
川西亚高山暗针叶林及其采伐次生林林下分层谱系结构*
2020-08-14 02:30陈欢欢许格希马凡强张淼淼曹向文赵广东杨洪国史作民
林业科学 2020年7期
陈欢欢 许格希 马凡强 刘 顺 张淼淼 曹向文 陈 健 赵广东 杨洪国 史作民,4
(1. 中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091; 2. 四川米亚罗森林生态系统定位观测研究站 阿坝 623100; 3. 中国林业科学研究院湿地研究所 北京 100091; 4. 南京林业大学 南方现代林业协同创新中心 南京 210037)
研究森林群落物种组成及其谱系结构,有助于揭示群落构建的驱动机制,是生物多样性保护研究的重点关注内容之一(Kressetal., 2009; Fengetal., 2012; 马克平, 2016)。在群落的谱系结构及其生态学意义的研究中,早期多关注结构相对简单的草原生态系统(Fargioneetal., 2003; Silvertownetal., 2006),近十年在陆地生物多样性最丰富的热带亚热带森林生态系统中相关领域研究得到充分发展(Smithetal., 2011; Swensonetal., 2013; Vamosietal., 2014; Döbertetal., 2017)。生物信息学与群落生态学的结合衍生了谱系群落生态学这一生态学分支学科(Qianetal., 2014)。群落谱系结构的量化指标为揭示群落构建机制提供可量化的评判标准,经过近20年的发展,已开发出数十个可用于衡量群落谱系结构的指数(Swenson, 2014),其中以净亲缘指数(NRI)和最近类群指数(NTI)应用最广泛(Fengetal., 2015; Khaliletal., 2016)。
近些年,谱系群落生态学的发展先后验证了森林群落构建的一系列理论和假说(Pringleetal., 2016; Wangetal., 2016; Welshetal., 2016; 马克平, 2016),其中以森林固定长期监测样地(简称“大样地”)的研究最为广泛。例如,在美国巴洛克罗拉多岛(Barro Colorado Island)(Chase, 2014)和我国的西双版纳热带雨林(Yangetal., 2013)大样地,科研工作者验证基于生态位理论的环境过滤(Murphyetal., 2014)和竞争互斥(Hartetal., 2016)以及基于中性理论的扩散限制性(Huetal., 2012)在森林群落结构和功能中发挥的作用(Lohbecketal., 2016)。但是,森林大样地植被多为保护较好的原始林,相比演替次生林具有较高的均质性,这就导致验证人为干扰后次生森林演替过程中驱动群落构建的生态学过程变得相对困难。从演替角度研究森林群落物种组成谱系结构,有助于更深入了解生境特异性在群落物种演化中的作用(Letcheretal., 2015)。
川西亚高山林区地处青藏高原东南缘与长江上游,历史上形成了以暗针叶林为特色的典型植被,森林动植物资源极其丰富,在生物多样性保育上发挥着“屏障”作用(蒋有绪, 1963; 1981)。川西米亚罗林区海拔落差大,植被呈明显垂直带特征: 亚高山针阔混交林主要分布于海拔2 700 m以下,亚高山针叶林及高山疏林带主要分布于海拔2 700~4 000 m,垂直往上则是稀疏灌丛、杜鹃(Rhododendronspp.)灌丛和高山草甸等的分布区(蒋有绪, 1963)。但是, 1950—1989年我国对该区域进行了以皆伐为主的大规模的森林开发利用(刘彦春等, 2010),伴随而来的是生物多样性丧失等生态问题(Zhouetal., 2008)。1998年天然林保护工程实施后该林区森林得到有效保护,当前采伐森林生态系统正处于全面恢复阶段。针对川西亚高山林区森林生态系统已开展了诸多研究,涵盖生态系统生物量-生产力(刘兴良等, 2006)、乔木优势种幼苗更新(王微等, 2007)、土壤理化性质和生态化学计量特征(邹婷婷等, 2017; 程欢等, 2018)、根系与微生物结构(刘顺等, 2019)等,但对林下(或林内)乔灌草分层物种组成的谱系结构及对其驱动的生态学过程还未见报道。鉴于此,本研究以川西米亚罗林区原始暗针叶林及其采伐次生林为对象,分析不同演替阶段森林群落林下层次的谱系结构及其物种共存的生态学驱动过程,为生物多样性保护、生态系统修复等提供理论依据。
1 研究区概况
研究区位于四川省理县米亚罗省级自然保护区303林场(102°53′―102°57′ E,31°14′―31°19′ N),海拔2 458~4 619 m,受青藏高原东缘高原峡谷影响,气候夏凉冬寒,属丹巴-松潘半湿润气候,年均气温2~4 ℃(最高气温23.7 ℃,最低气温-18.1 ℃),年降水量700~900 mm,集中在5—10月(唐仕姗等, 2015)。植被垂直带明显,原始森林以岷江冷杉(Abiesfaxoniana)和紫果云杉(Piceapurpurea)为优势种形成的亚高山暗针叶林为主,当前主要分布于海拔3 500~4 000 m。天然次生林主要分布于海拔2 400~3 800 m的暗针叶林采伐迹地,经过以悬钩子属(Rubus)和箭竹属(Fargesia)植物为主的演替初期,演替到当前以桦木属(Betula)、槭属(Acer)和椴树属(Tilia)为主,伴生有云杉属(Picea)和冷杉属(Abies)树种,森林土壤为山地棕壤(刘彦春等, 2010)。
2 研究方法
2.1 样地设置与调查
2017年6—10月,根据川西亚高山暗针叶林天然分布现状与历史采伐资料,在岷江冷杉原始林(林龄>120年)、1950—1969年和1970—1989年采伐后形成的天然次生林共布设22块20 m×20 m样地(其中原始林6块,另外2个年代次生林各8块)。将每块样地划分为100个2 m × 2 m样方,进行群落生态学调查,包括乔灌幼苗、灌木、乔木幼树、小乔木、高大乔木的个体数量,及其样方内坐标、胸径或基径、高度与冠幅,同时记录样地出现的草本、蕨类和苔藓植物的种数。川西亚高山不同演替阶段森林群落的乔灌草层次结构明显,林下植物物种丰富,但主林层(林冠层)乔木树种相对单一,针对主林层进行谱系结构分析的意义不大,因此该研究重点关注林下乔灌草层次的谱系结构。
根据接受光照程度及植物生长发育特性,参考在川西亚高山林区开展的已有研究(李媛等, 2007; 缪宁等, 2009),将川西亚高山不同演替阶段森林群落的林下层次结构划分为: 1)草本层(高度≤0.3 m) 以草本植物和乔灌幼苗为主; 2)灌木层(0.3 m<高度≤3 m) 以灌木植物为主,还包括一定比例的乔木幼苗、幼树; 3)小乔木层(3 m<高度≤ 5 m) 主要包括大部分的杜鹃属(Rhododendron)植物和乔木植物的幼树,此外还包括部分高大灌木和冷箭竹(Bashaniafangiana)等; 4)亚林层(5 m<高度≤主林层) 包括大部分群落优势种或建群种的未成熟个体,以及受遗传演化特性影响无法到达主林层的阔叶乔木,例如高大杜鹃属植物、西南樱桃(Cerasusduclouxii)和陕甘花楸(Sorbuskoehneana)等。川西亚高山森林亚林层与主林层的分界线主要受林型、林龄、树种组成及环境因子等多因素综合影响,因此本研究结合研究区调查的森林乔木的检尺数据以及蒋有绪等(1998)对该区域森林群落特征的描述,将原始林中胸径DBH ≥ 20 cm的活立木平均高作为该林型主林层与亚林层的分界线,活立木高度大于等于平均高(20.2 m)则视为进入主林层,反之归属于亚林层。类似地,将次生林DBH ≥ 15 cm的活立木平均高(1950―1969年与1970―1989年次生林分别为11.4和12.4 m)定为主林层与亚林层的分界线。
2.2 川西亚高山不同演替阶段森林群落谱系树构建
根据研究区森林样方植物调查数据,将乔灌草等种子植物的物种信息在国际植物清单数据库(http:∥www.theplantlist.org/)中进行检索后生成“科/属/种”模式的物种名录,继而在phylomatic平台(http:∥phylodiversity.net/phylomatic/)输入这些物种信息,根据Zanne等(2014)发表的物种系统进化框架生成具有进化枝长的谱系树。对于谱系树上出现的多分枝(polytomy)情况,利用R语言ape包multi2di命令将这些亲缘相近的多分枝进行随机分解,最终生成具有二分枝结构的谱系树,完成了川西亚高山不同演替阶段森林群落乔灌草植物谱系树的构建(图1)。
2.3 谱系多样性指数和物种多样性指数选择与计算
选择2个最广泛使用的谱系多样性指数,即净亲缘指数(NRI)和最近类群指数(NTI),综合评估不同演替阶段森林群落的林下乔灌草谱系结构的整体及局部特征(Swenson, 2014),选择能综合考虑物种多度与频率的香侬多样性指数H′表征乔灌草物种多样性(Pielou, 1966)。
NRI可用于量化群落谱系树上组成物种整体聚集情况,是群落中任意两个物种平均成对谱系距离相对于随机零模型(null model)生成平均成对谱系距离的标准化指数,计算方法如下:
(1)

NTI可用于量化群落谱系树末端分枝(局部分枝)亲缘相对较近物种的聚集程度,是对NRI的有效补充。NTI是群落中亲缘关系最相近物种平均成对谱系距离(mean nearest taxon distance)相对于随机零模型生成平均最近类群成对谱系距离的标准化指数,计算方法与NRI类似:
(2)

H′计算公式如下:
(3)
式中:N为群落物种数量;pi为物种i个体数量在群落中所占有的比例。群落组成物种越多且均匀度越高,H′值越大; 反之,群落优势种或建群种个体数占有比例越大,H′值越小。
2.4 数据分析
所有数据分析及图形绘制均在R语言中实现。分离不同演替阶段森林群落的林下乔灌草层次的物种组成及丰富度后,利用R语言geiger包中的name.check与drop.tip命令,将各层次物种组成的谱系框架从系统发育树中进行提取。结合各层次物种在各样地的分布情况,利用picante包中的ses.mpd与ses.mntd命令分别计算NRI与NTI。ses.mpd与ses.mntd命令运行过程中进行999次类群(物种)随机分配(taxa shuffle)模型模拟(1 000次迭代),模型模拟中考虑物种组成及其丰富度差异的影响。利用单因素方差分析(one-way ANOVA)和Turkey’s HSD分别检验林下乔灌草谱系结构的层次间与林型间的差异性; 采用线性回归,分析不同演替阶段森林群落的林下乔灌草NRI与NTI的相关性。利用局部多项式回归拟合分析乔灌草谱系结构(NRI和NTI)与H′的关系。
3 结果与分析
3.1 林下乔灌草分层物种谱系组成
川西亚高山原始暗针叶林及其采伐次生林群落植物的林下乔灌草分层物种组成丰富,共调查记录乔灌草植物个体17 812株,隶属44科100属145种。1970―1989年次生林相比其他2个林型具有更多植物物种(36科72属89种),林下乔木层建群种主要有红桦(Betulaalbosinensis)、长尾槭(Acercaudatum)、五尖槭(Acermaximowiczii)、华椴(Tiliachinensis)、西南樱桃和陕甘花楸,灌木层建群种有华西箭竹(Fargesianitida)、鲜黄小檗(Berberisdiaphana)、毛花忍冬(Loniceratrichosantha)和秀丽莓(Rubusamabilis),草本层建群种有车叶葎(Galiumasperuloides)、掌叶橐吾(Ligulariaprzewalskii)和宽叶荨麻(Urticalaetevirens)等。1950―1969年次生林有33科59属77种植物,乔木层建群种有糙皮桦(Betulautilis)、方枝柏(Sabinasaltuaria)、西南樱桃和陕甘花楸,灌木层建群种有喇叭杜鹃(Rhododendrondiscolor)、无柄杜鹃(Rhododendronwatsonii)、细枝茶藨子(Ribestenue)、峨眉蔷薇(Rosaomeiensis)、唐古特忍冬(Loniceratangutica)、毛花忍冬和秀丽莓,草本层建群种有高山露珠草(Circaeaalpina)、白花酢浆草(Oxalisacetosella)和轮叶黄精(Polygonatumverticillatum)等。原始林(林龄>120年)有31科54属72种植物,乔木层建群种有岷江冷杉(Abiesfaxoniana)、云杉(Piceaasperata)、四川红杉(Larixmastersiana)、糙皮桦、西南樱桃和陕甘花楸,灌木层建群种主要有无柄杜鹃、峨眉蔷薇、细枝茶藨子、绣球藤(Clematismontana)、唐古特忍冬和毛花忍冬,草本层主要物种有披针薹草(Carexlancifolia)、高山露珠草、和异型假鹤虱(Eritrichiumdifforme)等。
从组成物种间谱系关系看,川西亚高山原始暗针叶林及其采伐次生林的植物物种系统发育历史悠久,且物种系统发育组成相对离散(植物种属比为1.45)(图1)。杜鹃花科(Ericaceae)植物与猕猴桃科(Actinidiaceae)植物物种亲缘关系较近,菊科(Compositae)与伞形科(Umbelliferae)和五加科(Araliaceae)植物物种亲缘关系较其他物种相近,蔷薇科(Rosaceae)、荨麻科(Urticaceae)与杨柳科(Salicaceae)植物物种亲缘关系相对较近,毛茛科(Ranunculaceae)植物与以上几大科物种亲缘关系相疏。单子叶植物百合科(Liliaceae)、菝葜科(Smilacaceae)与禾本科(Gramineae)植物亲缘关系相近,归为一类; 几个裸子植物针叶树种(岷江冷杉、四川红杉、方枝柏、云杉)聚为一类,与其他植物区分开(图1)。
3.2 林下乔灌草分层谱系结构
川西亚高山不同演替阶段森林群落的林下乔灌草分层谱系结构净亲缘指数NRI与最近类群指数NTI的分布模式相似(图2)。从林下不同层次组成物种的整体谱系结构(NRI)来看,原始林中草本层、灌木层与小乔木层物种组成倾向于谱系聚集(NRI > 0),即这些林下层次组成物种亲缘关系相对更近。而亚林层物种组成则倾向于谱系离散(NRI < 0),表明亲缘相对较远的物种共存于该林层。2个不同年代的次生林分别在草本层和亚林层表现出林型间相似的谱系结构,其中在草本层均表现为谱系离散,而亚林层则均表现为谱系聚集。在灌木和小乔木层物种组成上,1950―1969年次生林均呈谱系聚集模式,而1970―1989年次生林均呈谱系离散模式(图2)。在原始林和1950―1969年次生林的灌木层中少数亲缘非常接近物种的占比极大(NTI > 0),而1970―1989年次生林草本层、灌木层与小乔木层较少由亲缘关系接近的物种组成(NTI < 0)。原始林的亚林层物种相对单一且物种亲缘相疏,表现出明显的离散(NTI < 0),局部分枝几乎不存在亲缘相近的物种; 而次生林在该层次上则多为谱系聚集,(主要由桦木科(Betulaceae)和槭树科(Aceraceae)等先锋树种组成)(图2)。
川西亚高山森林群落演替过程中的林下乔灌草谱系结构净亲缘指数(NRI)与最近类群指数(NTI)极显著正相关(P<0.001 ),且这种相关性在林型间具有一致性(图3)。其中1970―1989年次生林NRI与NTI相关性最为密切(R2=0.761,P<0.001 ),原始林次之(R2=0.741,P<0.001 ),1950―1969年次生林相对较弱(R2=0.688,P<0.001 )。
相同乔灌草层次不同林型间以及同一林型不同林下层次间的谱系指数多重比较结果如下。1)1970―1989年次生林灌木层和小乔木层的NRI与1950―1969年次生林对应层次NRI差异显著(P< 0.05); 原始林亚林层NRI显著(P< 0.05)不同于其他2个年代次生林,但是草本层NRI与NTI均未在林型间表现出显著差异性(P> 0.05)。2)同一林型不同林下层次多重比较发现原始林灌木层NRI与亚林层NRI差异显著(P< 0.05),但与另外2个层次(草本层和小乔木层)无显著差异,此外,亚林层NTI与其他3个林下层次NTI具有显著性差异,但这3个林下层次间无显著差异性; 1950―1969年次生林林下各层次谱系结构均无显著差异; 1970―1989年次生林草本层NRI与灌木层NRI无显著差异,但前者与小乔木层差异显著,后者与亚林层差异显著,亚林层的NTI则与另外3个层次的NTI存在显著差异性(表1)。

图1 川西亚高山不同演替阶段森林群落乔灌草谱系组成及其亲缘关系Fig.1 Phylogenetic assemblages and their relatedness of trees, shrubs and grasses in different successional forest communities on the subalpine of western Sichuan
3.3 林下层次谱系结构与物种多样性间的相关性
Pearson相关性分析发现NTI与H′仅在原始林亚林层中呈显著正相关(相关系数为0.823,P<0.05),而NRI与H′仅在1950―1969年次生林亚林层中表现出显著相关性(相关系数为0.822,P<0.05,表2)。图4表明: 1950―1969年次生林林下谱系结构随物种多样性的增加变化不明显,但原始林与1970―1989年次生林呈明显的波峰-波谷交替变化的现象; 当H′=1.0时原始林NRI与NTI达到最小值(明显的谱系离散),之后总体随H′增大而逐渐上升向谱系聚集转变; 1970―1989年次生林NRI、NTI随H′增大的变化趋势与原始林相反,当H′≥1.0时,NRI与NTI整体开始呈下降趋势,并在H′=2.0时谱系离散程度达到最大(明显的谱系离散)。
4 讨论
4.1 森林群落林下层次谱系结构及其生态学意义
光是影响绿色植物生长发育关键的环境因子(Givnish, 1988),森林群落林下乔灌草垂直分层是植物经过长期的系统演化适应了光梯度等环境异质性的结果(Jiaoetal., 2007)。亚高山森林生态系统具有明显的林下乔灌草分层结构,开展林下分层谱系结构研究有助于系统揭示影响群落构建的生态学过程。不同植物物种由于生活型、生长速度、年龄等的差异,相同径级的物种不一定处于相同的垂直高度,本研究根据林层划分去界定物种共存策略及其影响因子可能更加合理。
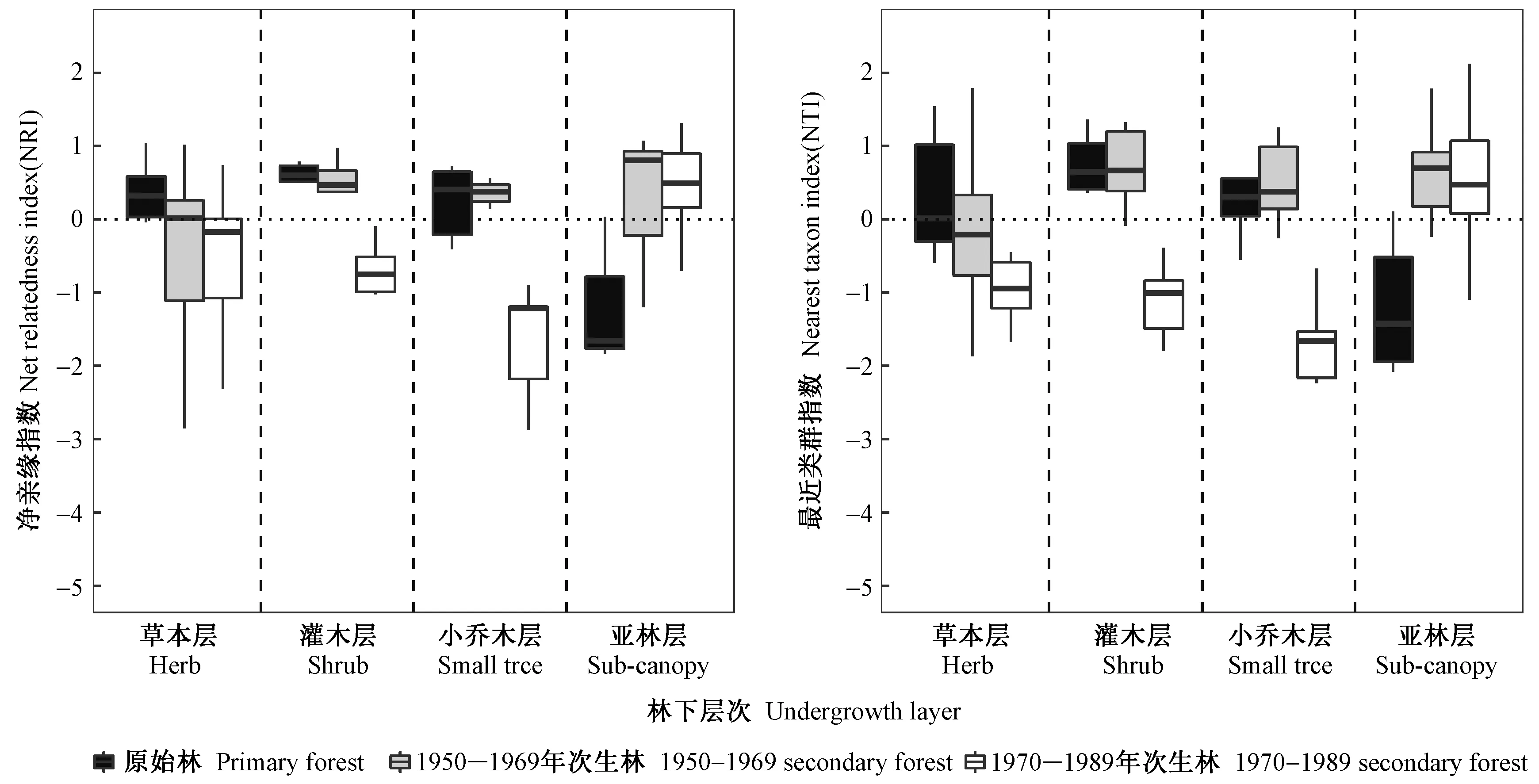
图2 川西亚高山森林群落林下乔灌草分层NRI与NTI的演替模式Fig.2 Successional patterns of NRI and NTI for undergrowth layers in forest communities on the subalpine of western Sichuan
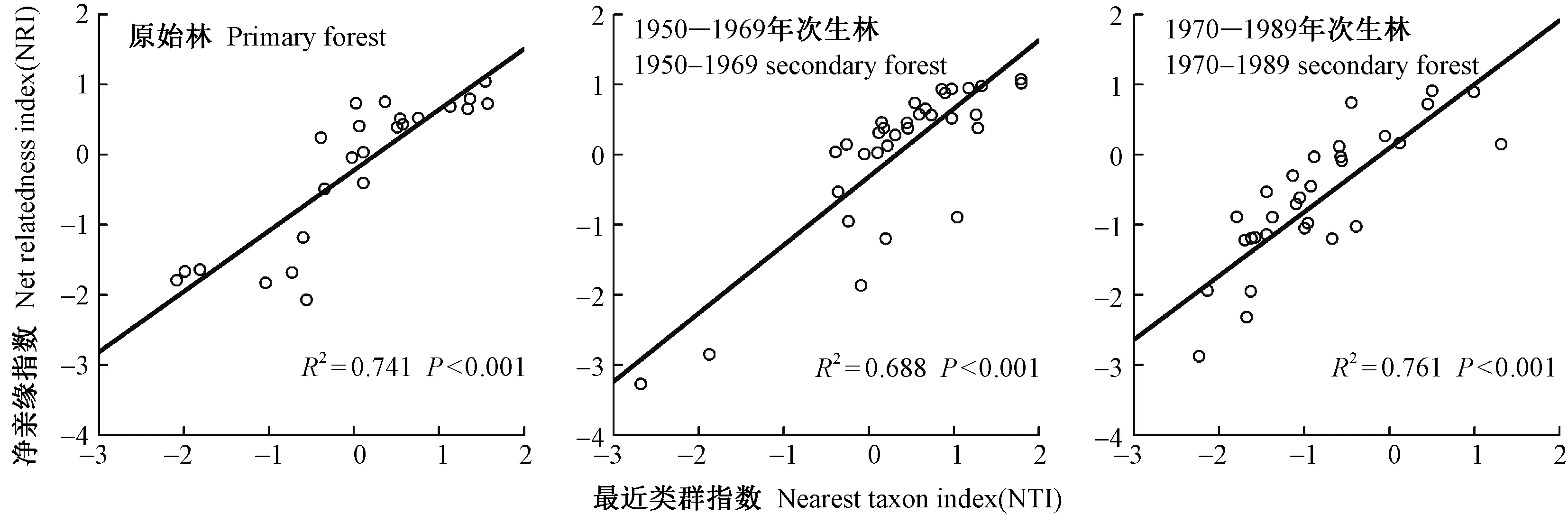
图3 川西亚高山不同演替阶段森林群落林下乔灌草NRI与NTI的相关性Fig.3 Correlations between NRI and NTI of undergrowth tree-shrub-herb layers across different successional forest communities on the subalpine of western Sichuan

表1 川西亚高山不同演替阶段森林群落的林下乔灌草分层NRI与NTI的多重比较①Tab.1 Multiple comparisons of NRI and NTI across undergrowth layers and across successional forest communities on the subalpine area of western Sichuan

表2 川西亚高山不同演替阶段森林群落的林下各层次NRI、NTI与H′的Pearson相关性分析①Tab.2 Pearson correlation analysis of NRI, NTI and H′ for each undergrowth layer of different successional forest communities in the subalpine area of western Sichuan

图4 川西亚高山不同演替阶段森林群落林下乔灌草谱系结构(NRI与NTI)与物种多样性(H′)的局部多项式回归拟合Fig.4 Local polynomial regression fitting between phylogenetic structure(NRI and NTI) and species diversity(H′) of undergrowth tree-shrub-herb layers across successional forest communities on the subalpine of western Sichuan
川西亚高山原始暗针叶林中环境过滤是林下草本层、灌木层和小乔木层物种谱系组成的主要驱动生态学过程,这可能是由于乔木层郁闭度非常高(本研究涉及的岷江冷杉原始林郁闭度为83.6% ± 4.9%),导致光作为最重要的“环境筛”选择那些具备耐荫生态位的物种组成对应的林下层次(Sapijanskasetal., 2014); 而在亚林层上则可能是竞争互斥发挥更大的作用,导致组成物种多亲缘相疏。通常认为物种演化过程中具有谱系生态位保守性(Losos, 2008),即亲缘相近的物种应具有相似的性状(Cooperetal., 2010),因此推断在老龄林中亚林层乔木生长长期受限于主林层,但随着主林层乔木的衰败和死亡(调查中常见衰倒木形成的林窗),亚林层乔木有望取而代之进入主林层(徐宁等, 2013)从而导致亚林层种间竞争异常激烈(Godoyetal., 2014),物种只能通过生态位互补充分利用林下资源(Niklausetal., 2017)。处于演替前期的1970―1989年次生林在草本层、灌木层以及小乔木层物种谱系组成上表现出显著的离散现象,此时种间竞争最为激烈,形成相对于其他2个林型最为丰富的物种多样性。由于1970―1989年次生林分布区的海拔较其他2个森林类型低400~500 m,随着温度增高物种多样性往往增加(Scherreretal., 2011)。
4.2 谱系结构与物种多样性相互独立
谱系结构(谱系多样性)与物种多样性是生物多样性的两个重要维度(马克平, 2016; Pavoineetal., 2016)。川西亚高山不同演替阶段森林群落的林下乔灌草谱系结构量化指数NRI和NTI与物种多样性量化指数H′均不存在显著的线性相关,表明该林区森林群落林下乔灌草物种谱系多样性极有可能与物种多样性相互独立,这与近期在全球尺度的分析结论一致(Pavoineetal., 2016)。局部多项式回归拟合发现原始林与演替前期次生林谱系多样性与物种多样性均存在明显的波峰-波谷现象,对于这是普遍规律还是偶然发现及其背后的生物学意义,还需要更多观测数据的支持与验证。总而言之,本文建议在进行川西亚高山林区植物多样性保护时既要考虑保存尽量多的物种,还应考虑有些物种在系统演化上的关键地位(彭丹晓等, 2017; 蒋有绪等, 1998)。
4.3 问题与展望
本研究调查了川西米亚罗林区不同演替阶段的22块样地(共调查记录个体17 812株),能相对准确地表征川西亚高山不同演替阶段森林群落的林下乔灌草谱系结构及对其驱动的生态学过程。但是,由于本研究构建的谱系树存在属种水平多分枝情况,虽通过亲缘相近随机分解生成二分枝结构,符合统计学上的要求(Zanneetal., 2014),但仍然会对NRI和NTI的计算产生一定的误差。另外,群落谱系结构及其推导的生态学过程是基于群落物种组成谱系生态位保守性假说(Losos, 2008),未考虑物种在群落中的表型可塑性(Andersonetal., 2015),具有一定的局限性(Fritzetal., 2010)。近年来,有学者提出基于植物功能性状进化谱系独立性比较的谱系多样性指数D(Pearseetal., 2013),综合考虑了物种、谱系和功能的信息,全面准确地揭示出了物种生存策略、共存机制及驱动的生态学过程。综上所述,本研究认为有必要尽快建立西南高山亚高山森林区域水平高分辨率(种水平)生命树(鲁丽敏等, 2014),促进和完善西南高山亚高山地区植被物种系统演化的生物地理格局研究(沈泽昊等, 2017; 王家坚等, 2017)。
川西亚高山天然次生林当前处于全面恢复阶段,同时以云杉属和冷杉属树种为主的原始暗针叶林处于过熟林阶段,林下乔灌草分层谱系结构生态学研究能为生态系统修复或林分调整带来一些启发(Zhouetal., 2008)。例如,天然次生林草本层谱系离散,表明物种竞争互斥作用较强,对资源的竞争会限制乔木幼苗的萌发和生长,可考虑采取适当的除草抚育措施促进幼苗更新(马姜明等, 2009); 而亚林层主要由桦木科和少数槭树科树种(先锋树种)占主导,物种负密度依赖效应作用显著,建议减少干预,发挥亚林层促进森林正向演替的作用(缪宁等, 2009)。相反,原始暗针叶林亚林层组成物种种间竞争激烈,为避免有限环境与养分资源的浪费,可根据亚林层树种的分布格局进行林分调整(例如开辟林窗),促进目的树种尽快成材且出好材。
5 结论
川西亚高山林区不同演替阶段森林群落的林下乔灌草谱系结构在层次间与林型间均表现出一定的差异性。川西亚高山不同演替阶段森林群落的林下乔灌草谱系结构量化指数NRI与NTI显著相关,但它们与H′几乎不相关,支持谱系多样性与物种多样性是生物多样性2个独立维度的假说。环境过滤(主要是光梯度)是驱动川西亚高山原始暗针叶林中草本层、灌木层和小乔木层物种谱系组成的主要生态学过程,竞争互斥则塑造了亚林层的物种组成。1950―1969年次生林草本层物种组成主要受竞争互斥影响,其余3个层次主要受环境过滤影响; 1970―1989年次生林亚林层主要受环境过滤影响,其余3个层次物种组成受制于竞争互斥。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
作物学报(2022年2期)2022-11-06
红领巾·成长(2022年6期)2022-07-23
金桥(2020年9期)2020-10-27
天然产物研究与开发(2018年10期)2018-11-06
现代农业研究(2017年11期)2018-01-12
海峡姐妹(2017年10期)2017-12-19
广东农业科学(2017年5期)2017-08-29
现代园艺(2017年20期)2017-02-03
农业与技术(2016年6期)2016-05-14
