
大麦EMS突变体库构建和突变体筛选
2020-07-30 08:08栾海业张英虎乔海龙杨红燕沈会权
麦类作物学报 2020年5期
栾海业,徐 肖,陶 红,臧 慧,张英虎,乔海龙,杨红燕,陈 健,陈 和,沈会权
(江苏沿海地区农业科学研究所,江苏盐城 224002)
大麦(HordeumvulgareL.)是世界上四大禾谷类作物之一,具有生育期短、抗病和耐逆(耐旱、耐盐碱、耐贫瘠)等特点,对气候环境适应性较广泛;大麦主要用途有饲料、麦芽啤酒、食粮及工业原料等,并且是青藏高海拔地区最主要的粮食作物。大麦是二倍体自花授粉作物,染色体数为2n=2x=14,基因组大小为5.1 Gb,目前大麦基因组图谱已公布,2012年,多国科学家利用Illumina和Roche 454测序平台,构建了4.98 Gb的大麦基因组物理图谱,并揭示了3.2×104个基因的排列顺序和结构[1];2017年,国际大麦测序联盟综合运用染色体构象作图等多种最先进的测序和组装技术,组装出了迄今最完整的大麦基因组物理图谱,并提供了3.9×104个基因的高可信度信息[2]。大麦具有遗传背景简单,遗传转化相对容易的特点,是麦类作物功能基因组研究的重要模式植物。
突变体库的创建是植物功能基因组学研究的重要基础,目前在拟南芥[3]、水稻[4]等模式作物上均已构建了饱和的突变体库,并利用突变体材料,克隆了大量的功能基因[5-6]。突变体库的构建还可以创造新的种质资源,丰富品种的遗传多样性,为新品种选育提供基础材料。甲基磺酸乙酯(ethyl methane sulfonate,EMS)化学诱变技术是构建植物突变体库的重要方法之一,具有诱变效率高,染色体畸变率较低,且多为点突变等优点,所以被广泛应用于各种作物突变体库的构建[7]。孙玉龙等[8]通过EMS对高产、抗白粉小麦盛农1号进行诱变,获得174个突变穗系,在M3代中得到18个稳定遗传的株系,其中4个株系小穗性状优于盛农1号;张 彬等[9]通过对糜子进行EMS处理,构建了变异类型丰富的糜子突变体库,筛选到104个形态性状变异株系;霍朋杰[10]利用EMS对裸燕麦进行诱变处理,获得了表型变异丰富的EMS诱变系,2 000个M2株系中,鉴定到196份表型突变材料,变异率为9.8%。不同作物突变体的获得为作物功能基因的克隆及验证提供了丰富的基础材料。
目前国内外已构建了多个大麦EMS突变体库并利用突变体开展了相关研究,如Caldwell等[11]对二棱大麦品种Optic构建了2个EMS诱变突变体库,获得了大约20 000个M2株系,并对其中的9 216个株系进行了定向诱导基因组局部突变技术(targeting induced local lesions in genomes,TILLING)分析;齐新丽等[12]对美国大麦品种Tamalpais进行EMS诱变处理,构建了表型变异丰富的突变体库,并利用TILLING技术成功获得了大麦茉莉酸受体基因(COI1)的突变体;吕瑞华等[13]利用EMS诱变技术,构建了大麦品种西引2号的TILLING筛选群体,并利用该群体对大麦抗病基因(EDR1)的突变体进行了检测。与拟南芥、水稻和小麦相比,大麦突变库的构建还远没有达到饱和状态,尤其是我国构建的大麦突变体库还较少,严重阻碍了我国大麦功能基因组学的发展;要获得具有自主知识产权的重要的功能基因,首先应以当地推广品种为材料自主构建大麦突变体库。本研究旨在利用抗黄花叶病的优质啤酒大麦品种苏啤6号构建大麦突变体库,为大麦功能基因组学研究和遗传育种提供新突变体材料。
1 材料与方法
1.1 试验材料
供试大麦品种苏啤6号由江苏沿海地区农业科学研究所利用浙皮1号为母本,用该所选育的单二大麦品种为父本,进行人工杂交组配,后经连续多年系谱选育而成。该品种为弱春性中熟二棱皮大麦,幼苗半直立,分蘖力中等偏上,株型较紧凑,耐肥抗倒性好,大穗大粒,后期熟相好,麦芽品质好,高抗大麦黄花叶病,目前是江苏及周边地区主栽大麦品种。
1.2 研究方法
1.2.1 EMS 诱变
选取籽粒均匀、饱满的苏啤6号种子6 000粒,用浓度为0.5%的EMS溶液浸泡,并在室温条件下于振荡器上350 r·min-1振荡10 h,种子用流水冲洗3 h后,自然晾干。
1.2.2 诱变种子的种植及性状调查
2015年11月将诱变种子播种于江苏沿海地区农业科学研究所南洋试验场,2016年6月收获大约2 400个M1代单株。2016年11月每个单株种子种植1行,行长1.5 m,行距25 cm,株距 5 cm,并设野生型苏啤6号为对照,对M2代株系表型进行全生育期调查记载,主要记录幼苗生长习性、叶形、叶色、分蘖力、株高、抽穗期、抗病性、棱型和穗型等性状,筛选形态明显变异的突变株行(以下称其为株系)。2017年11月将筛选出的变异材料继续播种于试验田中,鉴定其表型变异的稳定性。
1.2.3 M2代表型突变频率的计算
M2代突变频率=M2代突变株系/M2代总株系×100%
2 结果与分析
本试验用0.5%的EMS诱变处理大麦种子,萌发后的大麦幼苗生长明显受到影响,出苗比对照约迟5 d左右,并有黄化及白化苗现象,M1代单株成活率大约为40%。2 400个M1突变单株自交获得M2代,在正常播种条件下,对M2代株系的主要农艺性状及生物学性状进行调查,结果表明,经EMS诱变后,苏啤6号产生了丰富的表型变异,在所调查的幼苗生长习性、叶形、叶色、分蘖数、株高、抽穗期、棱型和穗型等性状中均发现了突变单株或株系(表1)。M2代共发现突变株系152个,突变频率为6.33%。
2.1 幼苗习性突变
野生型苏啤6号幼苗为半直立型,在调查的2 400个M2株系中,共发现36个株系的幼苗习性发生了5种类型(匍匐、白化、黄化、丛生、弯曲)的突变,总的突变频率为1.43%(表1)。其中,幼苗匍匐型突变株系有3个,突变频率为0.13%(图1A);幼苗发生白化死亡的株系有10个,突变频率为0.42%;幼苗整个植株或整个生育期发生黄化的突变株系有15个,突变频率为0.63%,该性状在M3代可以稳定遗传(图1B);幼苗丛生型突变株系有6个,突变频率为 0.25%(图1C);幼苗弯曲型突变株系有2个,突变频率为0.08%(图1D)。

A:幼苗匍匐;B:黄化苗;C:幼苗丛生;D:幼苗弯曲;E:野生型。
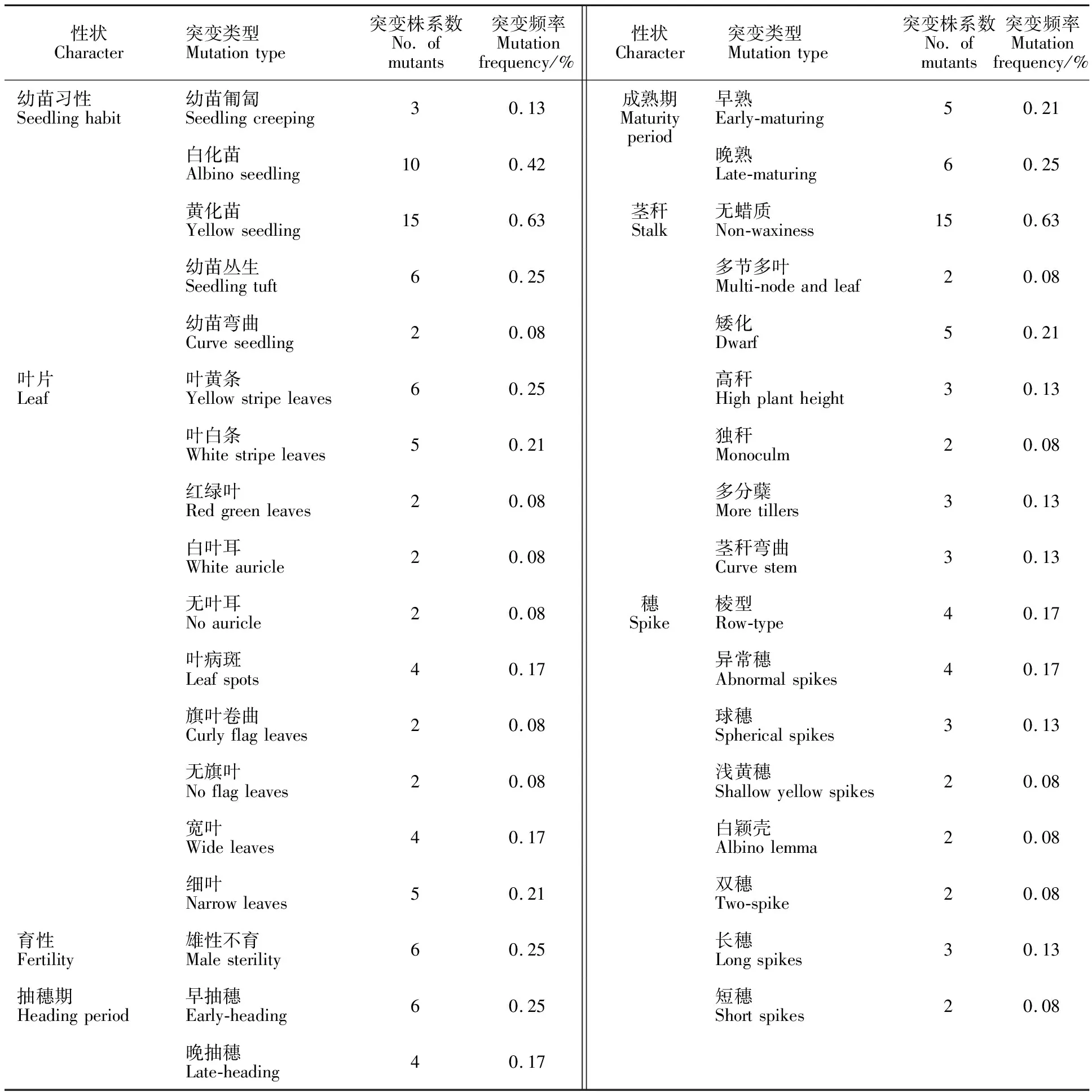
表1 EMS诱变M2代各种突变表型的突变频率
2.2 叶片突变
在2 400个M2代株系中,共发现34个株系的叶片发生了叶片黄条、白条、红绿叶、白叶耳、叶病斑、旗叶卷曲、无旗叶、宽窄叶以及无叶耳10种类型的变异,总的突变频率约为1.42%(表1)。叶黄条突变发生在分蘖期,部分叶片的边缘黄化,叶绿素含量明显减少,拔节期后,黄化叶片逐渐转为绿色,可以正常进行光合作用、抽穗和结实(图2A)。叶白条突变出现在幼苗期,返青拔节后部分植株死亡,部分可以逐渐恢复(图2B)。2个突变株系为红绿叶突变,叶片同时具有绿色、白色和红色,该性状不能稳定遗传到M3代(图2C)。白叶耳突变除叶耳颜色为明显的白色外,其他性状与野生型没有明显差别(图2D),M3验证该表型能够稳定遗传,M2群体中叶耳颜色正常植株与白叶耳植株的比例为3∶1。叶病斑突变有两种类型,一种是全生育期叶片有褐色的类病斑(图2E),另一种为全生育期所有绿叶都有白色病斑(图2F),这些突变体均不会导致叶片及植株的死亡,都可以正常抽穗、结实,M3代验证发现该突变表型均可以稳定遗传,后代无分离现象。旗叶卷曲突变表现为部分旗叶明显变小卷曲(图2G),不利于光合作用的进行。无旗叶突变表现为植株部分分蘖的旗叶缺失(图2H),但能够正常抽穗、结实,该性状并不能稳定遗传。宽窄叶突变表现为叶片变宽或变窄:苏啤6号野生型植株的旗叶平均叶宽为1.37 cm,宽叶突变体旗叶平均叶宽为2.75 cm(图2I);细叶突变体旗叶平均叶宽仅为0.72 cm(图2J),M3代验证宽窄叶突变表型均可以稳定遗传,后代无分离。无叶耳突变材料无叶耳和叶鞘,叶片竖直向上生长,株型紧凑,是改良大麦株型的理想材料,通过对后代遗传分析表明,该性状能稳定遗传,为单基因隐性突变(图2K)。

A:叶黄条;B:叶白条;C:红绿叶;D:白叶耳;E和F:叶病斑;G:旗叶卷曲;H:无旗叶;I:宽叶;J:细叶;K:无叶耳;L:野生型。
2.3 茎秆突变
M2代株系中,有33个突变性状与茎秆相关,突变频率为1.38%。茎秆相关突变主要包括无蜡质、多节多叶、株高异常、分蘖异常、茎秆异常及茎秆弯曲6种变异类型。野生型整个植株均覆盖有白色的蜡质,无蜡质突变单株表现为叶片、茎秆或穗部蜡质缺失(图3A,B)。多节多叶突变体从拔节期至抽穗期,节间和叶片数目明显多于野生型,苏啤6号野生型主茎叶片的数目大约为10个,而该突变体的主茎叶数目可以达到15个,并且每个节间明显变短,穗子变小,该性状可以稳定遗传(图3C)。株高突变表现为植株变高和植株矮化,苏啤6号植株的高度为76 cm,高秆突变单株有85 cm左右,矮化突变单株大约有20 cm(图3D)。分蘖突变表现为分蘖数减少或增加,部分单株仅有一个分蘖,部分单株有多个分蘖,分蘖数平均达到15个以上(图3E),并且该性状可以稳定遗传。茎秆异常表现为一个分蘖有多个茎秆,穗子不能正常抽出,该性状不能稳定遗传(图3F)。茎秆弯曲突变主要表现为抽穗后穗子茎部扭曲(图3G),其他性状与野生型没有明显差异。

A和B:无蜡质;C:多节多叶;D:矮化;E:多分蘖;F:茎秆异常;G:茎秆弯曲;H:野生型。
2.4 穗部突变
M2代株系中发现了22个与穗子相关的突变体,突变频率为0.92%(表1),主要包括棱型、异常穗、球穗、浅黄穗、白颖壳、双穗和穗长变异7种类型。野生型苏啤6号为二棱皮大麦品种,突变材料中发现了六棱的突变株,并且能够稳定遗传(图4A)。异常穗突变棱型介于二棱与六棱之间,一个穗子中间部位有多排杂乱无章的籽粒,成熟后,还有部分籽粒无皮,为裸大麦(图4B,C)。球穗突变表现为穗子变短、变圆,类似于球型,结实率降低(图4D)。浅黄穗突变表现为灌浆期穗子的颜色为浅黄色,其他性状与野生型相比,没有明显差异(图4E)。白颖壳突变出现在抽穗至灌浆期,突变株颖壳的颜色为白色,穗子缺失叶绿体,后代验证发现,该性状可以稳定遗传,为单基因隐性突变(图4F)。双穗突变表现为茎秆上面有两个穗子,M3代验证表明该性状不能稳定遗传(图4G)。穗子长度突变表现为穗子变长或变短:野生型穗子长度为7.2 cm,长穗突变体平均长度为9.9 cm,穗粒数也明显增多(图4H);短穗突变体籽粒变小,排列紧密,平均长度仅为4.35 cm(图4I)。
2.5 生育期突变
经EMS诱变后,部分大麦的抽穗期与成熟期均发生了较大的变化。M2代中有6个早抽穗株系,4个晚抽穗株系,突变频率分别为0.25%和0.17%。M2代中有5个早熟株系,成熟期平均提前了5~10 d;有6个晚熟株系,成熟期平均推迟了4~7 d。
2.6 育性突变
M2代中共有6个株系的育性降低,其中有2个为完全不育,另外4个育性为10%~40%;开花期不育材料的颖壳膨大、张开,部分接受外部花粉,可以结实。有1个不育材料颖壳中的三枚雄蕊全部突变成了雌蕊(图4J)。

A:异常棱型;B和C:异常穗;D:球穗;E:浅黄穗;F:白颖壳;G:双穗;H:长穗;I:短穗;J:雄性不育;K:野生型。
2.7 纯合突变体的农艺性状分析
对M3代稳定的突变体株系进行主要农艺性状调查,结果表明(表2),突变群体中,在抽穗期、株高、穗长、单穗穗粒数、总蛋白含量和总淀粉含量方面均产生了丰富的变异,抽穗期为208~ 227 d,株高为21.67~85.54 cm,穗长为4.53~10.9 cm,单穗穗粒数为14.25~72.52,总蛋白含量为12.42%~16.78%,总淀粉含量为 39.46%~ 53.42%;其中单穗穗粒数与穗长的变异系数较大,分别为31.02和13.81。通过EMS诱变处理,在主要农艺性状方面均产生了不同于野生型的突变体,为以后基础理论研究及育种工作提供了基础材料。
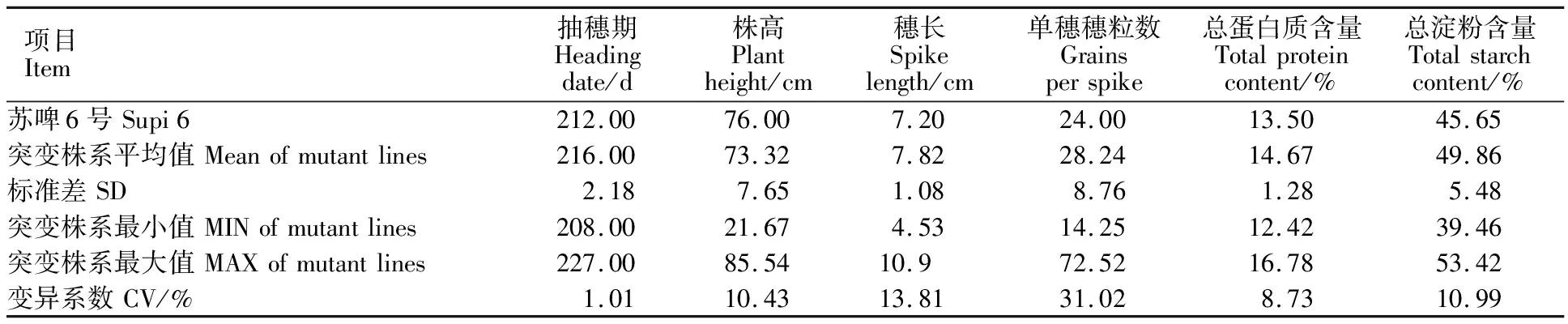
表2 M3代纯合突变体株系的农艺性状表现
3 讨 论
EMS诱变可以引起表型变异,目前,该技术已经应用于拟南芥、水稻、小麦、糜子、燕麦和大豆等植物中,创制出了许多有价值的种质资源,为作物重要农艺性状和品质特性等相关基因的克隆提供了材料基础[3-4,8-10]。然而目前关于大麦功能基因组学的研究还处于起步阶段,如Qin等[14]通过图位克隆的方法,成功克隆了大麦品种鄂大麦6号阶段性白化基因whs18。张晓勤等[15]对大麦浙农大3号进行EMS诱变,得到1 300个M2株系,筛选出各种形态突变株系97个,突变率为 7.46%。本研究以优质的啤酒大麦品种苏啤6号为材料,利用EMS诱变的方法构建了大麦突变体库,获得2 400个M2代材料。对M2株系进行全生育期考察,筛选到152个突变株系,并发现了多个未见报道的突变类型,如大麦多叶、三枚雄蕊雌化等。该突变体库的创建丰富了大麦突变体库,为我国大麦功能基因组学研究和新品种选育提供了新的实验材料。
叶色突变体是研究植物光合作用及光形态建成等代谢途径的重要材料,在杂交育种中还可以作为重要的形态标记。水稻上已报道了190份叶色突变体,并克隆到了30个相关调控基因[16]。大麦上也有相关叶色突变体的报道,但克隆的相关调控基因还较少。秦丹丹等[17]通过EMS诱导鄂大麦934,筛选到一个整个生育期叶色都为浅黄色的突变体,并将叶色突变基因初步定位到3H的12.7 cM区间内;Wang等[18]将受高温诱导的大麦黄化基因vvy初步定位到4H的长臂上;本课题组前期发现了一个大麦白色颖壳突变体,并通过转录组学分析初步了解了其形成的分子机理[19]。本研究所构建的苏啤6号突变体库中也发现多个叶色突变体,包括叶片黄化、白化、白条纹、黄条纹、红色条纹、白叶耳和白颖壳等,今后将通过构建遗传群体,进一步研究这些突变的形成机理。
表皮蜡质是覆盖于陆生植物外部的一层有机化合物,对于植物适应各种逆境环境(干旱、病虫害等)具有重要作用。在拟南芥、水稻和玉米等植物中已筛选到大量蜡质缺失突变体,目前在拟南芥中已克隆出几十个与蜡质合成和调控相关的基因,为表皮蜡质合成途径及调控网络的解析奠定了基础[20]。在麦类作物中,通过图位克隆得到的蜡质相关基因还较少,Li等[21]通过诱变得到了大麦叶片蜡质缺失突变体cer-ym,表现出对干旱敏感,并利用F2群体将该基因初步定位到4H染色体上;Zhou等[22]将大麦叶片和穗部蜡质缺失基因cer-b2定位到3H染色体1.3 cM区间内。本研究共鉴定出各种蜡质缺失突变体15份,并对穗部蜡质缺失突变体和野生型进行了形态结构和转录组学分析,从细胞和分子水平上初步解释了穗部蜡质缺失性状形成的机理[23]。
叶夹角是决定作物理想株型的重要因素之一,叶夹角大小与叶耳的结构密切相关。本研究发现了2个无叶耳突变系,叶片与主茎间夹角明显减小,叶片呈直立向上的形态,个体间对光能吸收的竞争减少,适合密植,有增产的潜力。在水稻和玉米上已有无叶耳突变体的相关报道[24-25],并对相关基因进行了精细定位,可能与SBP转录因子有关,大麦上还未见相关基因被克隆报道。本研究还发现了1个雄蕊雌化突变系,颖壳内的三枚雄蕊全部突变成了雌蕊,该突变体的发现对研究麦类作物花器官发育的分子机理以及不育系杂交育种研究都具有重要意义。
本研究对EMS诱变产生的突变株系的主要农艺性状进行了进一步的分析,发现了一批稳定的优良株系,比如矮秆、多叶(用于大麦青贮)、早熟和低蛋白等性状优良的突变株,这些优异种质资源可以直接应用到大麦育种中。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
核农学报(2022年6期)2022-05-20
辽宁农业科学(2021年6期)2021-12-02
蔬菜(2021年7期)2021-11-27
江西农业学报(2021年8期)2021-09-08
中国蔬菜(2021年3期)2021-03-29
科学与财富(2021年31期)2021-03-08
上海师范大学学报·自然科学版(2020年5期)2020-12-18
天津农林科技(2020年1期)2020-04-08
中国农业科技导报(2020年3期)2020-03-15
