
PACAP38调控TRPC6表达对糖尿病视网膜病变大鼠的保护作用
2020-07-24 02:08皮百木宋玫侠
眼科新进展 2020年7期
皮百木 宋玫侠
糖尿病视网膜病变(DR)是糖尿病中最常见的微血管并发症之一,也是导致患者视力丧失的主要原因[1],迄今为止,DR的发病机制尚未完全阐明。近年来有研究发现,垂体腺苷酸环化酶激活肽-38(pituitary adenylate cyclase-activating peptide-38,PACAP38)作为一种具有潜在价值的新型神经肽,对DR期间发生的一些病理生理变化具有保护作用[2-3]。PACAP38可改善谷氨酸诱导的神经毒性形态,抵消了由于近紫外线UVA辐射损伤引起的感光细胞退化,以及在高血糖早期通过调节炎性/低氧性事件保护血-视网膜屏障[4-5]。有研究报道,瞬时受体电位通道6(transient receptor potential channel 6,TRPC6)的过表达在DR发病过程中对大鼠视网膜神经节细胞(retinal ganglion cell,RGC)具有损伤作用[6]。在哺乳动物TRP超家族中,经典受体TRPC6在多种生物中表达,并表现出多种生理功能[7]。研究发现,TRPC6-/-小鼠免受高血糖诱发的血管退化,并对STZ诱导的视网膜层具有保护作用[7]。最新研究发现,一些肽的生物活性是通过瞬时受体电位(transient receptor potential,TRP)介导实现的,如神经元的抗凋亡作用、促进肺癌细胞增殖、角膜上皮和内皮的伤口愈合[8-9]。但有关PACAP38与TRPC6保护DR的关系目前尚不清楚。因此,本研究通过建立大鼠DR模型以及体外视神经RGC-5细胞模型,初步探讨PACAP38在DR过程中的视网膜保护作用是否由TRPC6介导实现的。
1 材料与方法
1.1 材料链脲佐菌素(STZ)、PACAP38(美国Sigma Aldrich公司),MicronⅣ视网膜影像系统(美国Phoenix Research Lab),GenTeal凝胶(美国Novartis公司),光学显微镜(德国Zeiss Axioplan公司),RIPA裂解液(瑞士Roche Diagnostics公司),BCA蛋白质检测试剂盒(美国Pierce公司),山羊抗TRPC6、鼠抗β-actin一抗、辣根过氧化物酶标记的二抗IgG(美国Santa Cruz Biotechnology公司),MTT试剂盒与超敏ECL试剂盒(上海碧云天生物技术有限公司),胎牛血清FBS(美国HyClone公司),DMEM/F12完全培养基(美国Thermo Scientific公司)等。
1.2 方法
1.2.1 动物饲养6~8周龄SPF级健康雄性SD大鼠30只,体质量100~120 g,购自上海SIPPR/BK实验动物有限公司(动物合格证编号:SCXK2008-0016)。将其饲养于温度为25~28 ℃、相对湿度为70%~85%、明/暗周期为12 h的标准化动物实验室,自由进食与饮水。
1.2.2 动物模型的构建及分组将SD大鼠分为空白对照组及DR模型组(STZ组),每组15只。大鼠糖尿病模型是通过单次腹腔注射STZ构建的。具体操作:注射STZ前大鼠禁食12 h,取适量STZ溶于柠檬酸钠缓冲液(pH 4.5),按照60 mg·kg-1体质量对大鼠进行一次性腹腔注射。对照组大鼠注射相同体积的柠檬酸钠缓冲液。药物注射48 h后检测大鼠血糖,其血糖浓度超过16.7 mmol·L-1、尿量及饮水明显增多即视为建模成功。STZ组根据不同的干预措施,再进行分组。注射STZ后2周,一次性向大鼠一眼玻璃体内注射含100 μmol·L-1PACAP38 PBS溶液4 μL,为STZ+PACAP38组;另一眼以同样的方法注射相同体积的PBS作为对照,为STZ对照组。注射PACAP38 6周后,腹腔注射致死量的戊巴比妥钠使各组大鼠安乐死,立即收集大鼠视网膜组织,一部分置于冰冻缓冲液中匀浆,进行Western blot检测,另一部分置于40 g·L-1多聚甲醛中固定,用于后续免疫组织化学分析。
1.2.3 光学相干断层扫描应用MicronⅣ视网膜影像系统分别在注射前及PACAP38注射后3周、6周对各组大鼠进行光学相干断层扫描(OCT)以确定大鼠视网膜厚度。具体操作:将大鼠麻醉,用10 g·L-1托品酰胺滴眼散大瞳孔。用GenTeal凝胶润滑角膜,并将Micron目镜置于凝胶与眼睛直接接触的位置。使用全扫描设置以平均10帧·s-1扫描速度捕获OCT图像。每只眼睛在相对于视神经上、下和鼻、颞方向各拍摄3张图像。
1.2.4 免疫组织化学分析取固定于40 g·L-1多聚甲醛的视网膜组织,用石蜡包埋,常规切片(厚3 μm),脱蜡、水化,并将切片置于体积分数0.3% H2O2/甲醇溶液中孵育30 min以淬灭内源性过氧化物酶活性,随后用PBS漂洗20 min。将切片用PBS中的体积分数1%牛血清白蛋白(BSA)处理1 h以减少非特异性染色,4 ℃下与山羊抗TRPC6孵育过夜(1100)。PBS漂洗后将切片于室温下辣根过氧化物酶标记的二抗(11000)孵育10 min,PBS冲洗后与二氨基联苯胺孵育5 min,苏木精复染。取染色的部分切片进行脱水、透化,最后用中性树脂封片,于光学显微镜下观察。
1.2.5 Western blot检测各组大鼠视网膜组织中TRPC6的表达收集各组大鼠视网膜组织,取适量置于含蛋白酶抑制剂的RIPA裂解液中冰浴进行裂解,低温离心收集上清液。采用BCA蛋白质检测试剂盒测定各组大鼠视网膜组织中蛋白质浓度。调整各组蛋白样本35 μg进行上样,采用100 g·L-1SDS-PAGE分离胶进行电泳,随后采用湿法转膜将蛋白质转移于PVDF膜。将膜用50 g·L-1脱脂牛奶在室温下封闭2 h,随后将PVDF膜分别置于含山羊抗TRPC6(1800)和兔抗β-actin(11000)一抗稀释液中4 ℃下孵育过夜。第2天用TBST漂洗3次,每次10 min,室温下用辣根过氧化物酶标记的二抗稀释液中(15000)孵育1 h。TBST漂洗同前。采用超敏ECL试剂盒(上海碧云天生物公司)显影曝光,采用自动图像分析系统对蛋白质条带进行扫描和光密度分析,以β-actin作为内参,进行统计分析。实验独立重复3次。
1.3 RGC-5细胞培养人RGC-5细胞购自美国ATCC公司,将其置于含有体积分数10%胎牛血清、100×103U·L-1青霉素和100×103U·L-1链霉素的DMEM/F12完全培养基中,置于37 ℃、含体积分数5%CO2培养箱中培养。每1~2 d换液1次,每周按照12的比例传代培养1次,取传至3代对数生长期的细胞用于后续实验。
1.3.1 RGC-5细胞分组及处理取对数生长期的RGC-5细胞,用胰蛋白酶消化后,调整细胞密度为500×103个·mL-1接种于HTS Transwell渗透板上培养7 d,隔天换液。随后,将Transwell渗透板中一半的细胞用含5.5 mmol·L-1D-葡萄糖DMEM/F12完全培养基继续培养7 d,作为对照(NG)组;另一半细胞用含25 mmol·L-1D-葡萄糖DMEM/F12培养液继续培养7 d,作为高葡萄糖(HG)组。
收集部分HG组细胞,调整细胞浓度为5000个·mL-1接种于96孔板中,用含25 mmol·L-1D-葡萄糖DMEM/F12培养液培养过夜。第2天弃去原有培养基,用单独含100 μmol·L-1低氧模拟剂去铁胺(desferrioxamine,DFO)或分别联合含100 nmol·L-1PACAP38、100 nmol·L-1TRPC6通道激动剂(OAG)、10 μmol·L-1PAC1受体拮抗剂(PACAP6-38)、20 nmol·L-1TRPC6通道阻滞剂(SKF96365)的高糖DMEM/F12培养液培养24 h。NG组细胞以相同的密度接种于96孔板中,用含5.5 mmol·L-1D-葡萄糖DMEM/F12完全培养基于37 ℃、含体积分数5%CO2培养箱中继续培养。实验结束后,用MTT法检测各组细胞的活性。
1.3.2 MTT法检测各组RGC-5细胞的活力每孔按照20 μL MTT溶液(0.5 g·L-1MTT溶于PBS)加入96孔板中,于37 ℃、含体积分数5%CO2培养箱中孵育4 h。将96孔板低速离心后,弃去原培养基,加入150 μL二甲基亚砜室温下孵育10 min。采用FL-600酶标仪于570 nm波长处检测各组的吸光度(A)值,每组5个复孔,实验独立重复3次。细胞活力=A实验组/A对照组×100%。
1.4 统计学方法数据采用SPSS 17.0统计学软件进行分析。计量数据采用均数±标准差表示,组间比较采用单因素方差分析(ANOVA)进行检验。检验水准:α=0.05。
2 结果
2.1 各组大鼠体质量和血糖变化与空白对照组大鼠相比,STZ组大鼠注射STZ后2周以及注射PACAP38 1周后、3周后、6周后体质量均显著降低(均为P<0.05),且血糖水平均明显升高(均为P<0.05)。见表1。

表1 各组大鼠体质量和血糖浓度变化
2.2 PACAP38对DR大鼠视网膜厚度的影响OCT扫描结果显示:造模前各组大鼠视网膜厚度相似。注射PACAP38 3周后,STZ对照组和STZ+PACAP38组的视网膜厚度显著高于空白对照组(P<0.05),且STZ对照组的视网膜厚度显著高于STZ+PACAP38组(P<0.05)。玻璃体内注射PACAP38 6周后,STZ对照组的视网膜厚度显著高于空白对照组(P<0.05),而STZ+PACAP38组视网膜厚度较空白对照组略高,其差异无统计学意义(P>0.05)。见表2。
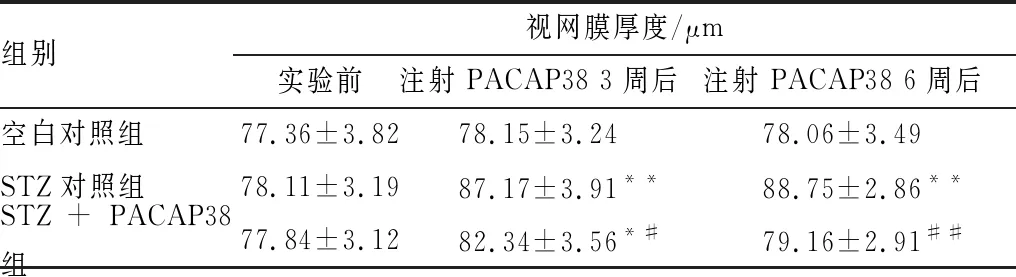
表2 PACAP38对DR大鼠视网膜厚度的影响
2.3 PACAP38对DR大鼠视网膜组织中TRPC6表达的影响免疫组织化学检测结果显示:空白对照组大鼠视网膜组织中发现较弱TRPC6的阳性表达(呈黄褐色斑点,图1A)。与空白对照组相比,STZ对照组大鼠视网膜神经节细胞层(ganglion cell layer,GCL)、内丛状层(inner plexiform layer,IPL)、内核层(inner kernel layer,INL)和外丛状层(outer plexiform layer,OPL)中均发现TRPC6阳性表达较显著(图1B);STZ+PACAP38组大鼠GCL、IPL、INL、OPL中TRPC6阳性表达较STZ对照组大鼠明显减少(图1C)。Western blot检测结果显示:与空白对照组相比,STZ对照组和STZ+PACAP38组大鼠视网膜组织中TRPC6蛋白表达均显著增加(均为P<0.05),且STZ对照组大鼠视网膜TRPC6蛋白表达显著高于STZ+PACAP38组(P<0.01)。见图2、表3。

表3 PACAP38对DR大鼠视网膜组织中TRPC6蛋白相对表达的影响
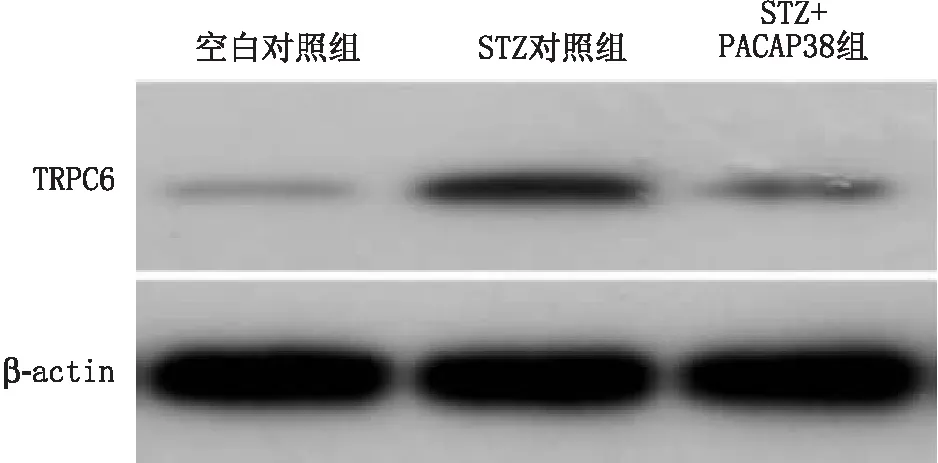
图2 Western blot检测PACAP38对DR大鼠视网膜组织中TRPC6蛋白表达的影响

图1 PACAP38对DR大鼠视网膜组织中TRPC6表达的影响 A:空白对照组;B:STZ对照组;C:STZ+PACAP38组
2.4 PACAP38对RGC-5细胞活力的影响MTT检测结果显示:与NG组相比,HG+DFO组的细胞活性显著降低(P<0.001)。与HG + DFO组相比,PACAP38或SKF96365处理可增强HG+DFO培养的细胞活力(均为P<0.01)。与HG+DFO组相比,PAC1受体拮抗剂(PACAP6-38)或TRPC6通道激动剂(OAG)处理24 h后RGC-5细胞活力明显降低(均为P<0.05)。与HG+DFO+OAG组相比,HG+DFO+PACAP38+OAG组RGC-5细胞活力显著增加(P<0.01);相反,与HG+DFO+SKF96365组相比,HG+DFO+SKF96365+ PACAP638组细胞活力未见增加。见表4。
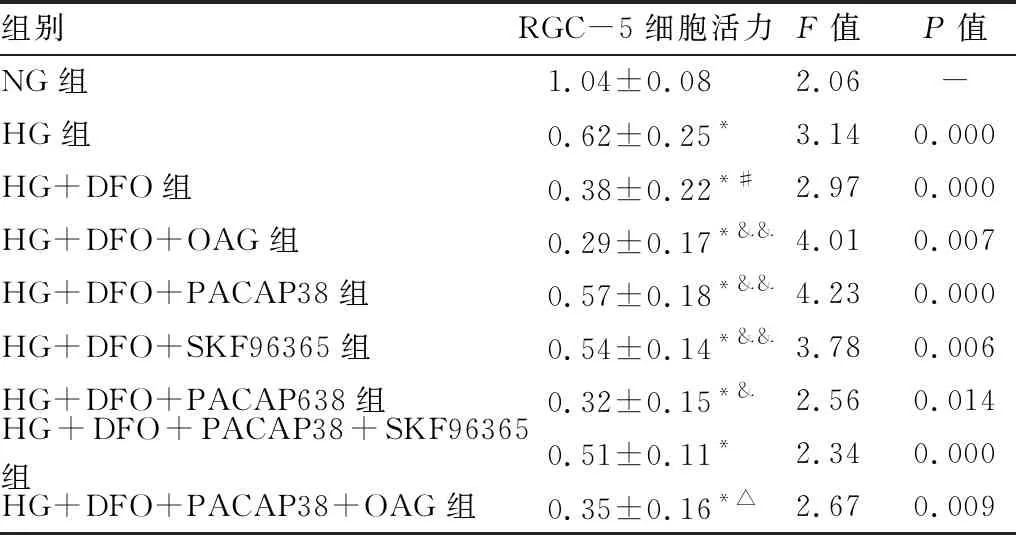
表4 PACAP38对暴露于高血糖/低氧的RGC-5细胞活力的影响
3 讨论
STZ大鼠是目前公认的经典DR模型,可模拟人体DR的大致病理变化[10]。本研究中腹腔一次性注射STZ诱导DR大鼠动物模型,与空白对照组相比,经腹腔注射STZ 48 h后,STZ大鼠血糖浓度大于16.7 mmol·L-1,尿量及饮水明显增多,且随着注射时间延长,其血糖及较空白对照组显著升高,体质量较对照组大鼠明显降低(P<0.05)。这提示DR动物模型制作成功,以确保后续实验结果顺利进行。
近年来有研究证实,内源性PACAP的减少导致N-甲基-D-天冬氨酸引起的神经元损伤增加,但外源性PACAP的加入具有代偿作用。由于PACAP和垂体腺苷酸环化酶1激活受体(pituitary adenylate cyclase 1 activated receptor,PAC1-R)均在RGCs中表达,玻璃体内注射PACAP可通过自分泌或旁分泌系统发挥视网膜神经保护作用[2]。有学者发现,PACAP可促使视网膜细胞分裂和诱导神经节细胞分化,有助于视网膜色素上皮的发展[11]。此外,还有学者发现PACAP38能够调节VEGF的释放,可在高糖/低氧双重损伤后恢复外部血-视网膜屏障完整性[12]。Seki等[13]研究证明,玻璃体内注射100 μmol·L-1的PACAP38具有良好的神经保护作用。因此,本研究采用与Seki等[13]报道相似的干预措施,发现PACAP38有效逆转了糖尿病大鼠视网膜增厚,且PACAP38处理可提高暴露于高血糖/低氧环境中RGC-5细胞活力。这说明PACAP38对DR期间发生的一些病理生理变化具有保护作用。
近年来还有研究表明,PACAP的某些生物效应是通过TRPC介导的[8-9]。自果蝇光转导突变中首次发现TRPC以来,越来越多的证据表明TRPC在神经退行性疾病中扮演着重要作用。汪书越[14]对视网膜TRPC的某些功能进行了初步研究,结果表明,TRPC可能介导RGC的基础Ca2+流入,激活TRPC导致视网膜Ca2+超载变性。最近有研究发现,抑制内源性TRPC6可能有助于改善视网膜缺血,发挥神经保护作用[15]。TRPC6的活化在视网膜损伤产生的反应性星形胶质细胞中大量表达,同时触发了表征反应性胶质增生的其他几种基因的上调或下调[16]。本研究免疫组织化学结果发现,STZ对照组大鼠视网膜的GCL、IPL、INL和OPL中均发现TRPC6阳性表达,这与Wang等[15]报道结果相一致,表明代谢异常的糖尿病微环境会触发星形胶质细胞的活化,从而产生多种促炎症细胞因子,导致TRPC6活化[17]。此外本研究还发现,PACAP38的干预可逆转由STZ诱导DR大鼠视网膜组织中TRPC6阳性表达,这提示PACAP38可通过调控TRPC6通路活化保护DR损害的视网膜屏障。
为阐明PACAP38与TRPC6通路之间的关系对DR生理病理变化的影响,本研究通过视网膜屏障的体外细胞模型作初步分析。将RGC-5细胞暴露于高糖/低氧环境中,来模拟DR的体内微环境。由MTT检测结果可知:与HG组及HG+DFO组相比,PACAP38处理可明显提高HG+DFO培养的RGC-5细胞活力(P<0.01);与HG+DFO+PACAP38组相比,采用PACAP38和OAG联合处理暴露于高糖/低氧环境中RGC-5细胞可显著降低细胞活性(P<0.05)。有研究发现TRPC6活化涉及多种细胞生理过程,包括细胞增殖、运动性和黏附性[18-19]。本研究结果提示PACAP38与TRPC6之间的相互作用与视神经细胞存活有关,且外源性PACAP38干预可提高高糖/低氧环境中RGC-5细胞的活性。
有关PACAP38与TRPC6的关联性在DR中的作用目前还尚未见报道。本研究采用STZ腹腔注射诱导大鼠DR模型,玻璃体内注射给予PACAP38干预,发现PACAP38可减轻STZ诱导的DR损伤,且通过抑制TRPC6的过表达有效逆转了糖尿病大鼠视网膜增厚,提高了暴露于高血糖/低氧环境中RGC-5细胞活力。这提示PACAP38与TRPC6参与DR的进程,具体的分子机制仍需进一步研究。
猜你喜欢
中日友好医院学报(2022年4期)2022-10-15
天津医科大学学报(2021年3期)2021-07-21
看世界·学术下半月(2020年7期)2020-09-10
江西医药(2020年4期)2020-04-28
中国特种设备安全(2019年4期)2019-05-20
广东农业科学(2017年10期)2018-01-25
中成药(2017年9期)2017-12-19
中国医学创新(2017年22期)2017-11-15
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
