
欧洲云杉PSⅡ的热稳定性对温度升高的响应*
2020-08-14 02:30贾子瑞马建伟安三平胡继文王军辉
林业科学 2020年7期
贾子瑞 王 亚 马建伟 安三平 胡继文 王军辉
(1. 中国林业科学研究院林业研究所 北京 100091; 2.林木遗传育种国家重点实验室 北京 100091; 3.甘肃省小陇山林业科学研究所 天水741020; 4. 甘肃省次生林培育重点实验室 天水 741020)
气候变暖显著影响温带、寒温带和高山森林的生长发育和健康(Leemansetal., 2004; Kauretal., 2017; Ponocnáetal., 2016)。根据气候变化对物种、生态系统和陆地景观以及相应的温度和降水模式的影响,温带和北方森林对气候变化最为敏感,尤其温带和寒温带针叶林更易受到显著影响(Leemansetal., 2004)。气候变暖对寒温带地区树木的影响依树种不同而异,如对松(Pinus)和桦(Betula)有利,对冷杉(Abies)的影响不大,对云杉(Picea)和山毛榉(Fagus)则不利(Carreretal., 2012; Takahashietal., 2013; Camareroetal., 2015; Ziacoetal., 2016; Franketal., 2017; Kolaretal., 2017; Ponocnáetal., 2016)。云杉属全世界约35种,主要分布于北半球的亚热带高海拔、温带和北方地区(郑万钧等,1978; Tayloretal., 1994),我国有18种,其中7个特有种,2个引进栽培种; 欧洲云杉(P.abies)在我国引种后成功驯化,其适应性和长势超过了许多乡土云杉种(李青粉等, 2012; 祁万宜等, 2006)。但作为北半球一种重要的经济生态树种,正面临气候变化产生的威胁(Tjoelkeretal., 2007; Bollietal., 2007)。
高温胁迫是影响植物光合作用的重要因素之一,叶绿体光合活性是高等植物的热敏感部位,很容易被热胁迫摧毁破坏。 光合系统的PSⅡ、PS I、电子传递链、ATP合成、碳固定等其他组分对热胁迫都很敏感,尤其PSⅡ更易被高温摧毁(Ashrafetal., 2013、Mathuretal., 2014)。因此,研究在热胁迫下PSⅡ的热稳定性,对促进植物适应未来气候变暖十分必要。在应用叶绿素荧光对树木PSⅡ热激反应的光合效率方面已有很多研究,例如: 短叶雪松(Cedrusbrevifolia)、黎巴嫩雪松(C.libani)和大西洋雪松(C.atlantica)(Ladjaletal., 2000),橡树(Quercuspalustris)(Daasetal., 2008),花旗松(Pseudotsugamenziesii)和西部黄松(P.ponderosa)(Mariasetal., 2016), 欧洲冷杉(A.alba)(Konopkovaetal., 2018)等。目前,已对分布在中国的云杉属红皮云杉(P.koraiensis)、青木千(P.wilsonii)、青海云杉(P.crassifolia)、粗枝云杉(P.asperata)、紫果云杉(P.purpurea)、川西云杉(P.likiangensisvar.rubescens)、西藏云杉(P.spinulosa)、林芝云杉(P.likiangensis. var.linzhiensis)(杨毅, 2017)9个树种的PSⅡ热稳定性进行了研究,但对欧洲云杉PSⅡ的热稳定性未展开系统的研究。
本研究以5年生欧洲云杉幼苗为对象,应用hand-PEA和image-PAM检测其针PSⅡ热反应,并根据前人对松科植物热激胁迫下PSⅡ热稳定性的研究,从40~54 ℃设定10个热激胁迫的温度梯度,来研究欧洲云杉PSⅡ的初始胁迫温度和极限胁迫温度以及热损伤的敏感部位,为探讨欧洲云杉在未来气候变暖下的适应能力提供参考。
1 研究方法
1.1 试验材料
选择3个欧洲云杉无性系的5年生幼苗、每个无性系10个分生株,共计30个分生株,放入人工气候室培养。培养条件是: 白天/夜间18/23 ℃变温,光密度是640 μmol·m-2s-1,空气相对湿度是40%,CO2浓度是370 μmol·mol-1。
1.2 热激处理前对照设置
在人工气候室培养2周的幼苗,应用hand-PEA 和image-PAM检测叶片的荧光参数和影像作为热激处理前的对照,记为CG。
1.3 热激临界温度的选择
在热激处理组(记为HS)中,将在人工气候室培养过2周的3个无性系幼苗放入38 ℃培养箱中,在低光条件下预热5 h(预热能增强云杉属植物的耐热性),光照强度215 μmol·m-2s-1,CO2浓度是370 μmol·mol-1,空气相对湿度40%。然后在40、42、43、44、46、 48、 50、 52、53和 54 ℃下分别热激30 mins,每个处理3个无性系(每个无性系1个分生株)。应用image-PAM检测荧光影像和PSⅡ的光化学能Y(Ⅱ),热耗散Y(NPQ)和荧光淬灭Y(NO)。选择欧洲云杉幼苗的初始胁迫温度、极限胁迫温度2个阈值。
1.4 热激后PSⅡ自行修复的测定
在热激后的恢复组(记为RT)中,将热激后选择的2个临界温度处理的幼苗再次放入人工气候室正常条件下培养2周(见1.1)。培养2周后的幼苗,用hand-PEA 和image-PAM检测幼苗叶片的荧光参数和影像。研究欧洲云杉PSⅡ热激后的自我修复能力。
1.5 荧光参数测定
1.5.1 热激临界温度选择 应用image-PAM检测PSⅡ的光化学能Y(Ⅱ),热耗散Y(NPQ)和荧光淬灭Y(NO)的荧光影像、参数和动力学曲线,用来筛选初始和极限胁迫温度,其中:

Y(NPQ)=1-Y(Ⅱ)-1/[NPQ+1+qL(Fm/F0-1)],式中,Y(NPQ)为PSⅡ处调节性能量耗散的量子产量。
Y(NO)=1/(NPQ+1+qL(Fm/F0-1)),Y(NO)为非调节性能量耗散的量子产量,且Y(Ⅱ)+Y(NPQ)+Y(NO)=1。
1.5.2 热激临界温度下PSⅡ荧光参数测定 应用Image-PAM检测PSⅡ的荧光参数Fm、Fv/Fm、NPQ、qL和qP的影像,荧光参数NPQ、qL和qP动力学曲线用来评估44和52 ℃热激处理组与热激处理后恢复组的热损伤情况(2018)。各参数的计算如下:
Fv/Fm=(Fm-F0)/Fm,
式中,Fv/Fm为PSⅡ的最大量子产量,反映了植物的潜在最大光合能力。
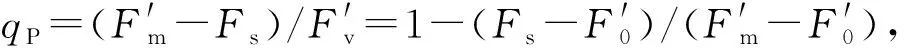

应用hand-PEA进一步检测叶片PSⅡ的荧光参数,各参数计算公式参考李鹏民等(2005)。ABS/RC是单位反应中心吸收的光能; DI0/RC为单位反应中心耗散掉的能量(在t=0时); TR0/RC为单位反应中心捕获的用于还原QA的能量(在t=0时); ET0/RC为单位反应中心捕获的用于还原QA的能量(在t=0时); RE0/RC用于热耗散的量子比率(在t=0时); phi(P0)=Fv/Fm,代表初级光化学反应的最大量子产量; psi(E0)=ψ0≡ET0/TR0,是捕获的激子将电子传递到电子传递链中超过QA的其他电子受体的概率(在t=0时); phi(E0)=ET0/ABS,是用于电子传递的量子产额(在t=0时); delta(R0)反应中心吸收的能量用于热耗散的比率(在t=0时); phi(R0)=RE0/ABS,代表天线色素(Chl)吸收能量(ABC)的光化学反应的能力; ABS/CS0≈Fo,单位面积吸收的光能(在t=0时); DI0/CS0为单位面积的热耗散(在t=0时); TR0/CS0为单位面积捕获的光能(在t=0时); ET0/CS0为单位面积电子传递的量子产额(在t=0时); RE0/CS0代表荧光淬灭通量,它是被激发的单位横截面积上荧光淬灭的通量(在t=0时); ABS/CSm≈Fm,单位面积吸收的光能(在t=m时); DI0/CSm为单位面积的热耗散(在t=m时); TR0/CSm为单位面积捕获的光能(在t=m时); ET0/CSm为单位面积电子传递的量子产额(在t=m时); RE0/CSm代表荧光淬灭通量,它是被激发的单位横截面积上荧光淬灭的通量(在t=m时); PIABS是在吸收基础上的执行指数。

图1 不同温度梯度下Y(Ⅱ)、Y(NO) 和 Y(NPQ)的荧光影像Fig.1 Fluorescence image of Y(Ⅱ), Y(NO) and Y(NPQ) by image-PAM under different temperature gradients
1.6 数据分析
应用image-PAM检测来筛选临界温度的参数Y(Ⅱ)、Y(NPQ)和Y(NO)的荧光动力学曲线和临界温度下参数NPQ、qL和qP的荧光动力学曲线,应用R语言软件包中的ggplot2绘制。用处理组/对照组将各参数进行数据标准化,再将标准化后的数据应用EXCEL绘制雷达图。OJIP曲线也是标准化后应用EXCEL进行绘制的,标准化公式为Vt=(Ft-F0)/(Fm-F0)。
2 结果与分析
2.1 热激临界温度的选择

图2 不同温度梯度Y(Ⅱ)、Y(NO) 和 Y(NPQ)的荧光动力学曲线Fig.2 Kinetics curves of Y(Ⅱ), Y(NO) and Y(NPQ) under different temperature gradients
Y(NPQ)是光保护的重要指标,若Y(NPQ)较高,一方面表明植物接受的光强过剩,另一方面则说明植物仍可以通过调节(如将过剩光能耗散为热)来保护自身; Y(NO)是光损伤的重要指标,若Y(NO)较高,则表明光化学能量转换和保护性的调节机制(如热耗散)不足以将植物吸收的光能完全消耗掉,入射光强超过了植物能接受的程度,这时植物可能已经受到损伤,或者(尽管还未受到损伤)继续照光的话植物将要受到损伤; Y(Ⅱ)代表PSⅡ的光化学能力。应用image-PAM对10个温度梯度热激胁迫下测定的参数Y(NPQ)、Y(NO)和Y(Ⅱ)的荧光影像和曲线进行对比研究,见图1、图2。最终选择了初始胁迫和极限胁迫的2个阈值。当欧洲云杉幼苗在44 ℃热激后与对照组比: 它的荧光影像开始发生变化,Y(Ⅱ)和Y(NO)的影像开始变淡; 并且它的PSⅡ的光化学能Y(Ⅱ)的荧光动力学曲线开始下降,热耗散Y(NPQ)和荧光淬灭Y(NO)的影像动力学曲线开始上升; 因此,选择44 ℃作为热激胁迫的初始胁迫温度。当幼苗在53 ℃热激下时,叶片开始脱落,组织器官开始摧毁,形成不可逆的损伤; 并且光化学能Y(Ⅱ)的动力学曲线开始趋向0,Y(NPQ)曲线远远低于对照,而Y(NO)曲线远远高于对照,说明叶片已经受到严重的损伤。 因此,选择52 ℃热激作为热激胁迫的极限温度。
2.2 应用image-PAM测定临界温度下PSⅡ的荧光参数
由图3、图4可知,荧光影像显示在44 ℃热激胁迫下,Fm、Fv/Fm的影像被减弱,而qL和qP的荧光影像被增强,NPQ的影像没有变化。而在52 ℃热激胁迫下,Fm和NPQ被增强,而Fv/Fm、qL和qP影像被减弱,通过分析NPQ、qL和qP荧光动力学曲线发现,在44 ℃热激下3个参数的曲线都高于对照组,而在52 ℃热激下,这3个参数的曲线低于对照组。以上结果说明,44 ℃热激下PSⅡ的电子吸收、捕获与传导、以及非光化学荧光淬灭的执行效率在一定程度上有促进作用,而在52 ℃热激下,它们受到严重抑制作用。

图3 临界温度下F0、Fm、Fv/Fm、NPQ、qL和qP的荧光影像Fig. 3 Determinating fluorescence image of F0,Fm,Fv/Fm,NPQ,qL and qP under the critical temperature of heat shock stress

图4 临界温度下NPQ、qL和qP的荧光动力学曲线Fig.4 Determinating fluorescence curves of NPQ, qL and qP by under the critical temperature of heat shock stress
2.3 应用hand-PEA测定的临界温度下的PSⅡ的荧光参数
2.3.1 叶绿素(Chl)对光能的吸收和捕获 由图5和表1可知,在44 ℃热激下,雷达图显示荧光参数ABS/RC、TR0/RC、ABS/CS0、(F0)、ABS/CSm(Fm)、TR0/CS0和TR0/CSm都偏离对照不是很明显(图5),它们分别是对照的1.056、1.03、0.946、1.056、1.031和0.925倍。而在52 ℃热激下,都显著偏离对照,其中ABS/RC、TR0/RC、ABS/CS0和TR0/CS0上升为对照的2.782、1.562、2.142和1.248倍,而ABS/CSm和TR0/CSm下降为对照组的0.745和0.484倍。以上结果说明,在44 ℃热激下,叶绿素吸收和捕获光能的能力并没有受到显著影响,但在52 ℃热激下,F0时刻的反应中心吸收的光能和捕获的光能增强,而Fm时刻叶绿素吸收和捕获的光能受到明显的抑制。
2.3.2 光化学反应 在44 ℃热激胁迫下phi(P0)参数曲线与对照组偏离不明显,为对照组的97.7%; 而phi(R0)参数曲线明显偏离对照,phi(R0)升为对照组的121.8%。在52 ℃热激胁迫下,2个参数曲线都明显偏离对照,phi(P0)降为对照组的63.2%,而phi(R0)降为对照组的71.2%。以上结果说明,在44 ℃热激胁迫下对初级光化学反应的最大量子产量没有变化,而使天线色素(Chl)吸收能量(ABC)的光化学反应的能力增强了; 在52 ℃热激下二者都被抑制。
2.3.3 电子传递链效率 在44 ℃ 热激下,ET0/RC、ET0/CS0、ET0/CSm、psi(E0) 和phi(E0)参数曲线偏离对照不明显,量值分别是对照组的1.009、1.009、0.904、0.978和0.955倍。在52 ℃热激下,ET0/CS0、ET0/CSm, psi(E0) 和 phi(E0) 参数曲线明显偏离对照,量值分别降为对照组的0.401、0.179、0.486和0.336倍。以上结果说明,44 ℃ 热激下对PSⅡ的电子传递没有影响,但52 ℃热激胁迫明显抑制了PSⅡ的电子传递通量。
2.3.4 荧光淬灭的能力 在44 ℃热激下,RE0/CS0和RE0/CSm参数曲线开始偏离对照,升为对照的1.284和1.158倍(见表1); 在52 ℃热激下,两参数曲线明显偏离对照,分别为对照的1.483和0.499倍。以上结果说明,在44 ℃热激下,PSⅡ调节过剩光能量的机制,可以通过荧光淬灭的形式把过剩的热量散失掉; 但52 ℃热激下F0时刻可以通过荧光淬灭的形式将过剩的热量散失掉,而Fm时刻通过荧光淬灭调节热耗散的机制被抑制。
2.3.5 热耗散的能力 在44 ℃热激下,DI0/RC、DI0/CS0、DI0/CSm和delta(R0)参数曲线开始偏离对照,量值分别为对照的1.193、1.189、1.056和1.218倍,这说明一定程度上促进热耗散。但52 ℃热激下,DI0/RC、DI0/CS0、DI0/CSm和delta(R0)参数曲线明显偏离对照,量值分别为对照的9.296、6.927、2.142和17.806倍,说明大部分能量以热耗散的能力散失掉,并且热耗散的量子比率上升至对照的17.806倍。
2.3.6 PSⅡ光合作用执行效率 在44 ℃热激胁迫下,OJIP曲线与对照组基本一致,没发生明显偏离; PIABS曲线开始偏离对照,为对照组的79.5%。在52 ℃热激胁迫下,OJIP曲线明显偏离了对照组,出现“K”点,表明PSⅡ中的放氧复合体(OCE)遭到破坏; 而PIABS曲线明显偏离对照,仅为对照组的6.6%。以上结果说明,44 ℃热激胁迫下,PSⅡ光合作用的执行效率没受到大的影响; 而在52 ℃热激胁迫下,光能的吸收、捕获和传递都明显发生抑制。
2.4 临界温度胁迫后恢复的PSⅡ的荧光参数
由图3、图6、图7和表1可知,恢复组检测的PSⅡ荧光参数和影像,又几乎回到对照水平,显示从44~52 ℃度之间这8 ℃温差的过渡都没有引起欧洲云杉PSⅡ光能捕获、吸收和电子传递效率受到不可恢复的损伤,说明欧洲云杉对温度有较高的弹性适应机制。

图5 临界温度胁迫下标准化后PSⅡ荧光参数Fig.5 Radar chart of standardizing fluorescence parameter under the critical temperature of heat shock stress
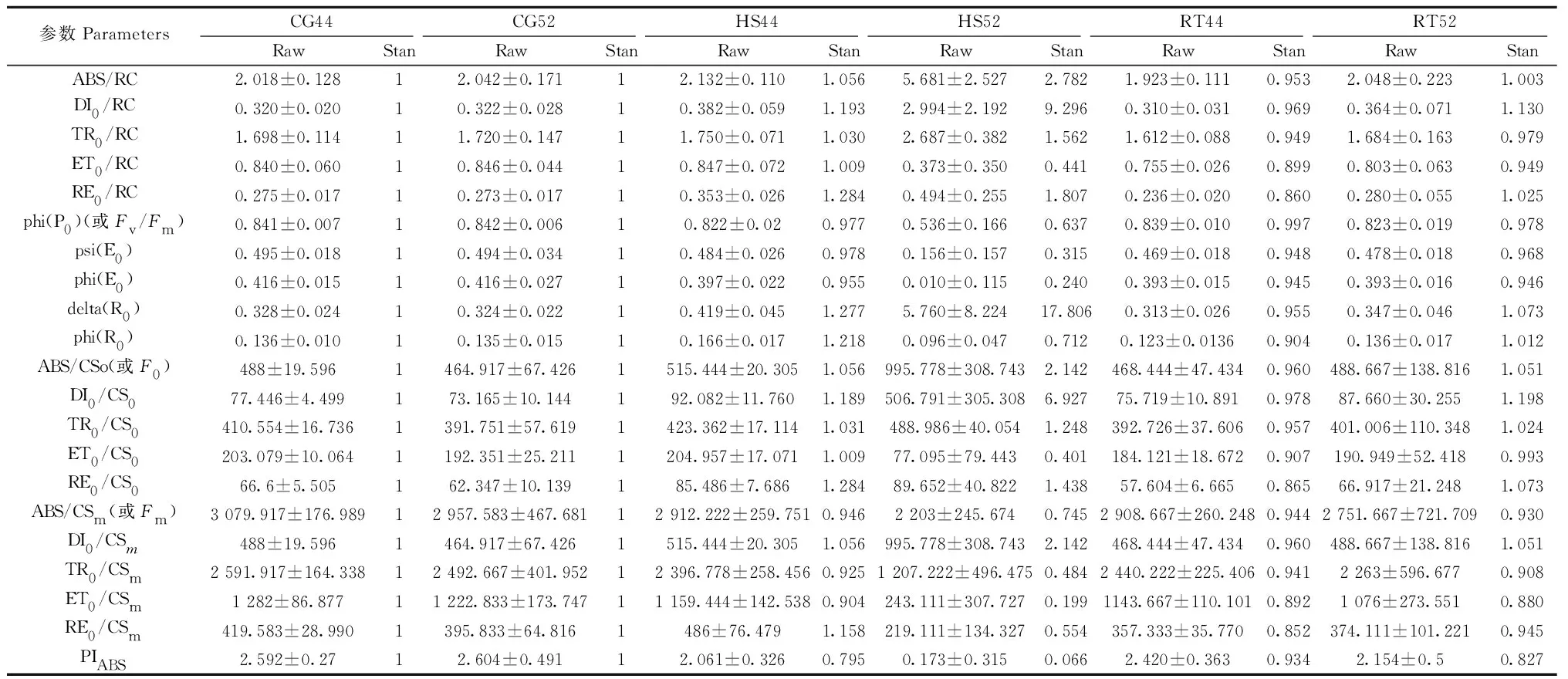
表1 临界温度胁迫下PSⅡ荧光参数和标准化后数据①Tab.1 Fluorescence parameter and standardizing result under the critical temperature of heat shock stress

图6 临界温度胁迫下标准化后的OJIP曲线Fig.6 Standardizing OJIP curvesunder the critical temperature of heat shock stress

图7 应用hand-PEA测定的临界温度胁迫下二周自行恢复后标准化PSⅡ荧光参数Fig.7 Radar chart of standardizing fluorescence parameter by hand-PEA determining under the critical temperature of heat shock stress after self-recovery for two weeks
3 讨论
3.1 欧洲云杉PSⅡ抗热能力评估
执行效率指数(PIABS)和PSⅡ的最大量子产量φPo=Fv/Fm是检测各种胁迫比较敏感的参数,它们被广泛用作对比研究光合作用系统的整体光合效率。对小麦(Tritiumaestivum)的研究,表明在活体植物的叶绿素荧光测定中Fv/Fm和光化学淬灭系数(qP)在高于37.5 ℃时就开始下降(Lu, 2000);而在40 ℃热激下,PIABS降至14%(Mathuretal., 2011)。在巴旦木(Prunuspersica)叶片研究中,观察到了PIABS值在30和35 ℃热激下升高,在40 ℃热激下降低,在45 ℃热激下最小。此外,当叶片暴露在40和45 ℃时,Fv/Fm显著降低,“K”和“L”带大幅度呈现(Martinazzoetal., 2012 )。在对栓皮栎(Q.variabilis)的研究中,热激初始胁迫温度T50是42 ℃, 当温度达到55 ℃时,任一光照状态下的PSⅡ实际量子产量Y(Ⅱ)趋向于0,因此它的胁迫温度是42~54 ℃(陈景玲等,2013)。而仁用杏(Armeniacavulgarisvs.sibirica)30 ℃为初始胁迫温度,PIABS降为对照组的64%,Fv/Fm和OJIP曲线维持不变; 50 ℃为极限胁迫温度,PIABS降为对照组的10.3%,Fv/Fm为对照的37.8%,OJIP曲线在300 us处K点急剧上升(杜国栋等,2011)。在对欧洲冷杉的研究中,发现不同种源初始和极限胁迫温度都不相同,初始胁迫温度范围是42.7~43.2 ℃,Fv/Fm降至50%的极限胁迫温度是51 ℃,种源地温度越高,临界温度的值就越高(Konpkováetal., 2018)。对国内9种云杉属植物PSⅡ热稳定性研究中发现,半致死温度FT50在45~50 ℃之间,长叶云杉(Piceasmithiana)的半致死温度48.35 ℃为9种云杉中的最高值,具有最强的PS Ⅱ热稳定性;红皮云杉、青海云杉和紫果云杉的半致死温度均在46~47 ℃之间相对较低,PSⅡ热稳定性较差;其余云杉属植物类群的半致死温度都在47~48 ℃,PSⅡ热稳定性处于中间水平(杨毅,2017)。总体来说针叶树种Fv/Fm降到50%的温度,处于45~53 ℃之间(Krauseetal., 2010; Mariasetal., 2017),也有研究报道欧洲冷杉的Fv/Fm在40 ℃时急剧降低(Robakowskietal., 2002.)。在本研究发现44 ℃短时热激下image-PAM检测的PSⅡ光化学能Y(Ⅱ),热耗散Y(NPQ)和荧光淬灭Y(NO)的荧光参数的影像和动力学曲线都开始发生改变;但应用hand-PEA检测发现Fv/Fm、OJIP曲线都没有明显变化,但PIABS降至为对照组的79.2%,说明胁迫刚刚开始;而52 ℃短时热激下,PSⅡ的光化学能Y(Ⅱ),热耗散Y(NPQ)和荧光淬灭Y(NO)已发生明显的抑制,OJIP曲线也出现大幅度的“K”带,Fv/Fm降为对照组的62.4%,PIABS仅为对照组的6.6%;在53 ℃短时热激下,Y(Ⅱ)趋向于0,叶片脱落,组织器官开始被破坏;因此本研究未发现Fv/Fm降为对照组的50%的FT50的温度点,52 ℃是欧洲云杉幼苗短时热激的极限温度;将44 ℃和52 ℃短时热激的幼苗放入人工气候室中正常条件下恢复2周后,各荧光参数的及其影像又几乎都恢复到正常水平,说明欧洲云杉的PSⅡ在44~52 ℃之间的短时热激下的损伤可以自行修复。在杨树(Populussimonii)高温胁迫的研究也发现,在42 ℃下热激胁迫短于6 h 时,PSⅡ经过自我修复可恢复正常功能;但温度超过6 h就会造成不可逆转的损伤(Songetal., 2014)。与以往研究对比发现,欧洲云杉的抗高温能力远远高于一些草本植物,也高于一些木本被子植物,如栓皮栎(Quercusvariabilis)和仁用杏,在针叶树种(欧洲冷杉和花旗松)中它的高温耐力处于较高水平,也高于国内的已研究的9种云杉(杨毅,2017),这与它的原生境条件较温暖湿润相关。
3.2 欧洲云杉PSⅡ临界温度下损伤分析
短期高温处理造成的云杉属植物叶片PSⅡ的损伤,可通过已测定的叶绿素荧光参数反映(杨毅,2017)。本研究通过叶绿素荧光参数,对短时不同程度的热激下PSⅡ的量子产量、光能的吸收和捕获、电子传递以及荧光淬灭和热耗散进行研究,揭示了高温胁迫下欧洲云杉PSⅡ的损伤与自我修复能力。
高温胁迫下PSⅡ的量子产量的估测,常用PSⅡ的最大量子产量phi(P0)(或Fv/Fm)和还原PSI受体侧末端电子受体的量子产额phi(R0)来表述。在本研究中,44 ℃热激胁迫下phi(P0)(或Fv/Fm)降为对照的97.7%,而phi(R0)升高为对照的1.218;在52 ℃热激胁迫下phi(P0)(或Fv/Fm)降为对照的63.7%,而phi(R0)降为对照的71.2%;说明44 ℃胁迫对PSⅡ侧光化学效率的抑制刚开始,但促进了PSⅠ的光化学效率,52 ℃胁迫对光系统PSⅠ和PSⅡ侧的光化学效率都产生抑制。在对我国9种云杉属植物的短时热激研究中发现,45 ℃瞬时高温胁迫对PSⅡ光化学效率就产生抑制作用(杨毅,2017),说明欧洲云杉与其他云杉属相比较有较强的抗高温能力。
高温胁迫下,参数ABS/CS0(或F0)和ABS/CSm(或Fm)也是反应植物热损伤程度的重要指标。一般来说,由于受到高温胁迫,植物类囊体膜结构会出现变化导致基础荧光的缓慢增加,而使最大荧光降低,因此基础荧光值的增加值与最大荧光的降低与植物PSⅡ的反应中心受到的损伤程度呈现相关关系。本研究中,44 ℃热激胁迫下ABS/CSm(或Fm)降为对照的94.6%,而ABS/CS0(或F0)升高为对照的105.6倍,也说明胁迫刚开始;在52 ℃热激胁迫下ABS/CSm(或Fm) 降为对照的74.5%,ABS/CS0(或F0)升高为对照的214.2%。我国9种云杉属植物45 ℃瞬时高温胁迫基础荧光值都开始增加,到50 ℃高温胁迫下基础荧光也增加2倍以上(杨毅,2017)。仁用杏的研究中也发现,50 ℃高温导致初始荧光(Fo) 也显著升高,为对照的2.26 倍(杜国栋等,2011)。本研究中光能的吸收和捕获参数44 ℃热激胁迫无明显变化,52 ℃热激胁迫下初始光能的吸收和捕获参数ABS/RC、TR0/RC、ABS/CS0和TR0/CS0上升为对照的2.782、1.562、2.142和1.248倍,而t=m时的ABS/CSm和TR0/CSm下降为对照组的0.745和0.484倍,说明52 ℃热激下PSⅡ对能量的捕获和吸收的能力增强,而天线色素捕获和吸收能量的能力减弱。已研究的我国9种云杉属植物,ABS/RC和TR0/RC在20~40 ℃热激时变化也不明显,而在到50 ℃时反应中心有活性的数量越来越少,使ABS/RC和TR0/RC显著升高,其中ABS/RC升高为对照的40倍以上,ABS/CSm和TR0/CSm显著下降(杨毅,2017)。与其对比,说明欧洲云杉PSⅡ供体侧的抗高温能力显著高于国内其他云杉属。在高温胁迫下PSⅡ的电子传递的能力也受到了不同程度的损伤阻断。本研究44 ℃热激下,电子传递效率变化不明显。在52 ℃热激下,ET0/RC、ET0/CS0、ET0/CSm、psi(E0) 和 phi(E0)分别降为对照组的0.441、0.401、0.179、0.486和0.336倍。在对我国9种云杉属植物,20~45 ℃短时热激下ET0/RC和ET0/CS下降;在50 ℃热激下都明显下降,ET0/RC降为对照的0.47倍左右(杨毅,2017)。与其对比,说明欧洲云杉PSⅡ受体侧的抗高温能力处于中等水平。
荧光淬灭参数,在44 ℃热激下,RE0/CS0和RE0/CSm升至对照的1.284和1.158倍;在52 ℃热激下,RE0/CS0和RE0/CSm分别为对照的1.483和0.499倍。热耗散参数在44 ℃热激下DI0/RC、DI0/CS0、DI0/CSm和delta(R0)分别为对照的1.193、1.189、1.056和1.218倍;但在52 ℃热激下,DI0/RC、DI0/CS0、DI0/CSm和delta(R0)分别为对照的9.296、6.927、2.142和17.806倍。我国9种云杉属植物20~45 ℃短时热激下,DI0/RC和DI0/CS略有增加,而在50 ℃热激下是正常状态下的40倍左右(杨毅,2017)。说明欧洲云杉PSⅡ在热激状态下自主调节热量散失的能力高于国内其他云杉属树种。
4 结论
1) 找到了欧洲云杉幼苗短时热激胁迫下的初始胁迫温度44 ℃和极限胁迫温度52 ℃,并预测欧洲云杉在未来气候变暖的情况下有较强的抗热能力,在抗热性上处于同属物种及其近缘物种的较高水平。2) 预测44 ℃短时热激下,PSⅡ的光化学反应、热耗散和荧光淬灭的能力都被促进;但52 ℃短时热激下,PSⅡ的光化学反应、热耗散和荧光淬灭的能力都被明显抑制。3) 44 ℃初始胁迫温度虽使PIABS略有降低,但叶绿素(Chl)对光能的吸收和捕获、电子的传递、电子传递链中心蛋白的活性都没受到明显的抑制,52 ℃极限胁迫温度下叶绿素(Chl)对光能的吸收和捕获、光化学反应、荧光淬灭和电子传递的能力遭到严重的抑制,并且电子传递链的放氧复合体(OEC)也被破坏。说明在未来气候变暖的情况下欧洲云杉在午间瞬时高温的影响下,会主要抑制叶片PSⅡ反应中心的活性和受体侧电子传递,改变PSⅡ反应中心能量流分配,而对PSⅡ供体侧的影响较小。4) 在恢复组中,2种胁迫温度下幼苗针叶的荧光参数和影像都恢复接近对照水平,说明欧洲云杉对短期的胁迫有较强的自我修复能力。因此可预测,在未来气候变化的情况下欧洲云杉有较强的弹性适应生理机制,来应对极端天气午间的短时高温,它在短时热胁迫下可自我恢复的温度范围在44~52 ℃。
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
数学物理学报(2020年6期)2021-01-14
现代农村科技(2020年7期)2020-12-19
花卉(2020年6期)2020-01-07
计算机时代(2019年10期)2019-11-11
乡村科技(2019年14期)2019-02-21
基层中医药(2018年8期)2018-11-10
启迪与智慧·教育版(2018年12期)2018-02-26
