum; 3: japonica sorghum
2.3.4 淀粉粒内部结构特征 不同类型籽粒淀粉粒内部结构差异较大,糯型高粱籽粒淀粉粒内部分布较多圆形孔洞,平均直径约3.5 µm;粳型高粱淀粉粒内部比较光滑,未见明显孔洞,半粳半糯型高粱淀粉粒内部孔洞较少,且均呈不规则形状,孔洞最宽处在6—8 µm(图7)。

1:糯高粱;2:半粳半糯高粱;3:粳型高粱 1: Waxy sorghum; 2: Semi-waxy sorghum; 3: Nonwaxy sorghum
为了验证上述观测结果,又增加了8个不同类型的品种,并观测了它们的淀粉粒内部结构。粳型高粱淀粉粒内部只有极个别的出现圆形空洞,空洞的直径极小,深度也有限,有的仅是小的凹坑(图8-c—图8-e)。半粳半糯型高粱淀粉粒内部分布较多三角形、楔形或是星形的空洞(图8-a和图8-b)。糯型高粱淀粉粒内部也可观测到空洞的存在(图8-f—图8-h),空洞的形状多为楔形或圆形,在个别品种的淀粉粒表面也可观测到疑似孔洞的斑点(图8-h)。

a、b:半粳半糯型胚乳淀粉粒断面结构图;c、d、e:粳型胚乳淀粉粒断面结构图;f、g、h:糯型淀粉粒断面结构图
3 讨论
3.1 不同类型高粱淀粉合成的调控
籽粒淀粉的合成受相关合成酶类的调控,主要包括UDPG、ADPG、SSS、GBSS、SBE、DBE等。UDPG主要负责形成淀粉合成的葡萄糖供体G-1-P(1-磷酸葡萄糖),ADPG利用G-1-P合成淀粉合成的前体,因此这两种酶被认为是淀粉合成的限速酶[14-18]。本研究表明,不同类型高粱籽粒中UDPG、ADPG活性变化均呈单峰曲线,在开花后21 d酶活性最高,淀粉含量较高的辽杂10号2种酶的活性显著高于淀粉含量较低的2个品种,相关分析表明,不同类型品种的直链淀粉、支链淀粉的合成速率与其均呈显著正相关,表明在高粱淀粉合成过程中,UDPG、ADPG主要调控淀粉含量,对直链淀粉和支链淀粉的比例则没有显著作用,这与前人的研究结果基本一致[14-18]。
许多研究结果显示,支链淀粉含量主要由SSS、SBE和DBE 3种酶调控,其中SSS作用于α-1,4糖苷键,负责淀粉链的延长;SBE作用于α-1,6糖苷键,负责合成支链;DBE负责切除不合理的支链,修饰支链淀粉链的结构[19-20]。曹颖妮等[21]。盛婧[22]、王月福等[23]和赵步洪等[24]研究表明,SSS活性和支链淀粉合成显著正相关,Gao等[25]、Abel等[26]关于SSS基因突变体的研究也证明,SSS活性的下降使得支链淀粉的合成受到抑制,而直链淀粉的含量增加。本研究不同类型高粱SSS、SBE、DBE均表现单峰曲线变化趋势,SSS峰值出现较早,SBE、DBE峰值出现较晚,其中,粳高粱DBE峰值出现的时间明显晚于糯高粱。不同类型高粱SSS、SBE、DBE活性差异显著,糯高粱3种酶的活性始终高于粳高粱,这可能是糯高粱支链淀粉含量较高的主要原因。相关分析也表明,支链淀粉的积累速率与3种酶均表现显著正相关关系。
辽粘3号开花后5 d才检测到GBSS活性,晚于辽杂10号,其峰值出现的时间也晚于辽杂10号,而且整个灌浆过程中酶活性值均显著低于辽杂10号,这可能是其直链淀粉含量较低的主要因素。
直链淀粉合成过程中,其他酶类的作用也不能忽视。赵俊晔等[10]对9个不同直链淀粉含量的小麦的研究表明,SSS活性在灌浆后期与直链淀粉的积累负相关。曹颖妮等[21]研究小麦不同胚乳类型籽粒淀粉积累与酶活性的相关性,证明SSS、SBE活性与直链淀粉的合成速率显著正相关。胡阳阳等[27]也发现,GBSS、SBE活性和支链淀粉、直链淀粉积累速率均呈显著正相关。本研究发现,SSS活性与不同类型品种的直链淀粉和支链淀粉积累速率均呈显著正相关,SBE、DBE与直链淀粉的积累速率没有显著的相关,与前人的试验结果不尽相同,品种特性差异可能是造成这一结果的主要原因,这也从一个侧面说明SSS酶活性对淀粉籽粒的调控作用可能受品种特性的影响。依兵[28]对27份高粱籽粒淀粉调控研究也表明,SSS酶活性与淀粉合成的关系,受品种遗传因素影响。本研究中,GBSS活性与粳型高粱、半粳半糯型高粱直链淀粉积累速率显著正相关,与糯型高粱直链淀粉积累速率相关不显著。据此推测GBSS对直链淀粉的调控作用受底物含量的影响,也可能是糯高粱籽粒中较高的SBE与DBE活性抑制了GBSS的作用,导致GBSS活性对直链淀粉的调控作用受抑制。夏楠等[29]发现较高的SBE活性显著抑制了直链淀粉的形成。
淀粉合成相关酶活性受栽培技术措施的影响,研究表明,氮、磷、钾、灌水均影响淀粉合成酶类的活性[30-33],因此,在高粱栽培过程中,可采用适当栽培技术措施通过酶活性变化调控淀粉合成。
3.2 淀粉形成过程的显微结构变化
3.2.1 淀粉粒形成中心 李栋梁等[3]以KS-304为试验材料,利用电镜技术观测胚乳中淀粉形成过程,认为花后35 d,淀粉粒已经充斥细胞。本研究表明,花后35 d,细胞中仍可观测到淀粉粒形成中心,仍有淀粉粒的积累。王玲玲等[4]同样以KS-304为试验材料,利用电镜技术观测胚乳中淀粉的发育过程,认为淀粉粒是在淀粉粒合成中心由质体形成的,质体首先形成环膜结构,然后形成淀粉粒,淀粉粒的体积由形成中心向外呈辐射状增加。本研究也观测到淀粉粒形成中心的形成,淀粉粒的大小由形成中心向外呈轮式增加,但是没有观测到质体形成的环膜结构。
3.2.2 淀粉粒形成过程与淀粉积累间关系 通过显微技术,可以观测到淀粉粒的积累程度,开花后7 d已经观测到淀粉粒积累,开花后14 d淀粉粒积累量不多,开花后21—35 d可以观测到淀粉粒迅速增多,逐渐充满胚乳细胞,这与淀粉的积累过程一致,表明通过淀粉粒积累过程的显微观测可以直观的反映淀粉的积累情况。这与盛婧[34]的研究结果一致。
3.3 淀粉粒结构的显微观察
3.3.1 淀粉粒外部特征的显微观察 唐忠厚等[35]、赵米雪等[36]、史春余等[7]发现,不同来源淀粉粒的大小、形态各不相同。刘东莉[37]对不同链支比玉米淀粉颗粒特征的研究发现,玉米淀粉粒的大小受淀粉链支比的影响。余静等[38]观测了不同糯型小麦籽粒淀粉粒形态,发现糯型小麦淀粉粒平均粒径小于非糯型。本研究发现,不同胚乳类型高粱籽粒淀粉粒的大小差异显著,糯型高粱籽粒淀粉粒粒径较小,普遍在10 µm以下;粳型高粱淀粉粒较大,一般在15 µm左右;半粳半糯型高粱淀粉粒粒径分布较广。三类型淀粉粒形态相似,基本都为不规则的球形。
3.3.2 淀粉粒内部特征的显微观察 Huber等[6]通过共焦扫描激光显微镜观测高粱籽粒淀粉粒结构,发现淀粉粒表面存在微孔,这些微孔可能深入淀粉粒中心,并在淀粉粒的内部形成不同形状的空洞。本研究也发现淀粉粒内部存在空洞,但是空洞的有无及形状与胚乳类型相关,粳高粱淀粉粒中没有发现有空洞的分布,糯型高粱淀粉粒的空洞呈圆形,只有半粳半糯型高粱的淀粉粒中存在星形的空洞。多品种验证结果也证明,粳高粱淀粉粒内部空洞出现的几率很小,糯型高粱淀粉粒内部除了圆形的空洞外,也有较多楔形的空洞。本研究推测每个淀粉粒中直链与支链淀粉的比例可能不同,淀粉粒内部结构可能与直链/支链淀粉比值密切相关,直链淀粉占据比例高的淀粉粒内部形成空洞的几率较小,支链淀粉所占比例较高的淀粉粒内部形成空洞的几率较高,至于空洞的形状、大小的可能原因还需要进一步的研究。詹鹏杰等[39]研究表明,不同淀粉结构高粱的酿酒参数差异显著,糯高粱吸水速率快,糊化温度高,糯高粱淀粉粒内部较多的空洞可能是造成这一现象的主要原因。
4 结论
UDPG、ADPG和SSS是调控高粱籽粒淀粉积累量的关键酶。SBE、DBE是调控支链淀粉积累的关键酶。GBSS是调控直链淀粉积累的关键酶。
糯型高粱淀粉粒较小,粳型高粱淀粉粒较大。糯型高粱淀粉粒内部存在空洞,粳型高粱淀粉粒内部基本不存在空洞,半粳半糯型高粱淀粉粒内部空洞形状不规则。淀粉粒的大小及内部结构与直链淀粉/支链淀粉的比例密切相关。
[1] 丁国祥, 戴清炳, 曾庆曦. 不同淀粉结构高粱籽粒的酿酒工艺参数研究. 绵阳农专学报, 1996, 13(4): 4-5.
DING G X, Dai Q B, ZENG Q X. A study on technological parameters for brewing wine from sorghum grains with different starch structure., 1996, 13(4): 4-5. (in Chinese)
[2] 周宇飞, 依兵, 吴奇, 张姣, 孙飞, 赵子川, 许文娟, 黄瑞冬. 不同淀粉含量高粱子粒淀粉积累差异及其生理基础//2016年全国青年作物栽培与生理学术研讨会, 2016.
ZHOU Y F, YI B, WU Q, ZHANG J, SUN F, ZHAO Z C, XU W J, HUANG R D. Difference of starch accumulation in sorghum seeds with different starch content and its physiological basis//, 2016(in Chinese)
[3] 李栋梁, 荆彦平, 李小刚, 顾蕴洁, 王忠. 高粱胚乳细胞与母体组织发育关系的研究. 中国农业科学, 2014, 47(17): 3336-3347.
LI D L, JING Y P, LI X G, GU Y J, WANG Z. Investigation of sorghum endosperm cell development and the relationship with its maternal tissue., 2014, 47(17): 3336-3347. (in Chinese)
[4] 王玲玲, 王忠, 顾蕴洁. 高粱胚乳淀粉体发育研究. 安徽农业科学, 2017, 45(30): 28-31, 35.
WANG L L, WANG Z, GU Y J. Investigation of amyloplast development in sorghum endosperm cells., 2017, 45(30): 28-31, 35. (in Chinese)
[5] 闫松显, 袁河, 雷元春, 赵亮, 罗汝叶, 倪德让, 王莉. 酿酒高粱籽粒微观形态分析及其果皮厚度和单宁含量的相关性. 中国酿造, 2018, 37(3): 67-71.
YAN S X, YUAN H, LEI Y C, ZHAO L, LUO R Y, NI D R, WANG L. Micro-morphology analysis of liquor-making sorghum grain and the correlation of pericarp thickness and tannin content., 2018, 37(3): 67-71. (in Chinese)
[6] Huber K C, Bemiller J N., 2000, 41: 269-276.
[7] 史春余, 姚海兰, 张立明, 柳洪鹃, 张超, 刘桂玲. 不同类型甘薯品种块根淀粉粒粒度的分布特征. 中国农业科学, 2011, 44(21): 4537-4543.
SHI C Y, YAO H L, ZHANG L M, LIU H J, ZHANG C, LIU G L. Starch granule size distribution in storage roots of different types of sweet potato cultivars., 2011, 44(21): 4537-4543. (in Chinese)
[8] 石德杨, 张海艳, 董树亭. 补充灌溉和施氮对玉米籽粒淀粉粒粒度分布的影响. 中国农业科学, 2014, 47(4): 633-643.
SHI D Y, ZHANG H Y, DONG S T. Effects of supplemental irrigation and nitrogen application on starch granule size distribution of maize grain., 2014, 47(4): 633-643. (in Chinese)
[9] 杨勇, 陆彦, 郭淑青, 石仲慧, 赵杰, 范晓磊, 李钱峰, 刘巧泉, 张昌泉. 籼稻背景下导入Wxin等位基因改良稻米食味和理化品质. 作物学报, 2019, 45(11): 1628-1637.
YANG Y, LU Y, GUO S Q , SHI Z H, ZHAO J, FAN X L, LI Q F , LIU Q Q, ZHANG C Q. Improvement of rice eating quality and physicochemical properties by introgression of Wxinallele invarieties, 2019, 45(11): 1628-1637. (in Chinese)
[10] 赵俊晔, 于振文, 孙慧敏, 马兴华, 孙强. 不同小麦品种籽粒淀粉组分及相关酶活性的差异. 作物学报, 2004, 30(6): 525-530.
ZHAO J Y, YU Z W, SUN H M, MA X H, SUN Q. Differences in starch components and related enzymes activity in the grains of different wheat cultivars., 2004, 30(6): 525-530. (in Chinese)
[11] Nakamura Y, YuKi k, Park S. Carbohydrate metabolism in the developing endosperm of rice grains., 1989, 30: 833-839.
[12] Nakamura Y, Umemoto T, Ogata N, Kuboki Y, Yano M, Sasaki T. Starch debranching enzyme (R-enzyme or pullulanase) from developing rice endosperm: purification, cDNA and chromosomal localization of the gene., 1996, 199: 209-218.
[13] 赵法茂, 齐霞, 肖军, 王宪泽. 测定淀粉分支酶活性方法的改进. 植物生理学通讯, 2007, 43(6): 1167-1169.
ZHAO F M, QI X, XIAO J, WANG X Z. Improvemed method for determining starch branching enzyme activity., 2007, 43(6): 1167-1169. (in Chinese)
[14] 胡博文, 谷娇娇, 贾琰, 沙汉景, 张君颜, 黄书勤, 赵宏伟. 盐胁迫对寒地粳稻籽粒淀粉形成积累及产量的影响. 华北农学报, 2019, 34(1): 115-123.
HU B W, GU J J, JIA Y, SHA H J, ZHANG J Y, HUANG S Q, ZHAO H W. Effect of salt stress on starch formation and yield ofrice in cold-region., 2019, 34(1): 115-123.(in Chinese)
[15] 王自布, 李卫华, 齐军仓, 银永安, 曹连莆, 王泽民, 侯睿睿, 王亮. 小麦籽粒胚乳淀粉合成酶基因表达及酶活性分析. 核农学报, 2010, 24(6): 1117-1123.
WANG Z B, LI W H, QI J C, YIN Y A, CAO L P, WANG Z M, HOU R R, WANG L. Analysis of gene expression of enzymes involved in starch synthesis and enzymes activity in wheat grain., 2010, 24(6): 1117-1123. (in Chinese)
[16] 赵宏伟, 赵景云. 钾素用量对春玉米淀粉合成酶活性及产量的影响. 玉米科学, 2010, 18(3): 140-143, 146.
ZHAO H W, ZHAO J Y. Effects of potassium application on enzyme activity of starch synthesis and yield in spring maize., 2010, 18(3): 140-143, 146. (in Chinese)
[17] 田云录, 陈金, 董文军, 邓艾兴, 张卫建. 非对称性增温对冬小麦强势粒和弱势粒淀粉合成关键酶活性的影响. 作物学报, 2011, 37(6): 1031-1038.
TIAN Y L, CHEN J, DONG W J, DENG A X, ZHANG W J. Effects of asymmetric warming on key enzyme activities of starch synthesis in superior and inferior grains of winter wheat under FATI facility., 2011, 37(6): 1031-1038. (in Chinese)
[18] 周达, 杨璞, 李夏, 赵伟杰, 周瑜, 杨清华, 晁桂梅, 冯佰利. 水分胁迫下糜子籽粒淀粉合成关键酶及与灌浆特性的关联. 西北农业学报, 2015, 24(9): 63-71.
ZHOU D, YANG P, LI X, ZHAO W J, ZHOU Y, YANG Q H, CHAO G M, FENG B L. Activities of key enzymes for starch synthesis and grain-filling characteristics under water stress in broomcorn millet., 2015, 24(9): 63-71. (in Chinese)
[19] 李瑞清, 谭瑗瑗, 闫影, 张丽霞, 曹黎明, 吴书俊. 水稻胚乳淀粉合成及其育种应用. 核农学报, 2019, 33(9): 1742-1748.
LI R Q, TAN Y Y, YAN Y, ZHANG L X, CAO L M, WU S J. Starch biosynthesis in rice endosperm and its applications in breeding., 2019, 33(9): 1742-1748. (in Chinese)
[20] 李强, 李涛, 白玉婷, 任芹勇, 郭世华. 粳谷和糯谷胚乳淀粉合成酶活性的动态变化. 分子植物育种, 2019: 1-11.
LI Q, LI T, BAI Y T, REN Q Y, GUO S H. Dynamic changes of endosperm starch synthetase activity in no-waxy and waxy foxtail millets., 2019: 1-11. (in Chinese)
[21] 曹颖妮, 胡卫国, 王根平, 刘录祥, 王成社. 糯性和非糯性小麦灌浆期胚乳直/支链淀粉积累及其相关酶活性研究. 西北植物学报, 2010, 30(10): 1995-2001.
CAO Y N, HU W G, WANG G P, LIU L X, WANG C S. Dynamic changes of starch accumulation and enzymes relating to starch biosynthesis of kernel during grain filling in waxy and non-waxy winter wheat., 2010, 30(10): 1995-2001. (in Chinese)
[22] 盛婧. 不同类型专用小麦籽粒淀粉形成及其与加工品质的关系[D]. 扬州: 扬州大学, 2004.
SHENG J. Starch development and its relationship with processing quality in wheat varieties for different end uses[D]. Yangzhou: Yangzhou University, 2004. (in Chinese)
[23] 王月福, 于振文, 李尚霞, 余松烈. 小麦籽粒灌浆过程中有关淀粉合成酶的活性及其效应. 作物学报, 2003, 29(1): 75-81.
WANG Y F, YU Z W, LI S X, YU S L. Activity of enzymes related to starch synthesis and their effect during the filling of winter wheat., 2003, 29(1): 75-81. (in Chinese)
[24] 赵步洪, 张文杰, 常二华, 王志琴, 杨建昌. 水稻灌浆期籽粒中淀粉合成关键酶的活性变化及其与灌蜘墓辫口蒸煮品质的关系. 中国农业科学, 2004, 37(8): 1123-1129.
ZHAO B H, ZHANG W J, CHANG E H, WANG Z Q, YANG J C. Changes in activities of the key enzymes related to starch synthesis in rice grains during grain filling and their relationships with the filling rate and cooking quality., 2004, 37(8): 1123-1129. (in Chinese)
[25] GAO M, Chibbar R N. Molecular and biochemical characterization of soluble starch syntheses in wheat kernels//. Saskatchewan: University Extension Press, 1998(4): 151-153.
[26] Abel G J W, Springer F, Willmitzer L,KOSSMANN J. Cloning and functional analysis of a cDNA encoding a novel 139-kda starch synthase from potato (SL.)., 1996(10): 981-991.
[27] 胡阳阳, 卢红芳, 刘卫星, 康娟, 马耕, 李莎莎, 褚莹莹, 王晨阳. 灌浆期高温与干旱胁迫对小麦籽粒淀粉合成关键酶活性及淀粉积累的影响. 作物学报, 2018, 44(4): 591-600.
HU Y Y, LU H F, LIU W X, KANG J, MA G, LI S S, CHU Y Y, WANG C Y. Effects of high temperature and water deficiency during grain filling on activities of key starch synthesis enzymes and starch accumulation in wheat, 2018, 44(4): 591-600. (in Chinese)
[28] 依兵. 高粱子粒淀粉积累与合成相关酶活性研究[D]. 沈阳: 沈阳农业大学, 2014.
YI B. Starch Accumulation and enzymes activities for starch synthesis in sorghum grains[D]. Shenyang: Shenyang Agricultural University, 2014. (in Chinese)
[29] 夏楠, 赵宏伟, 吕艳超, 赵振东, 邹德堂, 刘化龙, 王敬国, 贾琰. 灌浆结实期冷水胁迫对寒地粳稻籽粒淀粉积累及相关酶活性的影响. 中国水稻科学, 2016, 30(1): 62-71.
XIA N, ZHAO H W, Lü Y C, ZHAO Z D, ZOU D T, LIU H L, WANG J G, JIA Y. Effect of cold-water stress at filling stage on related enzyme activities in drains of japonica rice starch accumulation and in cold-region., 2016, 30(1): 62-71. (in Chinese)
[30] 王劲松, 董二伟, 武爱莲, 白文斌, 王媛, 焦晓燕. 不同费力条件下施肥对粒用高粱产量、品质及养分吸收利用的影响. 中国农业科学, 2019, 52(22): 4166-4176.
WANG J S, DONG E W, WU A L, BAI W B, WANG Y, JIAO X Y. Responses of fertilization on sorghum grain yield, quality and nutrient utilization to soil fertility., 2019, 52(22): 4166-4176. (in Chinese)
[31] 邹铁祥, 戴廷波, 姜东, 荆奇, 曹卫星. 氮素和钾素对小麦籽粒淀粉合成关键酶活性的影响. 中国农业科学, 2008, 41(11): 3858-3864.
ZOU T X, DAI T B, JIANG D, JING Q, CAO W X. Effects of nitrogen and potassium on key regulatory enzyme activities for grain starch in winter wheat., 2008, 41(11): 3858-3864. (in Chinese)
[32] 李春艳, 张润琪, 付凯勇, 李超, 朱长安, 李诚. 小麦淀粉合成关键酶基因和相关蛋白表达对不同施磷量的响应. 麦类作物学报, 2018, 38(4): 401-409.
LI C Y, ZHANG R Q, FU K Y, LI C, ZHU C A, LI C. Effect of phosphorus on the gene and protein expression related to starch biosynthesis in wheat., 2018, 38(4): 401-409. (in Chinese)
[33] 柳洪鹃, 姚海兰, 史春余, 张立明. 施钾时期对甘薯济徐23块根淀粉积累与品质的影响及酶学生理机制. 中国农业科学, 2014, 47(1): 43-52.
LIU H J, YAO H L, SHI C Y, ZHANG L M. Effect of potassium application time on starch accumulation and related enzyme activities of sweet potato variety Jixu 23., 2014, 47(1): 43-52. (in Chinese)
[34] 盛婧, 郭文善, 朱新开, 封超年, 彭永欣. 不同专用类型小麦籽粒淀粉粒形成过程. 作物学报, 2004, 30(9): 953-954.
SHENG J, GUO W S, ZHU X K, FENG C N, PENG Y X. Starch granules development in grain endosperm of wheat for different end uses., 2004, 30(9): 953-954. (in Chinese)
[35] 唐忠厚, 张爱君, 陈晓光, 靳容, 刘明, 李洪民, 丁艳锋. 低钾胁迫对甘薯块根淀粉理化特性的影响及其基因型差异. 中国农业科学, 2017, 50(3): 513-525.
TANG Z H, ZHANG A J, CHEN X G, JIN R, LIU M, LI H M, DING Y F.Starch physico-chemical properties and their difference in three sweet potato ((L.) Lam) genotypes under low potassium stress., 2017, 50(3): 513-525. (in Chinese)
[36] 赵米雪, 包亚莉, 刘培玲. 淀粉颗粒微观精细结构研究进展. 食品科学, 2018, 39(11): 284-294.
ZHAO M X, BAO Y L, LIU P L. Progress in research on fine microstructure of starch granules., 2018, 39(11): 284-294. (in Chinese)
[37] 刘东莉. 不同链支比玉米淀粉颗粒结构原位表征与分析[D]. 杭州: 浙江大学, 2014.
LIU D L. In situ characterization and analysis of the structures of maize starches with different amylose/amylopectin ratios[D]. Hangzhou: Zhejiang University, 2014. (in Chinese)
[38] 余静, 冉从福, 李学军, 邵慧, 李立群. 陕糯1号与非糯小麦西农1330胚乳发育及淀粉形态、粒径分析. 中国农业科学, 2014, 47(22): 4405-4416.
YU J, RAN C F, LI X J, SHAO H, LI L Q. Study on endosperm development and morphological features of starch granules in waxy wheat shannuo1 and non-waxy wheat Xinong 1330., 2014, 47(22): 4405-4416. (in Chinese)
[39] 詹鹏杰, 张福耀, 王瑞, 赵蜻, 于纪珍, 李燕. 不同淀粉类别高粱品种酿酒相关性能分析. 山西农业科学, 2013, 41(9): 897-898, 952.
ZHAN P J, ZHANG F Y, WANG R, ZHAO Q, YU J Z, LI Y. Analysis on brewing properties of sorghum with different starch categories., 2013, 41(9): 897-898, 952. (in Chinese)
Formation Regulating and Micro-structure of sorghum starch with different types of endosperm
KE FuLai, ZHU Kai, Li ZhiHua, SHI YongShun, ZOU JianQiu, WANG YanQiu
(Sorghum research institute, Liaoning Academy of Agricultural Sciences, Shenyang 110161)
【】 Sorghum is the main material for liquor-making and brewing well-aged vinegar, and its starch composition and structure are the main factors that influence the brewing quality of sorghum. In this paper, the Dynamic Changes of Key Enzymes Involved in Starch Synthesis are analyzed, the ultrastructure of starch is observed, for confirming the regulatory effect of related enzymes on sorghum starch synthesis, and revealing the ultrastructure characteristics of sorghum starch.【】Three sorghum cultivars(Liaonian3, Liaoza19, Liaoza10) with different endosperm types were used as experimental materials. The changes of key enzymes involved in starch synthesis were detected with enzymological technique. Scanning electron microscopy was used to observe the morphology features of starch granules. The regulation of related enzymes in starch synthesis was studied by correlation analysis. 【】 The activity of UDPG, ADPG, SSS, SBE, DBE enzymes of sorghum with different endosperm types showed a single peak curve during starch accumulation. The activity of GBSS enzyme of sorghum with waxy endosperm showed a single peak curve, while other types sorghum showed bimodal curves. Activities of UDPG and ADPG, SSS enzyme were significantly correlated with amylose and amylopectin accumulation rate. Activities of SBE, DBE enzyme were positively correlated with amylopectin accumulation rate. Activities of GBSS enzyme was positively and significantly correlated with amylopectin accumulation rate in non-waxy sorghum and semi-waxy sorghum, it showed a positively correlation trend, but not significant in waxy sorghum. For sorghum with different endosperms, the formation process of starch grains showed a similar tendency. The starch accumulation rate was higher during 14-35 d after flowering. The starch grains of waxy sorghum are the smallest, with a diameter within 10 µm, and carved with round-hole or wedge-shaped cavity inside. The starch grains of nonwaxy sorghum are the largest, irregularly globular, with scarcely cavity inside. The size distribution of sorghum starch granules with semi-waxy endosperm ranges widely, most of them with wedge-shaped or star-shaped cavities inside, only a few with no cave. 【】 UDPG, ADPG, SSS play important adjustive roles in the accumulation of amylopectin and amylose. GBSS, SBE and DBE are the key enzymes to regulate the ratio of amylose/amylopectin. Higher activities of SBE and DBE are the main reason that makes waxy sorghum contains higher amylopectin. The ratio of amylose/amylopectin may be the main factor affecting the starch grain structure of sorghum.
L.; amylose; amylopectin; starch synthesis related enzymes; microstructure

10.3864/j.issn.0578-1752.2020.14.002
2019-07-31;
2020-01-02
国家重点研发计划(2019YFD1001704/2019YFD1001700)、国家谷子高粱产业技术体系(CARS-06-13.5-A22)、中央引导地方项目(2018010035-201)、辽宁省农业领域青年人才培养计划(2014019)
柯福来,E-mail:397634262@qq.com。通信作者王艳秋,E-mail:wangyanqiu73@126.com
(责任编辑 李莉)
猜你喜欢
包装工程(2022年1期)2022-01-26
农业机械学报(2021年10期)2021-11-09
中国农业科学(2021年7期)2021-04-21
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
农产品加工(2020年3期)2020-03-11
北京航空航天大学学报(2019年3期)2019-04-08
小学阅读指南·高年级版(2016年9期)2016-10-31
安徽农业科学(2015年9期)2015-01-12
食品工业科技(2014年13期)2014-12-16
