
磷硒配施对郑麦9023硒吸收及土壤硒形态转化的影响
2020-08-01 01:21祝姣姣聂兆君李金峰秦世玉刘红恩李永革
麦类作物学报 2020年6期
祝姣姣,聂兆君,赵 鹏,李金峰,秦世玉,李 畅,刘红恩,李永革
(1.河南农业大学资源与环境学院,河南郑州 450002; 2.安阳市农业环境监测站,河南安阳 455000)
硒(Se)是天然存在的非金属元素,是人类、动物以及微生物必需的微量元素,它参与人体中的抗氧化防御系统和调节激素平衡等过程[1]。此外,硒也是一些植物的有益元素,适量的硒不仅能够促进植物的生长发育、提高产量品质,而且也在提高环境胁迫抗性、拮抗重金属、调节光合作用等方面发挥着重要的作用[2]。资料显示,全球估计多达10亿人的日常饮食可能缺乏足够的硒来维持他们的健康。在一些国家,如中国、埃及和泰国,土壤中固有的硒浓度很低,导致人类饮食中缺硒[3-4]。硒缺乏会造成人体甲状腺机能减退、免疫系统减弱、认知能力下降、心血管疾病和各种癌症发病率增加[5]。FAO、WHO、IAEA三个国际组织认可并采用的人体硒含量标准最低需要量为17 μg·d-1,生理需要量为40 μg·d-1,而我国成人硒摄入量不足26.63 μg·d-1,因此人体缺硒现象相当严重[6]。以含硒化学物质作为人类和动物的直接硒源,其风险较大,通过食物链的转化获得适量的硒营养,是一种安全、可行的办法[7]。小麦作为我国北方主要粮食作物,提高其籽粒硒含量对于该地区人体合理补充硒营养尤为重要。硒对植物的生物有效性及植物对硒的吸收累积量不仅取决于土壤中硒的总量,与硒的赋存形态也密切相关[8]。根据硒与土壤结合的成分不同,一般将土壤中硒的赋存形态划分为5种,即水溶态、可交换态、铁锰氧化物结合态、有机结合态和残渣态。水溶态和可交换态硒易被植物吸收利用被称为有效态硒。磷是作物生长必需的大量元素之一,与硒虽为不同族元素,但他们在土壤中均以阴离子形态被作物吸收,两者的离子半径和化学形态相似。Cater等[9]在美国发现,在14种土壤中添加磷,增加了硒在紫花苜蓿中的吸收和积累。然而,也有报道施磷能显著降低植物对硒的吸收[10]。研究表明,磷与硒在植物吸收方面既存在拮抗作用,也存在协同作用。当土壤中磷酸根含量较少时,磷与硒之间主要表现为拮抗作用,反之则表现为协同作用[11]。本研究通过盆栽试验,分析了磷硒配施对冬小麦硒吸收累积和土壤硒形态的影响,以期为深入了解磷硒互作规律以及提高土壤硒的有效性提供参考依据。
1 材料与方法
1.1 供试材料
供试土壤为壤质潮土,取自河南农业大学许昌校区试验场,其基本理化性质见表1。供试小麦品种为郑麦9023。
1.2 试验设计
将过20目筛的风干土8.5 kg装入直径 310 mm、深度 200 mm 的聚乙烯塑料盆中。设3个磷水平(0、80、160 mg·kg-1)和 2个硒水平(0、1 mg·kg-1),共6个处理(Se0P0、Se0P80、Se0P160、Se1P0、Se1P80、Se1P160),每个处理重复4次。分别以 KH2PO4·2H2O 和 Na2SeO3·5H2O 作为肥源,均为分析纯级别。底肥按照每公斤土壤施入0.2 g N和 0.2 g K2O,分别以尿素、氯化钾(分析纯)作为肥源,并于拔节期每盆追施尿素 1.0 g。每盆播种12粒种子,定株8棵。成熟期进行取样,将小麦植株样品分为籽粒、颖壳、茎叶和根部,分别于105 ℃下杀青30 min,60 ℃恒温烘干、称重、粉碎,过20目筛,以备分析使用。土壤样品采集后于室内阴凉处自然风干,除去植物残体后磨碎过100目筛,待测。
1.3 测定指标和方法
1.3.1 小麦各部位硒含量测定
参照国家标准方法[12]采用原子荧光分光光度法测定:称取待测样品 0.500 0 g(精确到 0.000 1 g)于三角瓶中,加 10 mL混合酸 HNO3-HClO4(4∶1)放置过夜,于电热板上180 ℃消解至试样澄清,冷却后加入10 mL HCl(1∶1)进行还原,继续加热至溶液剩余3 mL左右,用去离子水定容至 25 mL容量瓶中,过滤后利用原子荧光光度计(AFS-8220,中国)测定硒含量。
1.3.2 土壤总硒含量测定
参照刘新伟等[13]的方法采用(HNO3-HClO4(3∶2)) 消解,后续步骤同植物样硒含量的测定方法。
1.3.3 土壤硒形态含量测定
参照吴少尉等[14]的方法采用连续浸提法进行测定,具体步骤如下:
(1)水溶态(Ws):称取 2.000 0 g 土壤样品,放入干净的聚乙烯离心管中,加入10 mL 0.25 mol·L-1HCl,25 ℃ 200 r·min-1恒温震荡 1 h,然后4 000 r·min-1离心 10 min,上清液消煮后待测;
(2)交换态(Ex/Cb):在(1)含有沉淀的离心管中,加入 10 mL 0.7 mol·L-1KH2PO4(pH 5.0),25 ℃ 200 r·min-1恒温震荡 4 h,然后 4 000 r·min-1离心10 min,上清液消煮后待测;
(3)铁锰氧化物结合态(Fe/Mn):在(2)含有沉淀的离心管中,加入 10 mL 2.5 mol·L-1HCl,置于 90 ℃的恒温水浴中加热 50 min,间歇振荡,然后4 000 r·min-1离心 10 min,上清液消煮后待测;
(4)有机物硫化物结合态及元素态(Om-Su/El):在(3)含有沉淀的离心管中,加入 8 mL 5% K2S2O8和 2 mL HNO3(1∶1),置于 95 ℃恒温水浴中加热3 h,并间歇振荡,然后4 000 r·min-1离心 10 min,上清液消煮后待测;
(5)残渣态(Res):将(4)残渣用少量去离子水多次全部转移到150 mL 三角瓶中。
不同提取态硒含量的后续步骤参照土壤硒含量的测定方法。
1.4 数据计算、统计与分析方法
植株养分累积量 = 植株养分含量 × 干物质重
Se迁移系数(根系-茎叶)= 茎叶Se含量/根Se含量
Se迁移系数(茎叶-颖壳)= 颖壳Se含量/茎叶Se含量
Se迁移系数(颖壳-籽粒)= 籽粒Se含量/颖壳Se含量
试验数据采用 DPS 7.05 版进行双因素方差分析和F值检验,采用Duncan法(P<0.05)进行多重比较,采用Sigmaplot 10.0 软件作图。
2 结果与分析
2.1 磷硒配施对郑麦9023植株籽粒产量及产量构成要素的影响
如表2可知,施磷、施硒及磷硒交互对郑麦9023的穗数、穗粒数及籽粒产量的影响均达到显著或极显著水平(P<0.05或P<0.01);磷硒交互对其千粒重存在显著效应(P<0.05)。
与Se0处理相比,Se1处理可显著降低P0水平下郑麦9023的穗数、千粒重及P160水平下的籽粒产量,其降幅分别为24.0%、15.2%和30.8%,但显著提高了P80和P160水平下的穗粒数及P80水平下的籽粒产量,其增幅分别为 20.1%、 16.6%和16.6%(表2)。无论施硒与否,与P0处理相比,P80和P160处理均显著提高了郑麦9023穗数、穗粒数和籽粒产量,且P160显著提高了Se1水平下的千粒重,其增幅为15.5%。穗数和千粒重分别在Se1P160和Se0P0处理达到最大值,而籽粒产量和穗粒数均在Se1P80处理下达到最大值。

表2 磷硒配施对郑麦9023籽粒产量及其构成的影响Table 2 Effect of P and Se application on grain yield its components of Zhengmai 9023
2.2 磷硒配施对郑麦9023植株各部位硒含量及硒迁移系数的影响
如表3可知,施磷、施硒及磷硒交互对郑麦9023各部位硒含量的影响均达到极显著水平 (P<0.01)。施磷、施硒及磷硒交互对硒从茎叶到颖壳、颖壳到籽粒迁移系数的影响均达到显著或极显著水平(P<0.05或P<0.01),而只有施硒处理对硒从根系到茎叶迁移系数的影响达到极显著水平(P<0.01)。
郑麦9023成熟期各部位硒含量顺序为根系>籽粒>茎叶>颖壳(表3)。无论施磷与否,Se1处理较Se0处理均显著提高了小麦各部位的硒含量,其增幅范围为240.0%~1 516.5%。在Se0水平下,施磷对小麦各部位的硒含量无显著影响。在Se1水平下,施磷则显著降低了各部位的硒含量,其降幅范围为36.3%~72.7%。各部位硒含量均在Se1P0处理下达到最大值。

表3 磷硒配施对郑麦9023各部位硒含量和硒迁移系数的影响Table 3 Effect of P and Se application on the Se concentration and migration coefficient in different parts of Zhengmai 9023
与Se0处理相比,Se1处理显著降低了P0和P80水平下硒从根系向茎叶的迁移系数,其降幅分别为51.9%和35.7%,但显著增加了P160水平下硒从茎叶向颖壳的迁移系数以及三个施磷水平下硒从颖壳向籽粒的迁移系数,其增幅范围为18.2%~55.7%。在Se0水平下,P80较P0处理显著增加了硒从茎叶向颖壳的迁移系数,其增幅为34.3% ,但显著降低了硒从颖壳向籽粒的迁移系数,其降幅为21.2%;在Se1水平下,P160较P0处理显著增加硒从茎叶向颖壳的迁移系数,其增幅为51.4%,P80和P160显著降低硒从颖壳向籽粒的迁移系数,其降幅分别为21.3%和29.0%。
2.3 磷硒配施对郑麦9023植株各部位硒累积量的影响
如表4可知,施磷、施硒及磷硒交互对郑麦9023籽粒、颖壳、茎叶和根系以及整株硒累积量的影响均达到显著或极显著水平(P<0.05或P<0.01)。
在Se0水平下,Se在郑麦9023成熟期各部位的累积量为茎叶>籽粒>根>颖壳;在Se1水平下,Se在郑麦9023成熟期各部位的累积量为籽粒>茎叶>根>颖壳(表4)。Se1较Se0处理均显著提高三个施磷水平下郑麦9023各部位以及整株硒累积量,其增幅范围为224.5%~ 1 125.2%,且提高了籽粒和根中硒累积量的比例,但降低茎叶中硒累积量的比例。在Se0水平下,施磷对籽粒、颖壳和根系的硒累积量无显著影响,但P80和P160处理显著提高茎叶和整株硒累积量,其增幅范围为167.5%~215.6%,且提高了颖壳和根中硒累积量的比例,但降低茎叶中硒累积量的比例。在Se1水平下,与P0处理相比,P80和P160处理显著提高小麦各部位以及整株硒累积量,其增幅范围为27.0%~114.0%,且提高了茎叶和根中硒累积量的比例,但降低籽粒中硒累积量的比例。颖壳、茎叶和根系硒累积量最大值均在Se1P160处理,籽粒和整株硒累积量的最大值出现在Se1P80处理。籽粒硒累积量分配比例的最大值出现在Se1P0处理;颖壳和根中硒累积量分配比例的最大值均出现在Se1P160处理;茎叶硒累积量分配比例的最大值出现在Se0P0处理。
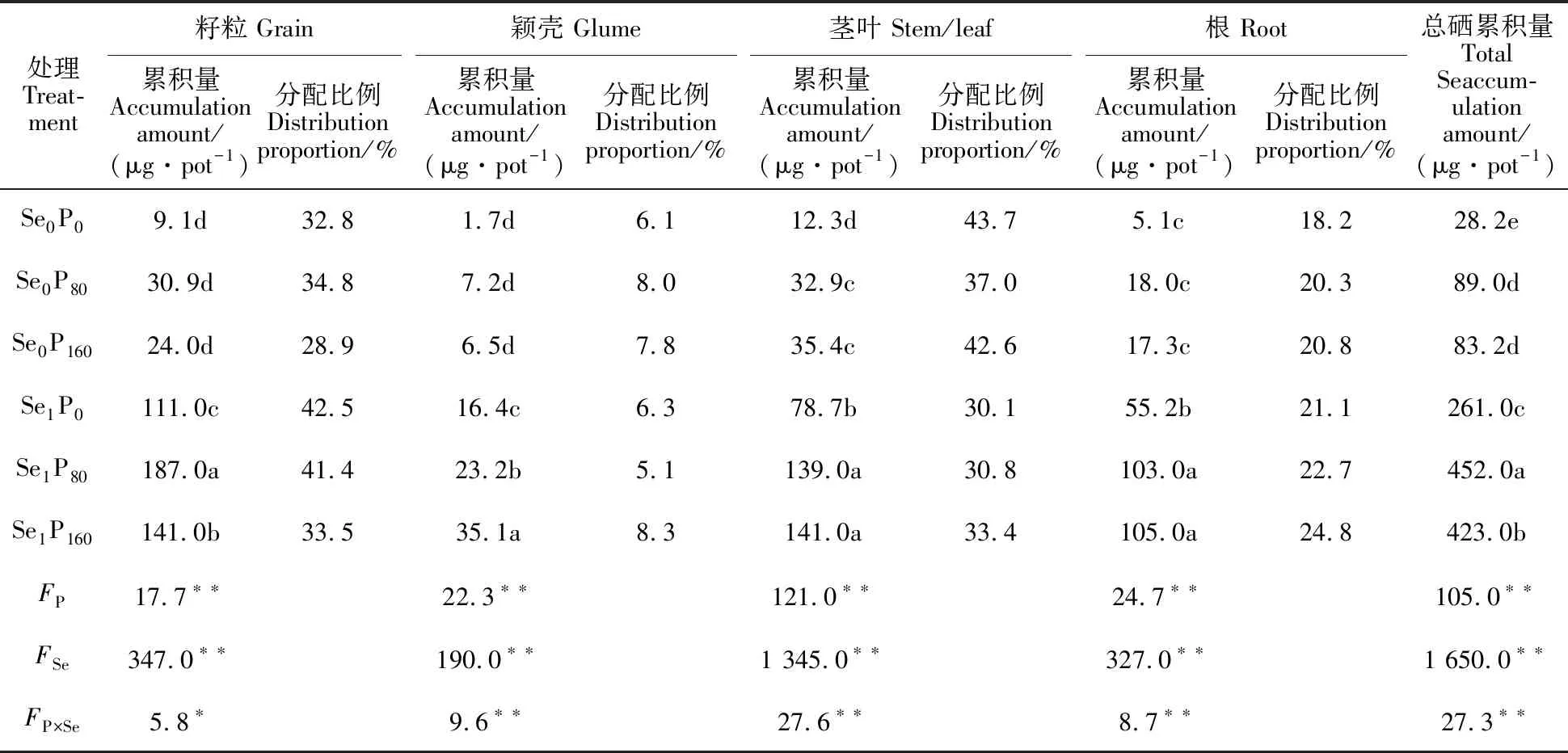
表4 磷硒配施对郑麦9023植株各部位硒累积量和分配比例的影响Table 4 Effect of P and Se application on the accumulation and distributed proportion of Se in each tissue of Zhengmai 9023
2.4 磷硒配施对土壤不同形态硒含量及比例的影响
施磷、施硒及磷硒交互对土壤中不同形态硒含量的影响均极显著(表5)。施硒可显著提高三个施磷水平下土壤中各种形态硒含量,且提高水溶态(Ws)、可交换态(Ex/Cb)硒含量所占的比例,但降低了铁锰氧化物结合态(Fe/Mn)、有机硫化物结合态及元素态硒(Om-Su/EI)含量所占的比例。在Se0水平下,施磷对土壤水溶态硒(Ws)、可交换态硒(Ex/Cb)、铁锰氧化物结合态硒(Fe/Mn)、有机物硫化物结合态及元态硒(Om-Su/El)含量无显著影响,但施磷增加了水溶态(Ws)、可交换态(Ex/Cb)及铁锰氧化物结合态硒(Fe/Mn)含量所占的比例,降低了有机物硫化物结合态及元素态硒(Om-Su/El)所占的比例。在Se1水平下,P80和P160处理较P0处理显著增加了水溶态硒(Ws)、有机物硫化物结合态及元素态硒(Om-Su/El)及残渣态硒(Res)含量和所占的比例,其含量增幅分别为56.9%和78.4%、 10.9%和28.7%、 19.0%和23.8%;但显著降低了可交换态(Ex/Cb)及铁锰氧化物结合态硒 (Fe/Mn)含量和所占的比例,其含量降幅分别为8.8%~21.9%、 8.3%~13.2%。
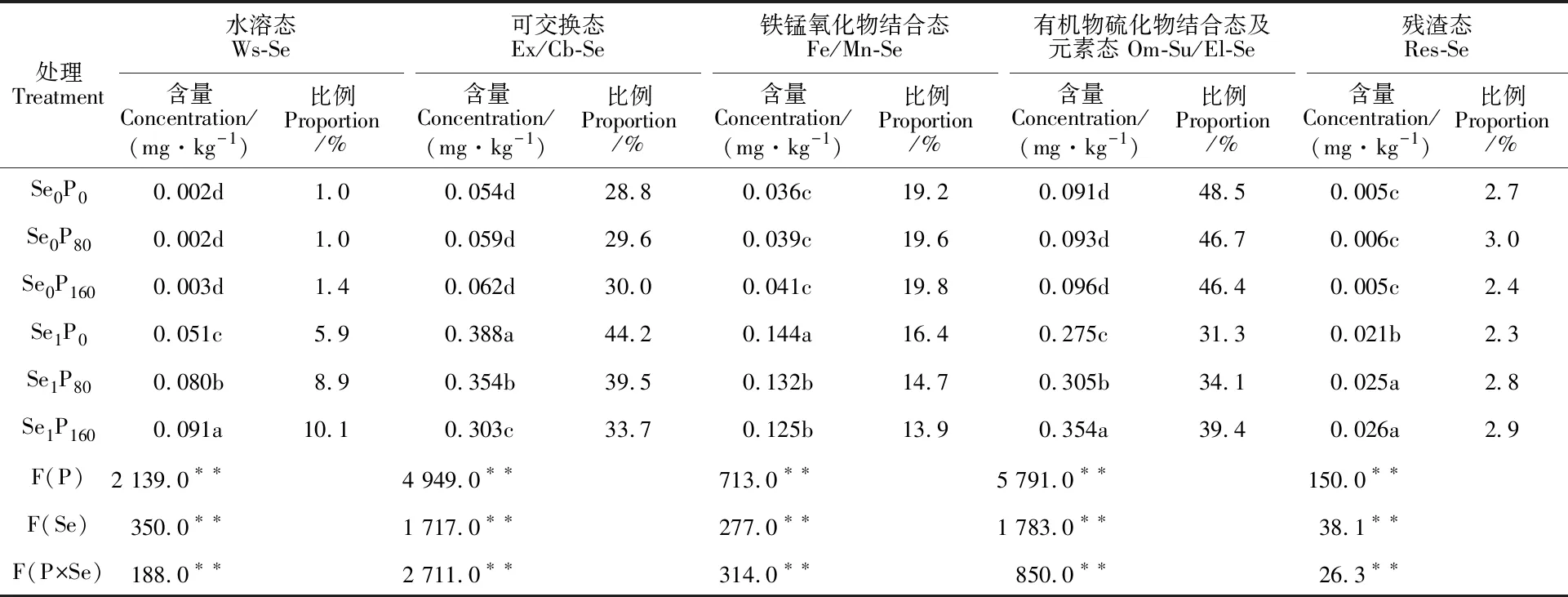
表5 磷硒配施对土壤不同形态硒含量和比例的影响Table 5 Interactive effect of P and Se application on the concentration and proportion of Se in different chemical forms in soil
2.5 籽粒硒含量、籽粒硒累积量以及整株硒累积量与土壤不同形态硒含量的相关性分析
如表6所示,籽粒以及整株硒累积量与不同形态硒含量均存在极显著的正相关。籽粒硒含量与土壤可交换态硒(Ex/Cb)和铁锰氧化物结合态硒(Fe/Mn)之间存在极显著或显著的正相关 关系。
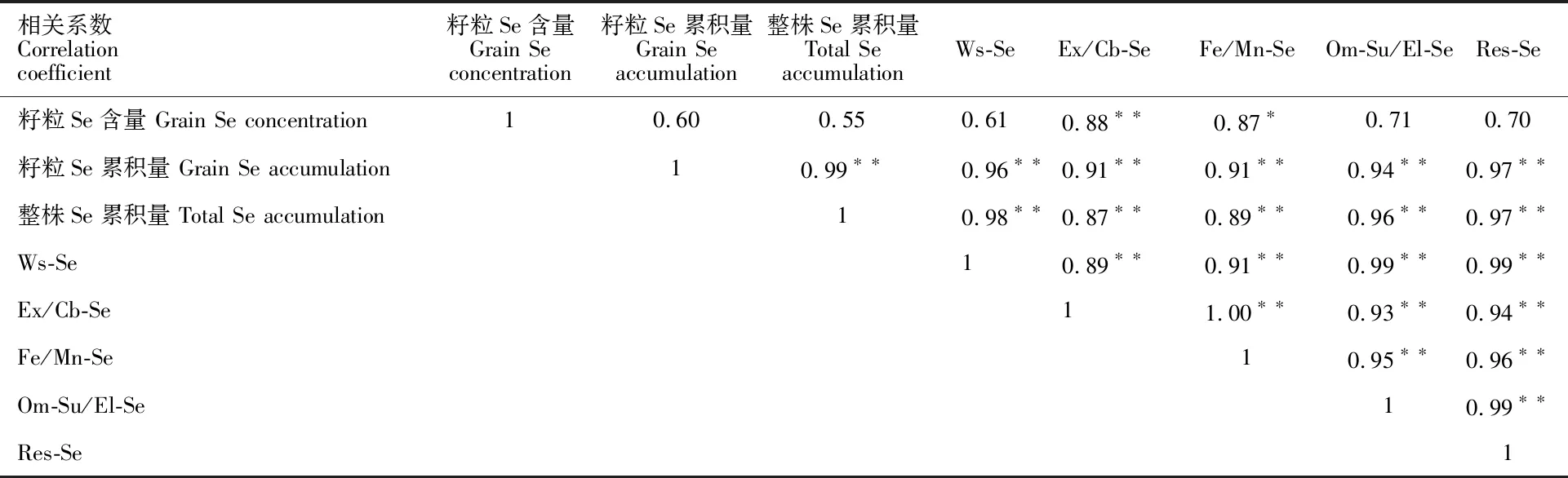
表6 籽粒硒含量、籽粒硒累积量和整株硒累积量与土壤不同形态硒含量相关性分析Table 6 Correlation analysis between grain Se concentration,grain Se accumulation,whole plant Se accumulation and Se concentrations in different chemical forms in soil
3 讨 论
3.1 磷硒配施可促进郑麦9023吸收土壤硒,但抑制硒向地上部的分配以及向籽粒的迁移和分配
本试验中,在Se1水平下,与不施磷相比,施磷显著增加了各器官和整株硒累积量,且最大值出现在P80处理。说明磷硒配施可促进郑麦9023吸收土壤硒,这与赵文龙[15]发现施磷能促进小白菜吸收硒的结果相一致。Liu等[16]研究发现,适量施磷能促进水稻地上部对四价硒的累积,但过量施磷却抑制了地上部对四价硒的累积,且磷不同施用量均抑制了水稻地下部硒的吸收。而Hopper等[17]在水培条件下的试验表明,施磷能显著降低黑麦草对硒的吸收。Mora等[18]研究也表明,磷可以减少植物对硒的吸收,进而缓解硒对植物的抑制作用。其他研究表明,施磷可促进植物吸收硒,一方面可能是由于磷酸根和亚硒酸根在土壤胶体上存在竞争吸附作用,促使土壤溶液解析出更多的硒供植物吸收,另一方面可能是施磷促进植物蒸腾作用以及根系的生长,进而促进植物对硒的吸收[19-20]。本试验在Se1水平下,施磷显著降低了郑麦9023各部位硒含量。赵文龙[21]等认为,施磷会导致小白菜地上及地下部硒含量下降,是由于磷的施入显著增加了小白菜生物量,进而引起的生物稀释效应所致。陈思杨等[22]研究发现,在极低硒含量条件下,施磷可显著增加水稻幼苗硒的转运系数。本试验中,硒含量的降低一方面可能由于施磷可促进小麦生长以及籽粒产量的显著增加,进而导致的稀释效应,另一方面可能由于施磷抑制了硒向地上部的分配以及向籽粒的迁移和分配。
3.2 磷硒配施可影响土壤不同硒形态之间的转化进而影响硒的有效性

猜你喜欢
江苏农业科学(2022年15期)2022-08-11
中国农学通报(2022年13期)2022-05-31
生态与农村环境学报(2022年3期)2022-03-27
麦类作物学报(2022年1期)2022-03-01
现代畜牧科技(2021年4期)2021-12-05
南昌航空大学学报(自然科学版)(2021年1期)2021-06-02
山东建筑大学学报(2020年5期)2020-09-16
农民致富之友(2019年33期)2019-12-20
江苏农业学报(2019年2期)2019-09-10
