
茶树体内咖啡碱生物代谢研究进展
2020-07-31 08:06闫振黄健垚高路王登良
中国茶叶 2020年7期
闫振,黄健垚,高路,王登良
1.广东德高信食品加工有限公司,513000;2.华南农业大学茶学系,510642
咖啡碱(1,3,7-三甲基黄嘌呤,Caffeine)是衍生于嘌呤核苷酸的一种嘌呤生物碱,已被发现存在于茶、咖啡和可可等60多种不同的植物中[1]。含咖啡碱植物一方面通过化感或自毒作用从种皮中释放咖啡碱到土壤中从而抑制邻近植物种子的萌发[2];另一方面通过化学防御作用抵御捕食性动物采食其嫩叶、果实等[3]。咖啡碱作为一种行为活性物质被广泛使用,并有很大一部分人在饮食中长期使用咖啡碱。咖啡碱是茶叶滋味物质的主要组成成分,除了其本身的苦味之外,还可与茶黄素、茶红素以氢键缔合形成络合物提高茶汤品质[4-5]。饮茶则有兴奋神经中枢、缓解疲劳、助消化、强心解痉和利尿等功效[6]。茶树中的咖啡碱主要集中分布在新梢中,且以鲜嫩的芽叶含量最高,并随芽叶的老化而逐渐减少,这种分布规律与咖啡碱在茶树体内的生物合成与代谢机制密切相关[7]。本文综述了茶树中咖啡碱的生物代谢研究进展,包括咖啡碱在茶树中的含量分布、生物合成与分解代谢途径以及相关基因的表达研究。
一、咖啡碱在茶树体内的生物合成
1.咖啡碱在茶树中的分布
茶树中的咖啡碱主要分布在叶组织并集中在新梢中,尤其以一叶中含量最高并随叶质的老化而逐渐降低。除叶片外,茶花、茶树茎梗、茶籽和根部也含有咖啡碱,但其含量甚微[8]。茶花的花瓣和雄蕊可以合成咖啡碱,且在茶花盛开前期的生物合成活性最高[9];茎梗中的咖啡碱以茎上部分居多并随着茎梗的硬化而呈下降趋势,茎下部含量仅为0.01%左右;在茶树果实即茶籽完全成熟之前,咖啡碱的含量在显著增加[10],但茶籽中的咖啡碱只分布在种皮中,占鲜重的0.09%左右;根部不足0.01%(表1)。Ashihara 等[8]通过[8-14C]腺嘌呤示踪实验证明咖啡碱的合成主要发生在茶树幼叶中并在叶片中积累,并发现咖啡碱的合成前体可可碱主要存在于茶树嫩梢中;大量的[8-14C]腺嘌呤在茶树幼叶中合成可可碱并进一步合成咖啡碱;茶树体内超过99%的咖啡碱存在于叶片中。
茶树中咖啡碱的含量与茶树品种、外部环境因子如季节、温度、光照和土壤条件等有关。王绍梅等[11]对越南大叶、安溪水仙、黔湄等12份茶树品种的烘干茶叶咖啡碱含量测定表明,越南大叶含咖啡碱含量最高(4.96%),安溪水仙最低(3.41%),从而证明大叶种茶树一般比中小叶种茶树的咖啡碱含量高,乔木型茶树(4.54%)一般比小乔木型(4.16%)和灌木型茶树(4.33%)的咖啡碱含量高。对春夏秋不同季节茶叶的咖啡碱含量测定表明,夏季鲜叶所制茶样中的咖啡碱含量(4.7%)一般比春季茶叶(4.5%)和秋季茶叶(4.2%)含量高。这与夏季温度高,茶树体内生物代谢旺盛密切相关[12]。生产经验和科学研究表明,适当遮阴一方面可抑制咖啡碱的分解,另一方面可促进茶树新梢中咖啡碱的合成,显著提高茶叶中咖啡碱的含量[13]。矿质元素是生命体生长的必要营养物质,镁离子作为生物蛋白酶的激活剂在生物学反应中起着重要的作用,S-腺苷蛋氨酸是生物碱合成过程中重要的甲基供体,而镁离子则是促进蛋氨酸活化酶催化合成S-腺苷蛋氨酸的必要激活因子。钼离子是酸式磷酸酶的专一性抑制剂,也是硝酸还原酶的辅基,对氮代谢有明显的影响,实验表明用较高浓度钼离子处理后的茶苗其咖啡碱含量会明显上升[7]。
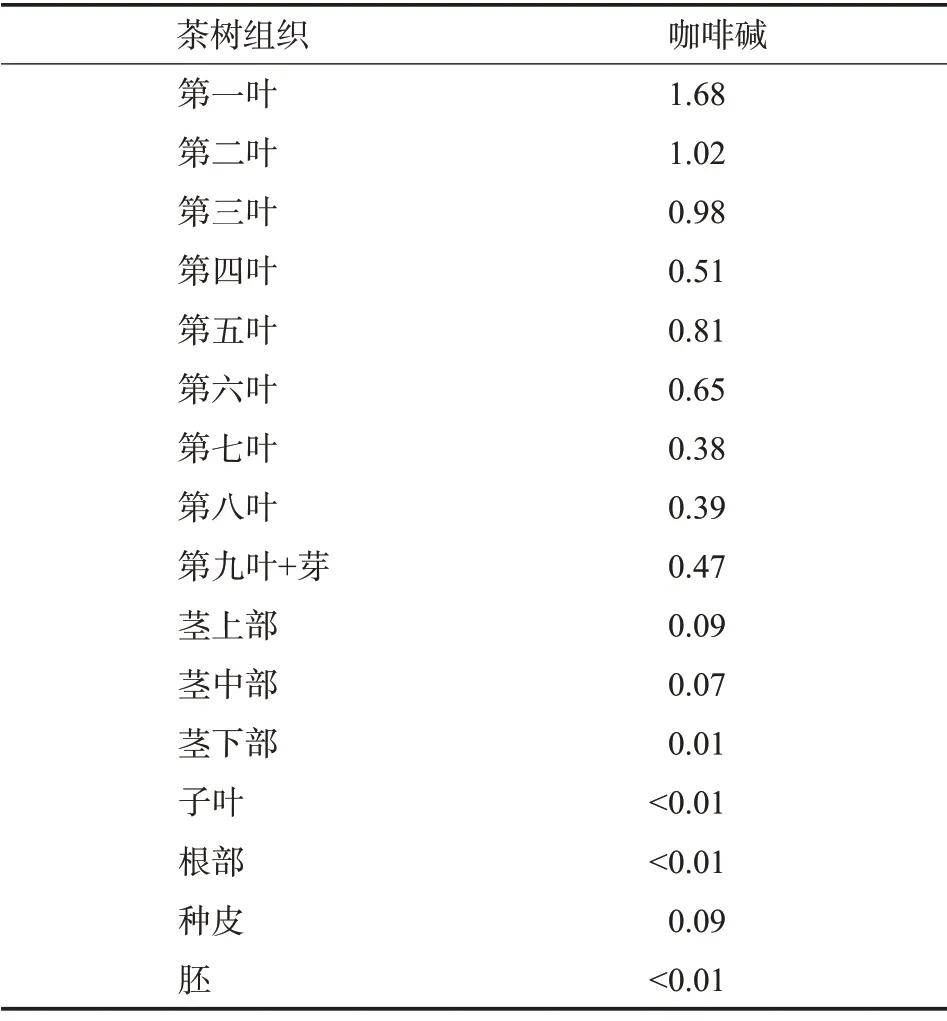
表1 茶树幼苗不同组织中咖啡碱含量%
2.咖啡碱在茶树体内的合成途径
(1)从黄苷到咖啡碱的合成
研究者们通过14C同位素示踪、底物专一性和重组N-甲基转移酶等实验对茶树、咖啡等植物体内的咖啡碱代谢反应研究证明,咖啡碱的骨架——黄嘌呤来自于嘌呤核苷,咖啡碱的主要合成过程由1步核苷酶反应和3步甲基化反应完成,其主要合成途径为:黄苷→7-甲基黄苷→7-甲基黄嘌呤→可可碱(3,7-二甲基黄嘌呤)→咖啡碱(1,3,7-三甲基黄嘌呤)(图1)[14-15]。
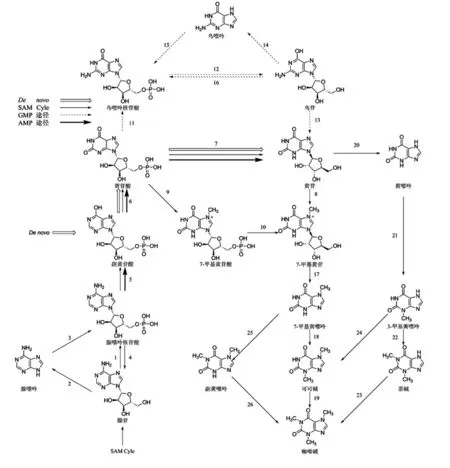
图1 咖啡碱合成途径
在黄苷-N-甲基转移酶的作用下,黄苷的7-N-甲基化并转化为7-甲基黄苷;7-甲基黄苷在黄苷酶的催化下水解生成7-甲基黄嘌呤;7-甲基黄嘌呤经可可碱合成酶(单甲基黄嘌呤-N-甲基转移酶)的作用进一步甲基化转化成可可碱;最后,可可碱在咖啡碱合成酶(二甲基黄嘌呤-N-甲基转移酶)的催化作用下进行第三步甲基化最终生成咖啡碱。
Ashihara 等[16]发现在茶树幼叶中还存在另外2条咖啡碱合成路径,其一为:黄嘌呤→3-甲基黄嘌呤→茶碱→咖啡碱;其二为:黄苷→7-甲基黄苷→7-甲基黄嘌呤→副黄嘌呤→咖啡碱。体外研究发现,以副黄嘌呤为底物,咖啡碱合成酶的催化活性最高,表明相对于可可碱和7-甲基黄嘌呤,副黄嘌呤是咖啡碱合成酶的最佳底物。但是,同位素示踪实验表明,在茶树体内,咖啡碱的合成以催化可可碱为主,几乎不经过副黄嘌呤[17]。
(2)黄苷的合成
黄苷是嘌呤生物碱合成的起始底物,目前已发现4条合成途径:腺嘌呤核苷酸合成途径(AMP途径)、鸟嘌呤核苷酸途径(GMP途径)、S-腺苷-L-蛋氨酸循环途径(SAM途径)以及二次利用途径(De novo途径)[18](图1)。
用于咖啡碱合成的黄苷有一部分来自于腺嘌呤和鸟嘌呤核苷酸池,这些核苷酸池是通过二次利用和补救途径产生的。
AMP途径:AMP(腺嘌呤核苷酸)→IMP(副黄苷酸)→XMP(黄苷酸)→黄苷,其过程为AMP 在脱氨酶的作用下脱去氨基转化为IMP,IMP在脱氢酶的作用下生成XMP,继而在5'-核苷酶的作用下生成黄苷[17]。
GMP途径:GMP(鸟嘌呤核苷酸)→鸟苷→黄苷,其过程为GMP 经5'-核苷酶的催化生成鸟苷,继而在脱氨酶的作用下生成黄苷。鸟苷除了生成黄苷进入嘌呤碱合成途径,还有一部分在鸟苷核苷酶的作用下转化为鸟嘌呤进入鸟嘌呤核苷酸池[19]。
SAM途径:腺苷→AMP→IMP→XMP→黄苷,SAM是咖啡碱合成途径中3步甲基化反应的甲基供体。在此过程中,SAM被转化为S-腺苷-L-半胱氨酸(SAH),然后水解为半胱氨酸和腺苷。半胱氨酸通过SAM 循环途径来补救SAM 水平,腺苷则从循环中释放出来继而直接转化为AMP,AMP进入黄苷合成途径或先经腺苷核苷酶作用转化为腺嘌呤,再经腺嘌呤磷酸核糖转移酶作用转化为AMP进入黄苷合成途径。腺苷除了生成AMP进入嘌呤碱合成途径,还有一部分在腺苷核苷酶的作用下转化为腺嘌呤进入腺嘌呤核苷酸池[18]。
De novo途径:IMP→XMP→黄苷,在茶树嫩叶中,5'-核苷酶催化甘氨酸生成IMP,IMP在脱氢酶的作用下生成XMP,继而在5'-核苷酶的作用下生成黄苷。其中,IMP脱氢酶是该途径的控速因子,决定了De novo途径的效率[20]。
二、咖啡碱在茶树体内的分解代谢
咖啡碱的分解代谢最早发现于咖啡植物体内,随后大量的14C同位素标记示踪实验证明咖啡碱在茶树等植物体中的分解首先经过3步脱甲基化转化为黄嘌呤,即咖啡碱→茶碱→3-甲基黄嘌呤→黄嘌呤;继而,黄嘌呤通过传统的嘌呤代谢途径最终分解为CO2和NH3,即黄嘌呤→尿酸→尿囊素→尿囊酸→尿素→CO2和NH3[20](图2)。
咖啡碱(1,3,7-三甲基黄嘌呤)首先在7-N-脱甲基酶的作用下脱去7-甲基转化为茶碱(1,3-二甲基黄嘌呤);茶碱经1-N-脱甲基酶的催化再脱去1-甲基生成3-甲基黄嘌呤;在3-N-脱甲基酶的作用下,3-甲基黄嘌呤脱去3-N-甲基后转化为黄嘌呤。咖啡碱转化为黄嘌呤的分解途径在茶树等植物体和动物体中是相同的,但黄嘌呤的进一步分解因物种的不同而存在差异。在茶树等植物体内,黄嘌呤最初经黄嘌呤脱氢酶氧化生成尿酸;尿酸继而被尿酸氧化酶氧化成尿囊素;在尿囊素酶的催化下,尿囊素水解为尿囊酸;进一步在尿囊酸酶的作用下,水解生成尿素和乙醛酸;尿素在尿酶的作用下最终分解生成CO2和NH3。咖啡碱等嘌呤生物碱在茶树体内的分解代谢主要发生在老叶中,产生的尿酸、尿囊素可以从茶叶中转运出来,作为储备物质供再次利用。而人和动物体内因缺少尿囊素酶、尿囊酸酶和尿酶等一系列酶,故分解尿酸的能力不及植物,常以尿酸为代谢产物排出体外[17]。
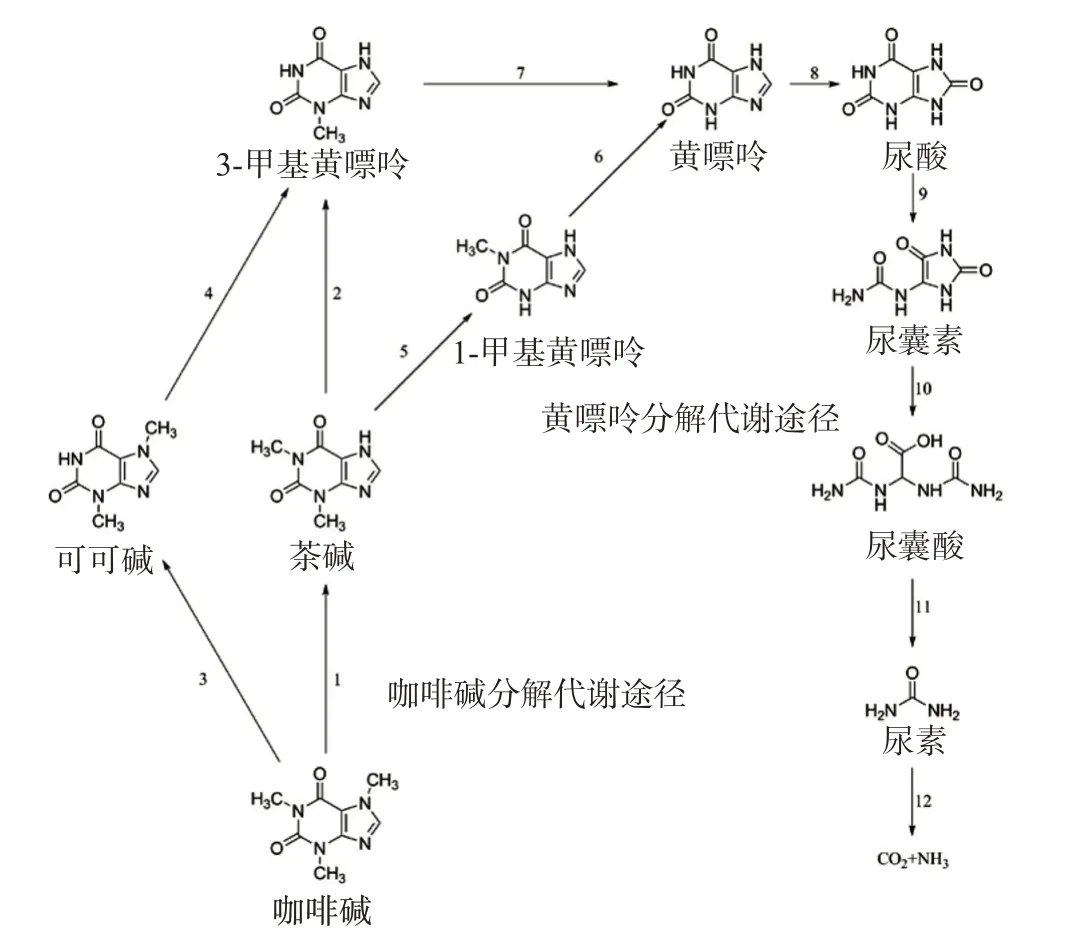
图2 咖啡碱分解代谢途径
[8-14C]示踪实验证明茶碱分解代谢的速度比咖啡碱快,这说明咖啡碱转化为茶碱的过程是咖啡碱分解代谢途径中主要的控速因子,也是咖啡碱在茶叶中积累的重要原因。另一方面,从咖啡碱的合成和分解过程可以明确咖啡碱的代谢与核酸特别是嘌呤核苷酸、蛋白质的代谢密切相关,故而在核酸、蛋白质代谢旺盛的茶树新梢中,咖啡碱合成代谢旺盛、分解代谢缓慢,故含量高。随着叶质的老化,核酸、蛋白质的分解代谢加强,咖啡碱的合成代谢减慢并以分解代谢为主,因此,老叶中的咖啡碱含量比新梢中的低。
三、咖啡碱合成酶基因
S-腺苷-L-蛋氨酸(SAM)广泛分布在生物体内,是嘌呤碱发生甲基化的甲基供体。而保守结构域A、B和C是大多数植物中甲基供体SAM的靶基因结合位点,在识别嘌呤环N-甲基化位置和催化甲基化反应中起着重要的作用[21]。咖啡碱的生物合成包括3步甲基化过程,由3种不同的SAM依赖型N-甲基转移酶催化完成。根据其底物特异性可分为3种类型[22]:7-甲基黄苷合成酶(催化黄苷形成7-甲基黄苷)、可可碱合成酶(3,7-二甲基黄嘌呤合成酶,催化7-甲基黄嘌呤形成可可碱)、咖啡碱合成酶(1,3,7-三甲基黄嘌呤合成酶,催化可可碱形成咖啡碱)。
7-甲基黄苷合成酶是一类底物专一性的酶,参与第一步甲基化反应催化黄苷形成7-甲基黄苷。Mizuno 等[23]第一次从咖啡叶片中克隆得到7-甲基黄苷合成酶基因(CmXRS1),其编码cDNA序列长1 307对碱基,编码372个氨基酸。
第二步甲基化反应是7-甲基黄嘌呤转化为可可碱,由可可碱合成酶催化完成。Suzuki 等[24]在茶叶粗提取物中首次发现并证实了7-甲基黄嘌呤N-甲基转移酶(可可碱合成酶)和可可碱N-甲基转移酶(咖啡碱合成酶)的催化活性。可可碱合成酶也具有底物专一性,仅催化7-甲基黄嘌呤发生3-N-甲基化并转化为可可碱。在咖啡植物体内已发现的可可碱合成酶编码基因有CTS1、CaMXMT1、CTS2、CaMXMT2和BTS1等,其氨基酸序列相似度达80%,均具有保守的SAM 结合位点A、B'、C和YFFF等结构域[14,25]。CgcTS基因是从广西大瑶山秃房茶品种(Camellia gymnogynaChang)茶树幼叶中克隆得到的1个可可碱合成酶基因,当以不同的嘌呤衍生物为底物时,CgcTS蛋白酶催化7-甲基黄嘌呤的活性最高,其次是黄苷,而以可可碱、咖啡碱和茶碱为底物时,该酶未显示出活性。另外,CgcTS基因在mRNA 水平和蛋白水平的表达模式是一致的,两者均在幼嫩的一叶中表达水平高而在老化的第四叶中表达水平低,这与可可碱在秃房茶中的分布规律也是一致的[26]。
咖啡碱合成酶参与咖啡碱生物合成的最后两步,既可催化7-甲基黄嘌呤转化为可可碱,也可催化可可碱形成咖啡碱。TCS1是最初从茶树(Camellia sinensis)幼叶中提取纯化得到的咖啡碱合成酶,其编码cDNA 由1 483对碱基组成,编码369个氨基酸。体外实验证明TCS1的底物特异性与纯化的天然酶十分相似,主要催化单甲基黄嘌呤和二甲基黄嘌呤的3-N-甲基化和1-N-甲基化,即以7-甲基黄嘌呤为底物催化发生3-N-甲基化形成可可碱,以可可碱为底物催化发生1-N-甲基化形成咖啡碱。此外,TCS1基因在幼叶中的转录水平很高,而在老叶中的转录水平极低,TCS1的基因表达模式和TCS1酶活特性与咖啡碱在茶叶中的含量分布是一致的[27-29]。
此外,一些茶种体内的生物碱以可可碱为主,如可可茶、后轴茶、红芽茶和秃房茶,这些茶树体内的咖啡碱含量显著低于可可碱的含量。Yoneyama 等[30]在这些高可可碱低咖啡碱茶树中发现尽管其咖啡碱合成酶氨基酸序列与咖啡植物有高达80%的相似性,但同源性较低,咖啡碱合成酶在这些茶种中可能只具有3-N-甲基化活性而不具有1-N-甲基化活性。HycCS1和CctCS1分别是从低咖啡碱的红芽茶品种和可可茶品种中克隆得到的咖啡碱合成酶基因,其cDNA序列均为1 098 bp,编码365个氨基酸,仅第227位(Glu227Lys)和第287位(Arg287His)氨基酸不同。其编码蛋白酶HycCS1和CctCS1均只能催化可可碱的形成而不能以可可碱为底物合成咖啡碱,而HycCS1的催化活性要高于CctCS1[31]。与TCS1相比,HycCS1和CctCS1的第221位氨基酸均为组氨酸,TCS1的为精氨酸,而组氨酸在茶树体中扮演着重要的底物识别作用[31],这与HycCS1和CctCS1只具有3-N-甲基化活性催化7-甲基黄嘌呤形成可可碱而无法进一步形成咖啡碱有着重要的关系。
四、展望
咖啡碱对人类的精神运动和认知能力有积极的影响,包括在习惯性摄入低至中等剂量咖啡碱的情况下提高警觉性、能量和兴奋;相反,高剂量的咖啡碱会引起诸如血压升高、紧张、心悸、焦燥不安、失眠和胚胎畸形等负面影响,且长期食用会使人产生依赖感,不适于儿童、孕妇、老人及神经衰弱者[6,32]。而低咖啡碱茶产品可以满足这些特殊人群对茶叶的需求,也可充分发挥茶叶的保健功效。因此,低咖啡碱茶产品越来越受到茶叶市场的重视。目前国内外主流的脱茶叶咖啡碱的方法是利用物理、化学、生化等加工工艺,但这些方法多用于深加工茶产品,加工成本较高且存在一定的溶剂残留,在很大程度上影响成品茶的感官品质[33]。而福建南部山区的红芽茶、广东南昆山的可可茶、广西大瑶山的秃房茶等则是天然的低咖啡碱高可可碱茶树资源,其成品茶不仅可以避免过多摄入咖啡碱带来的负面影响还省去了脱咖啡碱的加工成本[26,31]。此外,这些低咖啡碱茶树因其又富含可可碱和非表型儿茶素而具有较突出的抗氧化等生物活性和抗炎等保健功效。茶树等植物体内包括嘌呤核苷酸代谢、细胞间易位和在特定细胞部位(如叶绿体和液泡)积累的咖啡碱合成与分解的细胞代谢机制已经明确。这为研究天然低咖啡碱茶树资源的咖啡碱代谢机制提供了理论基础,同时为繁育天然低咖啡碱茶树品种提供了技术支持。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-21
山东农业科学(2021年1期)2021-12-09
教育周报·教育论坛(2020年3期)2020-10-21
分析化学(2020年9期)2020-09-14
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
家庭医药·快乐养生(2017年6期)2017-06-16
科技资讯(2017年12期)2017-06-09
家庭科学·新健康(2016年5期)2016-05-12
分析化学(2014年7期)2014-12-13
