
棉花基因组中赤霉素氧化酶基因的鉴定与分析
2020-07-23 16:37赵亮狄佳春陈旭升
江苏农业学报 2020年3期
赵亮 狄佳春 陈旭升

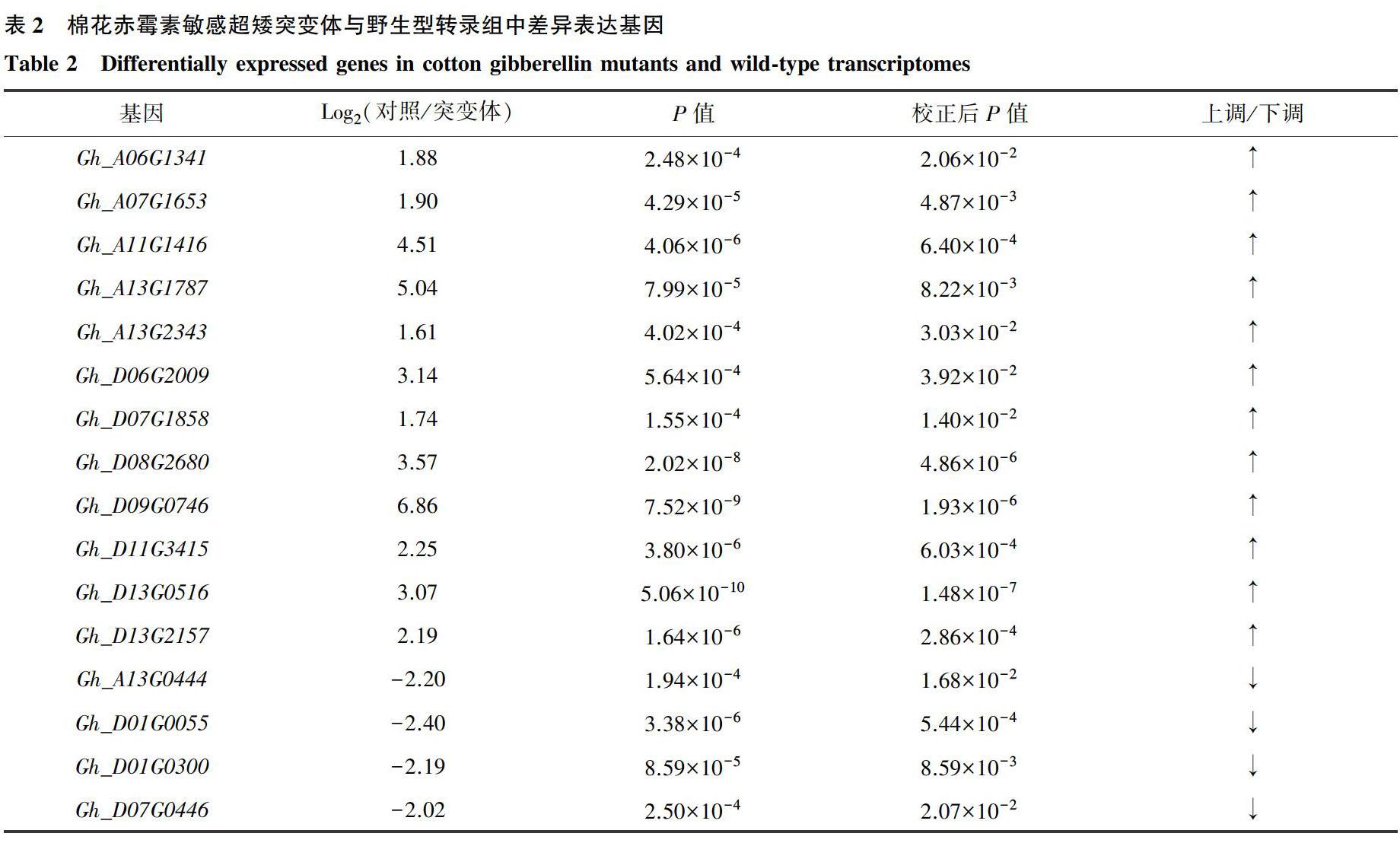

摘要:根据赤霉素氧化酶氨基酸序列的保守结构域和生物学信息,对棉花2个二倍体野生种(A基因组的亚洲棉和D基因组的雷蒙德氏棉)和2个异源四倍体遺传标准系(陆地棉TM-1和海岛棉H7124)进行了全基因组的查找。结果表明,在2个二倍体野生种的A和D基因组中分别预测到219和188个该酶类基因家族成员;在异源四倍体陆地棉TM-1和H7124中分别预测到了310和428个该酶类基因家族成员。通过分析陆地棉中预测到的310个该酶类基因家族成员在赤霉素敏感超矮突变体和野生型转录组中表达差异,发现16个基因具有显著性差异。利用这16个基因编码的蛋白质氨基酸序列与报道的拟南芥赤霉素氧化酶氨基酸序列进行进化分析,结果表明Gh_D06G2009属于GA3ox氧化酶,Gh_D01G0300和Gh_D09G0746为 GA2ox氧化酶,Gh_A06G1341、Gh_A07G1653、Gh_A11G1416、Gh_A13G0444、Gh_A13G1787、Gh_A13G2343、Gh_D01G0055、Gh_D07G0446、Gh_D07G1858、Gh_D08G2680、Gh_D11G3415、Gh_D13G0516、Gh_D13G2157为GA20ox氧化酶。
关键词:棉花;赤霉素;赤霉素氧化酶;基因家族
中图分类号:S562.01文献标识码:A文章编号:1000-4440(2020)03-0553-08
Identification and analysis of gibberellin oxidase gene in cotton genome
ZHAO Liang,DI Jia-chun,CHEN Xu-sheng
(Institute of Industrial Crops, Jiangsu Academy of Agricultural Sciences/ Key Laboratory of Cotton and Canola Research at the Lower Reach of the Yangtze River Plain, Ministry of Agriculture and Rural Affairs, Nanjing 210014, China)
Abstract:Based on the conserved domain of gibberellin oxidase amino acid sequence and bioinformatics, genome-wide searches were performed on two diploid wild cotton species (Gossypium arboreum with A genome and G. Raimondii with D genome) and two allotetraploid genetic standard lines (upland cotton TM-1 and island cotton H7124). In the two diploid wild species, 219 and 188 members of the enzyme family were predicted in the A and D genomes, respectively. In addition, 310 and 428 members of the enzyme family were predicted in allotetraploid upland cotton TM-1 and H7124, respectively. Subsequent analysis of 310 members predicted by us in upland cotton revealed significant differences in 16 genes between gibberellin mutants and wild-type transcriptomes. The amino acid sequence of the protein encoded by these 16 genes and the amino acid sequence of gibberellin oxidase in Arabidopsis thaliana were analyzed, it was found that Gh_D06G2009 could belong to GA3ox oxidase. Gh_D01G0300 and Gh_D09G0746 were GA2ox oxidase. However, Gh_A06G1341, Gh_A07G1653, Gh_A11G1416, Gh_A13G0444, Gh_A13G1787, Gh_A13G2343, Gh_D01G0055, Gh_D07G0446, Gh_D07G1858, Gh_D08G2680, Gh_D11G3415, Gh_D13G0516, Gh_D13G2157 were GA20ox oxidase.
Key words:cotton;gibberellic acid;gibberellin oxidase;gene family
赤霉素(GAs)是一种重要的植物类激素,参与植物生长与发育等多个生物学进程。在高等植物和真菌中,总计有136种赤霉素被鉴定[1],但是只有一少部分具有生物活性,例如GA1、GA3、GA4和GA7[2]。在高等植物中,赤霉素的生物合成总共涉及到8个关键酶,分别为CPS、KS、KO、KAO、GA20ox、GA13ox、GA2ox和GA3ox,生物合成划分为3个阶段:在质体中,牻牛儿牻牛儿二磷酸(GGDP)合成贝壳杉烯(ent-kaurene);通过细胞色素P450单加氧酶,贝壳杉烯(ent-kaurene)转变为GA12;在细胞质中,通过赤霉素氧化酶的作用形成C20-GAs和C19-GAs。植物中GAs最初来自于GGPP,它存在于质体上,是最常见的二萜类C20前体[3]。在模式植物拟南芥中发现的赤霉素敏感型突变体较多,每个合成途径基因都有相应的突变体:GA1对应CPS,GA2对应KS,GA3对应KO,GA4对应3βOX,GA5对应20OX。前3个突变体为极端矮化突变体,后两者属于矮化突变体,此外对这5个突变体连续施用GA3,均能使其恢复正常植株的高度。
利用預测的CDS序列,通过DnaSP V6软件对4个基因组中同源基因进行了同义替换和异义替换分析。分析结果表明,异源四倍体的陆地棉和海岛棉中At亚组与亚洲棉的A组之间的异义替换与同义替换的比值(Ka/Ks)都小于1,同样异源四倍体的Dt亚组与雷蒙德氏棉的D组之间的Ka/Ks值只有Gh_D11G3415基因的比值大于1,其余都小于1 (表3)。说明Ka/Ks值小于1的基因在进化过程中经历了自然界的纯化选择,而Ka/Ks值大于1的Gh_D11G3415基因在进化过程中经历了自然界的正向选择。
通过统计这16个基因在组织器官(根、茎和叶)中表达的相关性发现,总计有36对基因之间存在显著的正相关或负相关关系(r>0.9或r<-0.9),其中16对基因之间存在显著正相关关系,20对基因之间存在显著负相关关系(图3)。
3讨论
植物激素的研究一直是国内外植物科学研究的重点与热点[13-14]。农业生产上第一次“绿色革命”就是利用农作物本身的赤霉素合成和信号转导缺陷所产生的矮化植株来培育抗倒伏农作物新品种,从而大幅度提高了农作物的产量[15]。赤霉素氧化酶是重要的赤霉素生物合成和调控酶,该酶类是具有保守的结构域,同时基因家族成员的表达模式既有重叠,也存在区别[16]。本研究根据该酶类的保守结构域用生物信息学方法,对已经公开发表的4个棉种基因组数据中该酶类家族进行了鉴定。鉴定结果表明,在这4个棉种的基因组数据库中存在大量该酶类家族成员(亚洲棉219个,雷蒙德氏棉188个,陆地棉TM-1 310个,海岛棉H7124 428个)。由于通过生物信息学方法鉴定出的成员较多,因此我们对这些鉴定出的成员在本研究室发现的赤霉素敏感超矮突变体与野生型转录组数据库中进行了进一步的筛选,最终在陆地棉TM-1中得到16个赤霉素氧化酶家族成员。
系统发生树能够揭示生物进化过程的顺序,有助于了解生物进化的历史,从而解决生物学中的一些问题[17-18]。因此,我们对陆地棉TM-1中得到的16个可能参与赤霉素合成的氧化酶基因与拟南芥中的相关基因利用最大似然法进行进化树的构建。结果表明,在TM-1中,GA3ox类氧化酶基因有1个(Gh_D06G2009),GA2ox类氧化酶基因有2个(Gh_D01G0300和Gh_D09G0746),GA2ox类氧化酶基因有13个(Gh_A06G1341、Gh_A07G1653、Gh_A11G1416、Gh_A13G0444、Gh_A13G1787、Gh_A13G2343、Gh_D01G0055、Gh_D07G0446、Gh_D07G1858、Gh_D08G2680、Gh_D11G3415、Gh_D13G0516、Gh_D13G2157)。
未来,我们将对这些预测到的基因利用生物技术手段(例如病毒介导的转基因沉默技术等)进行反向遗传学验证,将这些基因敲除后通过对沉默植株的表型鉴定及赤霉素含量的测定来进一步确定参与赤霉素代谢的赤霉素氧化酶。
参考文献:
[1]MACMILLAN J. Occurrence of gibberellins in vascular plants, fungi, and bacteria[J]. J Plant Growth Regul, 2001,20(4):387-442.
[2]HEDDEN P, PHILLIPS A L. Gibberellin metabolism: new insights revealed by the genes[J]. Trends Plant Sci, 2000,5(12):523-530.
[3]UN T P. Gibberellin metabolism, perception and signaling pathways in Arabidopsis[J]. Arabidopsis Book, 2008,6:103.
[4]SCHOMBURG M F. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants[J]. Plant Cell, 2003,15(1):151-163.
[5]PLACKETT A R G, POWERS S J, FERNANDEZ-GARCIA N, et al. Analysis of the developmental roles of the Arabidopsis gibberellin 20-oxidases demonstrates that GA20ox1, -2, and -3 are the dominant paralogs[J]. Plant Cell, 2012,24(3):941-960.
[6]SONNHAMMER E L, EDDY S R, BIRNEY E, et al. Pfam: multiple sequence alignments and HMM-profiles of protein domains[J]. Nucleic Acids Res, 1998,26(1):320-322.
[7]KUMAR S, STECHER G, LI M, et al. MEGA X: Molecular evolutionary genetics analysis across computing platforms[J]. Mol Biol Evol, 2018,35(6):1547-1549.
[8]PRICE M N, DEHAL P S, ARKIN A P. FastTree 2-approximately maximum-likelihood trees for large alignments[J]. PLoS One, 2010,5(3):e9490.
[9]PRICE M N, DEHAL P S, ARKIN A P. FastTree: computing large minimum evolution trees with profiles instead of a distance matrix[J]. Mol Biol Evol, 2009,26(7):1641-1650.
[10]ROZAS J, FERRER-MATA A, SANCHEZ-DELBARRIO J C, et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets[J]. Mol Biol Evol, 2017,34(12):3299-3302.
[11]陈旭升,狄佳春,许乃银,等. 陆地棉超矮秆突变性状质量遗传规律分析[J]. 遗传, 2007, 29(4):471-474.
[12]LIANG Z, DI J, QI G, et al. The differentially expressed genes identification in dwarf mutant of Gossypium hirsutum by RNA-Seq approach[J]. Agri Gene, 2017,5:37-44.
[13]赵凤,王小乐,房伟民,等. 外源激素和温度对切花菊侧芽萌发与内源激素含量的影响[J].江苏农业学报,2018,34(1):145-151.
[14]李国龙,孙亚卿,邵世勤,等.甜菜幼苗叶片渗透调节系统及部分激素对干旱胁迫的响应[J].江苏农业科学,2018,46(7):80-84.
[15]谈心,马欣荣. 赤霉素生物合成途径及其相关研究进展[J]. 应用与环境生物学报, 2008,14(4):571-577.
[16]PHILLIPS A L, WARD D A, UKNES S, et al. Isolation and expression of three gibberellin 20-oxidase cDNA clones from Arabidopsis[J]. Plant Physiol, 1995,108(3):1049-1057.
[17]郭靜,王超,张宏彬,等. 系统发生树构建方法综述[J]. 计算机应用研究, 2013,30(3):647-655.
[18]李建伏,郭茂祖. 系统发生树构建技术综述[J]. 电子学报, 2006, 34(11):2047-2052.
(责任编辑:张震林)
猜你喜欢
农学学报(2022年4期)2022-07-14
少儿科学周刊·儿童版(2022年10期)2022-06-27
学苑创造·A版(2017年9期)2017-09-25
小溪流(画刊)(2017年1期)2017-03-16
现代农业科技(2016年20期)2016-12-20
现代农业科技(2016年20期)2016-12-20
天津农业科学(2015年12期)2015-12-03
山西果树(2014年3期)2014-07-15
小天使·一年级语数英综合(2014年7期)2014-06-26
果农之友(2009年8期)2009-09-11
