
长柄扁桃与蒙古扁桃幼苗抗旱性比较研究
2020-07-22 11:18:30张高如贾东波李奔奔韩瑞瑞李嘉丽段义忠柴乖强
榆林学院学报 2020年4期
张高如,贾东波,李奔奔,韩瑞瑞,李嘉丽,段义忠,柴乖强
(1.榆林市林业产业开发中心,陕西 榆林 719000;2. 榆林学院 生命科学学院,陕西 榆林 719000)
随着全球气候变暖,许多国家和地区都面临着不同程度的干旱、高温等非生物威胁,其中在干旱、半干旱自然环境植物生长过程中最依赖的是水分[1-2]。研究表明,高温和水分亏缺严重威胁着植物的正常生长和发育。为了应对逆境胁迫,植物会通过关闭气孔,维持体内细胞的渗透压进而继续进行细胞的代谢活动,如植物体内应答干旱胁迫的各种途径被激活如多酚氧化酶参与植物的干旱调节,从而减轻干旱对植物所造成的影响[3-4]。所以,从生理层面讲,增强植物抗旱性并提高水分利用率的一种行之有效的途径是减缓蒸腾速率。因此,筛选耐旱植物对干旱、半干旱地区植物资源的发展有深远意义。
长柄扁桃(Amygdalus pcdunculata Pall)和蒙古扁桃(Amygdalus mongolica (Maxim.) Ricker)因具有耐瘠薄、抗干旱、耐寒冷及适应性强等优点,是亚洲干旱地区防风固沙、水土保持的先锋树种,在荒山造林以及沙地治理等方面具有重要的功能,因而已经成为国内外研究的热点。但当前有关扁桃属的研究主要集中在沙漠化治理、药用成分的开发与利用、食用油的开发、地理分布与进化和功能基因的发掘等方面[5-7],对于长柄扁桃和蒙古扁桃抗旱的生理生化特征研究尚不清晰。本项目在前期研究发现长柄扁桃和蒙古扁桃抗干旱性都较强的基础上,拟采用生理生化指标测定、光合测定系统等,探究干旱胁迫对扁桃种子萌发和幼苗的损伤机理及扁桃的抗逆机制,总结两种扁桃应对干旱胁迫的生理特性响应及其变化规律,为加强干旱半干旱地区抗旱植物资源开发利用提供科学依据。
1材料与方法
1.1 植物材料
长柄扁桃与蒙古扁桃,采集于陕西省榆林市孟家湾(109°35' 50″E,38°41'42″N)的新鲜度过休眠期的种子,并于榆林学院生命科学学院地理与生态学试验室保存。长柄扁桃和蒙古扁桃种子经过夹破核壳和剥除核壳处理,并在体积浓度为10%的NaClO溶液中消毒处理10~15 min,后用蒸馏水冲洗4~5次,作为种子萌发及生理指标测定的材料。
1.2 试验方法
1.2.1 PEG-6000模拟干旱胁迫处理长柄扁桃种子
长柄扁桃种子的萌发试验参考谭春燕等[8]的方法,并做了部分修订。改进的方法如下:取15个直径15cm的大号玻璃培养皿,铺上定性滤纸。挑选750粒破壳且饱满的扁桃种子,放入培养皿中,每个培养皿放置50粒;分别用量筒量取15 mL质量分数为0%,5%,10%,15%和20%的PEG-6000溶液,每一个处理做3个平行重复,所有处理放置于光照培养箱中,培养温度统一为25℃。
发芽试验自第一天起每天定时(9点至12点)观察种子发芽状态(规定扁桃种子胚根露出大约2 mm视为发芽)并做统计记录,每天观察统计后,要更换培养皿中的PEG-6000溶液,同时,每3 d更换一次滤纸,以保持溶液渗透势恒定,连续进行25 d的跟踪观察,每天如实记录扁桃种子发芽状态并计算种子发芽率、发芽势以及发芽指数。计算参考何学青等[9]方法,其计算公式如下:
发芽率(%)= 已经发芽的种子数目/处理的种子数量×100;
发芽势(%)= 规定时间内发芽种子粒数/供试种子粒数×100;
种子活力指数VI=GI×S,S为处理第15天时的幼苗高度。
1.2.2干旱胁迫处理长柄扁桃幼苗
挑选饱满的长柄扁桃种子,机械夹破外表的种皮壳、用10%的次氯酸钠消毒处理10 min,无菌水冲洗5~6遍,用0.15%的双氧水催芽后播种至花盆中,每一花盆种6株,每个处理种植5盆,总共25盆。扁桃材料放置于玻璃科研温室中,温度控制在25℃左右,前期各个处理水、肥、光等管理保持统一,等到长柄扁桃幼苗长至20 cm高时(约两个月后),进行干旱胁迫处理。水分处理设置参考宋吉轩博士论文,并做了一定修改,处理主要分为:轻度干旱胁迫(Light stress,LS),中度干旱胁迫(Mild stress,MS)和重度干旱胁迫(Severe stress,SS)。
长柄扁桃苗期各个生理生化指标测定参考方法如下:光合速率的数据采集参考仪器说明书进行;植物叶片表皮气孔的观察参考Groll[10]的方法进行;幼苗叶片叶绿素含量测定采用80%的乙醇法;过氧化物酶(peroxidase,POD)、超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)和丙二醛(Malondialdehyde,MDA),测定的具体方法参照孙群和胡景江[11],每一个生理、生化指标均重复测定3~4次。
1.2.3干旱胁迫处理下扁桃幼苗叶片气孔的开张度检测及表皮蜡质的测定
为了检测各种胁迫条件下两种扁桃幼苗叶片气孔的开张程度,在三种干旱胁迫处理1~2小时后,用尖头镊子撕下扁桃叶片的下表皮层,放到载玻片上,在携带有照相机的光学显微镜下观察各个处理的气孔情况,显微镜的放大倍数为100×。
柄扁桃蜡质的提取方法参考柴乖强学位论文中的方法[12]。提取好的蜡质,在试验室抽风机内自然风干后,用千分之一天平称重。
1.3 数据处理与分析
试验中各数据均应用Excel 2016和DPS软件进行统计分析。采用单因素One-way ANOVA和Duncan进行差异显著性分析,并应用Sigmaplot 12.5进行作图分析。
2 结果与分析
2.1不同PEG-6000浓度对长柄扁桃和蒙古扁桃种子萌发期形态指标的影响
从表1可以看出,随着PEG6000浓度的逐渐增大,长柄扁桃和蒙古扁桃两个品种的发芽率和发芽势呈现出显著降低的趋势(P<0.01),当胁迫浓度达到20%时,两者均达到最低;发芽势、发芽指数、活力指数、幼苗高度、幼苗根长和幼苗鲜重随着PEG浓度增大,均呈现出先缓慢增高后显著降低的趋势,其中,PEG-6000浓度为5%时,各个指标均达到最大值,但与对照相比均没有显著差异,说明5%的PEG处理有助于长柄扁桃和蒙古扁桃的发芽生长,但当胁迫加大后,种子的受害程度逐渐加大,萌发率、发芽指数等极显著降低(P<0.01)。

表1 渗透胁迫对长柄扁桃种子萌发期形态指标的影响
注: 不同小写字母表示不同水平间差异显著(P<0.05);不同大写字母表示不同水平间差异极显著(P<0.01),下同。
从两个品种长柄扁桃和蒙古扁桃的对比来看,PEG处理后,两个品种的种子萌发指标表现出相似的规律。在重度胁迫下(20% PEG-6000),从发芽率、发芽指数、活力指数和幼苗根长等方面,蒙古扁桃的形态表现要比长柄扁桃较好。综合种子萌发过程中的各种形态指标,初步确定了蒙古扁桃的抗旱性能要比长柄扁桃强。
2.2 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗生理特性的影响
2.2.1不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗株高的影响
对培养约45 d的长柄扁桃和蒙古扁桃幼苗进行不同程度的干旱胁迫处理,处理结束后,分别对其进行株高的测量,结果表明(图 1),随着干旱胁迫的加重,长柄扁桃和蒙古扁桃的株高有所降低,但和对照相比没有显著性的差异(P>0.05)。两个品种比较后发现,长柄扁桃收到重度胁迫后,株高降低的更快,表明长柄扁桃对干旱胁迫表现比蒙古扁桃要敏感。
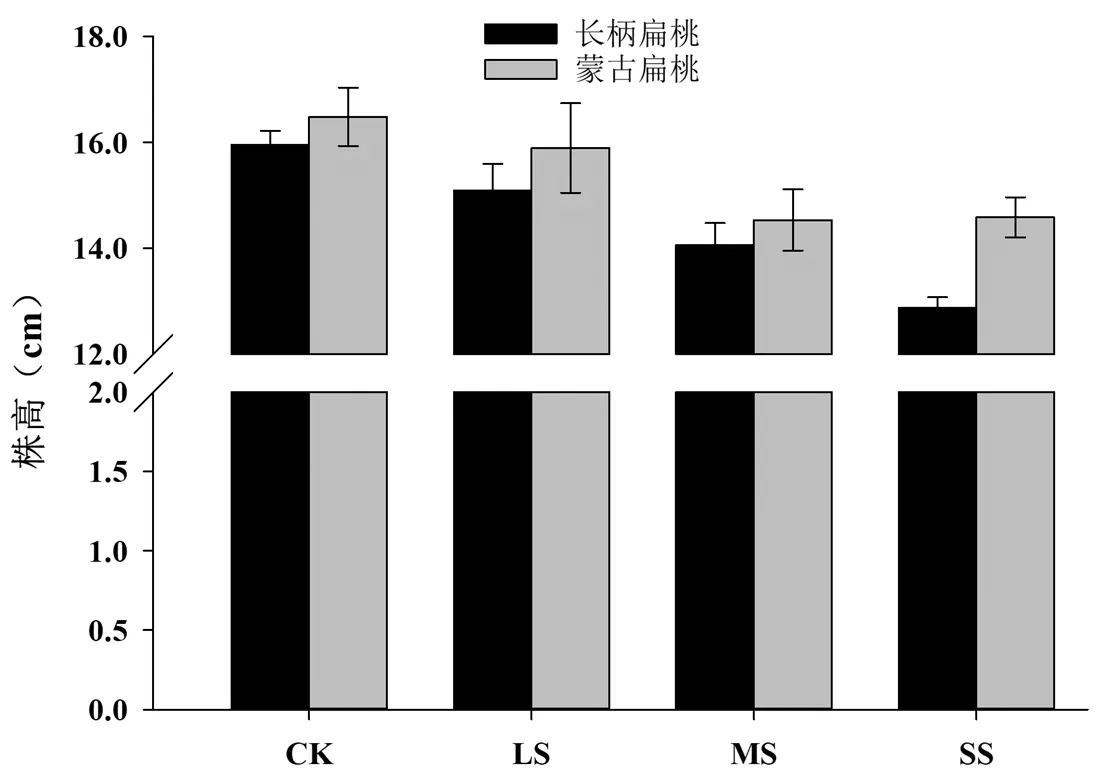
图1 干旱胁迫对长柄扁桃和蒙古扁桃幼苗株高的影响
2.2.2 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗净光合速率的影响
为了探明干旱胁迫对沙生植物扁桃属植物净光合速率的影响,我们利用光合仪测定了不同干旱胁迫处理下的扁桃光合速率,结果表明(图 2),随着干旱胁迫的逐渐加重,长柄扁桃和蒙古扁桃的净光合均呈现出先保持稳定变化后迅速降低的趋势。特别在中度胁迫和重度胁迫下,光合速率显著地低于对照(P<0.05)。
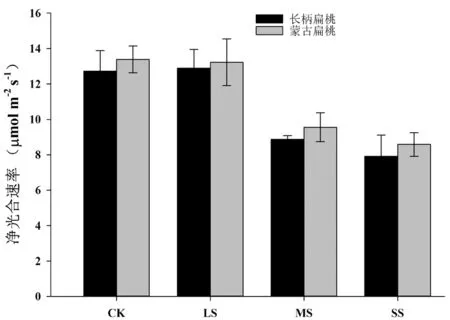
图2 干旱胁迫对长柄扁桃和蒙古扁桃幼苗叶片净光合速率的影响
2.2.3不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗气孔变化的影响
研究表明,植物在热、干旱、高盐等非生物逆境胁迫下,作出的响应之一是调控叶片表皮气孔的开张度来减少水分的散失。本试验通过对长柄扁桃和蒙古扁桃的干旱胁迫处理,发现在轻度干旱下,蒙古扁桃和长柄扁桃气孔没有关闭,当中度和重度干旱胁迫时,两种材料表皮气孔发生关闭或微张(图 3)。与长柄扁桃相比,蒙古扁桃在中度干旱胁迫下,仍有部分表皮气孔微微张开,依然能够发生蒸腾作用,说明蒙古扁桃对干旱胁迫的适应性要比长柄扁桃要强。

图3 干旱胁迫对长柄扁桃和蒙古扁桃幼苗叶片气孔的影响
2.2.4不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗脯氨酸含量的影响
前人研究表明,脯氨酸是逆境胁迫下生物体特别易积累的一种氨基酸之一,也能够作为一种渗透调节物,参与稳定蛋白结构及保护细胞内生物大分子等作用。同时,在同一逆境胁迫下,不同生物体表现出较大的差异性。本试验研究表明(图4),沙生孑遗植物扁桃两个亚种长柄扁桃和蒙古扁桃在三种不同胁迫处理下,随着干旱加重,脯氨酸呈现出先稳定不变后迅速升高的趋势,特别是在中度胁迫(LS)和重度胁迫(SS)下,两个品种的脯氨酸含量极显著地高于对照(P<0.01),且在重度干旱下,蒙古扁桃积累的脯氨酸要比长柄扁桃高,说明其抗旱能力要略强。

图4 干旱胁迫对长柄扁桃和蒙古扁桃幼苗脯氨酸含量的影响
2.2.5 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗可溶性糖含量的影响
可溶性糖是植物有机溶质的碳架和能量的主要来源之一,同时也作为多种生物体的渗透调节剂,参与细胞膜和原生质胶体的稳定形成。一般条件下,植物体内可溶性糖含量较低,但在生物胁迫或非生物逆境下,可溶性糖会在细胞中大量累积,为植物的各种新陈代谢提供较为稳定的内环境。研究表明,可溶性糖主要参与细胞的渗透调节作用,其含量可在一定程度上反映植株的耐旱性能。如图5所示,在轻度胁迫下,两种扁桃的可溶性糖含量开始显著升高(P<0.05),随着胁迫程度加强,在中度和重度胁迫下,长柄扁桃和蒙古扁桃的可溶性糖含量迅速升到很高的水平,且极显著的高于对照(P<0.01),特别是在重度干旱下,可溶性糖含量已经是对照的3-4倍以上,说明干旱处理显著促进了长柄扁桃的可溶性糖的积累。
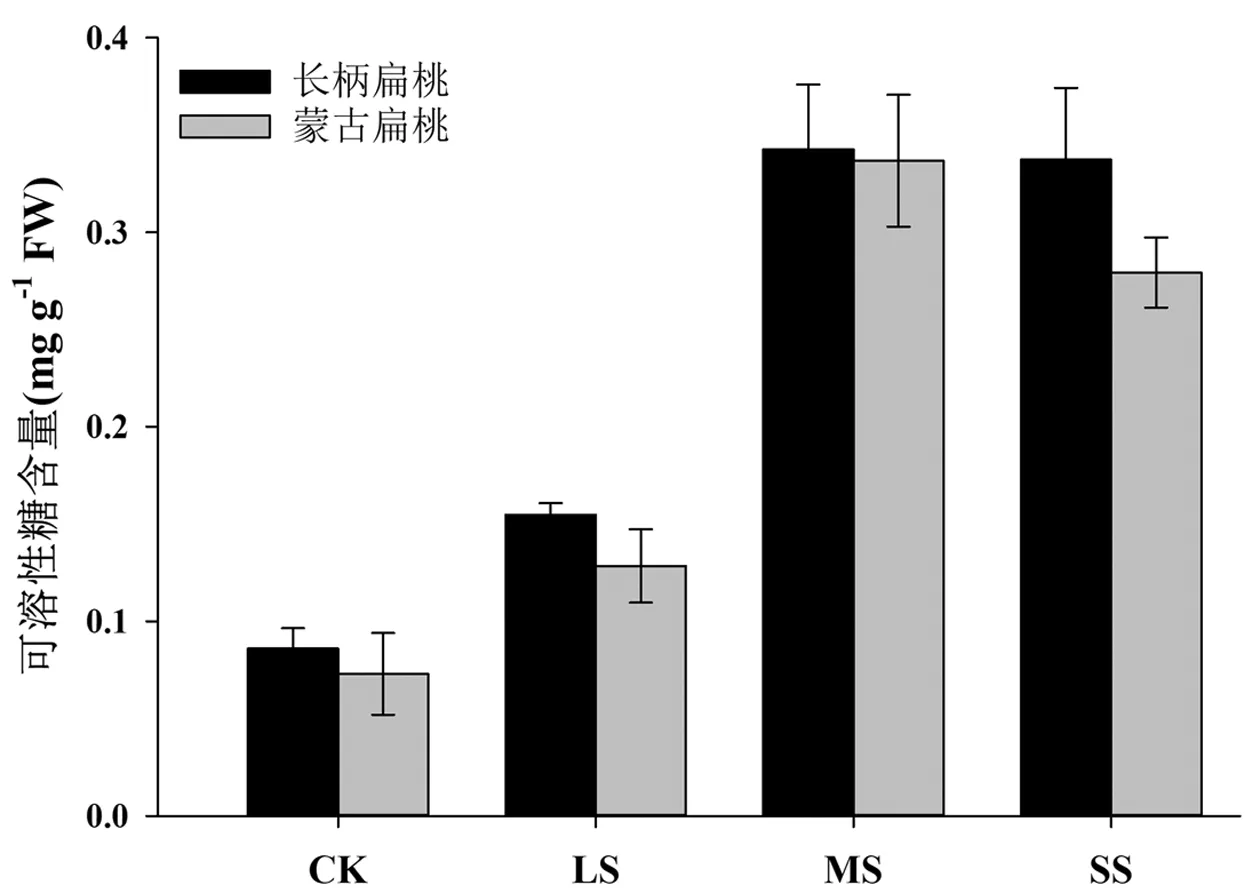
图5 干旱胁迫对长柄扁桃和蒙古扁桃幼苗可溶性糖含量的影响
2.2.6 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗叶绿素含量的影响
植物叶绿素的含量在一定程度上亦能反映出植物体是否遭到或遭受到多大生物非生物逆境胁迫的威胁。由图6可以看出,两种长柄扁桃在不同程度的干旱胁迫下,叶绿素含量呈现出先稳定不变后逐渐升高最后又缓慢回落的变化趋势。在轻度胁迫下,两种扁桃叶绿素含量没有受到影响,当中度干旱胁迫后,可能是植物的细胞膜遭到破坏,叶绿素浸提速率极显著升高(P<0.01),当干旱进一步加剧后,植物可能产生了某种生理防御机制来抵抗外界胁迫,从而使得叶绿素浸提速率有所回落。
2.2.7 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗POD含量的影响
过氧化物酶(POD)是生物体内普遍存在且活性较高的一种酶之一,其可催化过氧化氢、氧化酚类等化合物,参与消除氧代谢中产生的过氧化氢和酚类、胺类等毒性物质,进而起到解毒的作用,是植物体内重要的的保护酶之一。由图7可以看出,两种长柄扁桃在不同程度的干旱胁迫下,POD量呈现出逐渐升高的变化趋势。在轻度胁迫下,两种扁桃POD含量显著增高(P<0.05),当中度干旱胁迫后,两个品种的POD含量极显著升高(P<0.01),当重度胁迫后,材料的POD含量达到最大值,为对照的5~5.5倍。

图6 干旱胁迫对长柄扁桃和蒙古扁桃幼苗叶绿素含量的影响
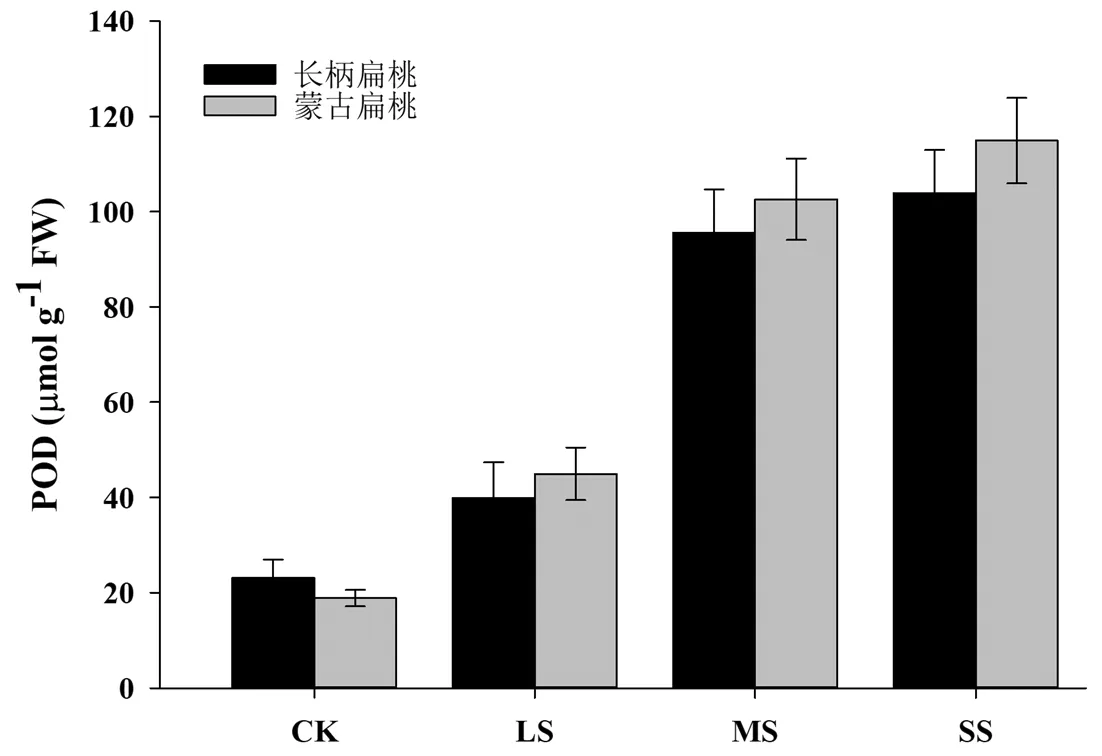
图7 干旱胁迫对长柄扁桃和蒙古扁桃幼苗POD含量的影响
2.2.8 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗SOD含量的影响
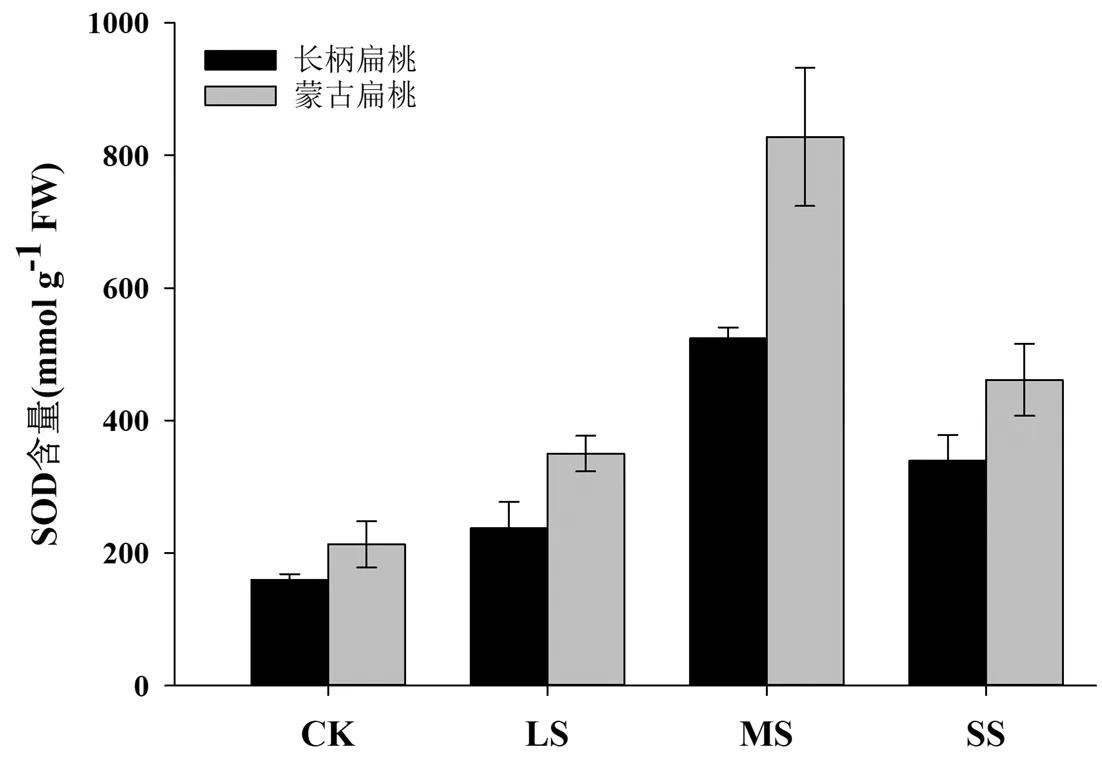
图8 干旱胁迫对长柄扁桃和蒙古扁桃幼苗SOD含量的影响
SOD( 超氧化物歧化酶) 是植物体内重要的保护氧自由基对细胞膜系统伤害的金属酶类。由图8可以看出,两种长柄扁桃在不同程度的干旱胁迫下,SOD含量呈现出先升高后降低的变化趋势。在轻度胁迫下,长柄扁桃和蒙古扁桃SOD含量显著增高(P<0.05),当中度干旱胁迫后,两个品种的SOD含量极显著升高(P<0.01),并且达到最大值,此时SOD含量分别为对照处理的3.3倍和3.9倍。当重度胁迫后,长柄扁桃和蒙古扁桃的SOD含量逐渐降低,说明此时干旱胁迫已经造成了细胞膜的严重破坏。进一步说明长柄扁桃幼苗能在水分胁迫下SOD活性维持在较高的水平,这对防止活性氧的大量积累,降低膜质过氧化程度,减轻膜伤害具有重要作用。
2.2.9 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗MDA量的影响
MDA是膜脂过氧化的产物其含量的高低可以作为细胞膜系受损伤程度的指标之一,也是衡量植物抗性强弱最直观的反应指标。研究表明,干旱胁迫下植物细胞膜受到损伤后会产生大量自由基,促进了MDA含量升高。由图9可以看出,两种长柄扁桃在不同程度的干旱胁迫下,MDA含量呈现出逐渐升高的变化趋势。在轻度胁迫下,长柄扁桃和蒙古扁桃MDA含量显著增高(P<0.05),当中度干旱胁迫后,两个品种的MDA含量极显著升高(P<0.01),当达到重度胁迫时,长柄扁桃和蒙古扁桃的MDA含量达到最大值,分别为对照处理的5倍和4.5倍。

图9 干旱胁迫对长柄扁桃和蒙古扁桃幼苗MDA含量的影响
2.2.10 不同干旱胁迫对长柄扁桃和蒙古扁桃幼苗CAT含量的影响
过氧化氢酶(CAT)普遍存在于植物组织中,是一种酶类清除剂,又称为触酶。其主要功能是参与清除植物体内的过氧化氢,从而使细胞免于H2O2的毒害,是生物防御体系的关键酶之一。由图10可知,长柄扁桃幼苗的CAT活性随着干旱胁迫的增加而呈现出上升趋势,不同基因型扁桃在受到干旱胁迫后,长柄扁桃的CAT活性总是略低于蒙古扁桃,但差异不显著(P>0.05)。进一步说明重度干旱胁迫时会对长柄扁桃和蒙古扁桃幼苗造成严重伤害并引起幼苗CAT活性的显著上升。

图10 干旱胁迫对长柄扁桃和蒙古扁桃幼苗CAT含量的影响
2.3 干旱胁迫对长柄扁桃和蒙古扁桃幼苗叶片表皮蜡质总量的影响
植物体对逆境胁迫的重要响应之一是调控并分泌蜡质到植物表皮层,并形成不同类型的蜡质晶体。我们的研究表明(图11),当长柄扁桃和蒙古扁桃遭受到不同程度的干旱胁迫后,其蜡质总量发生了变化,其变化规律是:总蜡质含量随着干旱胁迫的加深呈现出先稳定不变后显著升高的规律,当中度胁迫后,蜡质总量达到最大值。说明只有植物在中度和重度胁迫条件下,长柄扁桃和蒙古扁桃的表皮蜡质才能体现出抗旱保水的作用来。此外通过对两个品种的比较,发现在中度和重度干旱胁迫下,蒙古扁桃表皮蜡质的积累量要高于长柄扁桃(图11),因此,单从蜡质积累方面说明了蒙古扁桃对干旱的适应性要比长柄扁桃好。

图11 干旱胁迫对长柄扁桃和蒙古扁桃幼苗表皮蜡质积累的影响
3讨论与结论
3.1 干旱胁迫对长柄扁桃与蒙古扁桃种子萌发的影响
植物在生长和发育过程中受多种非生物胁迫(如干旱、高温、高盐等)的影响,其中,干旱是限制植物生长发育的主要非生物因素。在世界范围内的许多农业领域水分缺少显著限制植物的生长和产量。干旱作为植物所遭受的所有非生物胁迫中损害最为严重的不利因素,直接影响世界农业的生产。种子萌发是植物生长发育阶段关键达的环节之一,种子萌发策略对荒漠地区植物的种群更新及植被恢复都具有重要意义。
目前国内外学者采用PEG模拟干旱胁迫在牧草、黄麻、黄瓜、亚麻等作物的种子抗旱性方面研究较多。而在长柄扁桃和蒙古扁桃抗旱综合性研究等方面还未见报道,因此,本研究先采用PEG模拟干旱胁迫对抗旱能力都较强的长柄扁桃和蒙古扁桃种子萌发过程中的抗旱能力进行研究,以期筛选出抗旱性更强的材料,为揭示长柄扁桃耐瘠的分子、生理机制,加强干旱半干旱地区抗旱植物的种植利用提供科学依据。结果表明,轻微的干旱不会对种子的萌发产生重要影响,相反会促进两种扁桃种子的萌发及成苗过程,这与姜生秀等研究沙冬青的结果一致。然而中度和重度胁迫条件下,两种扁桃的发芽率、发芽势、发芽指数、活力指数、幼苗高度、根长和幼苗鲜重均表现出极其相似的规律,即随着干旱胁迫的加重表现为各形态学指标受抑制程度越明显。另外在同等条件的干旱胁迫下蒙古扁桃受抑制程度要小于长柄扁桃,进而得出初步的结论:蒙古扁桃的抗旱性要高于长柄扁桃。
3.2 不同干旱胁迫对两种扁桃幼苗生理生化指标的影响
植物在不同的生育期对外界环境都有不同的响应,苗期抗旱性研究是其中最重要的组成部分。植物体在不同的逆境条件下,很多生理生化指标如株高、净光合速率、气孔开张度、叶绿素含量、脯氨酸含量、可溶性蛋白含量、POD、SOD、MDA和CAT等都会发生不同程度的变化。前人研究表面,植物的抗旱性与以上各种指标密切相关。本项目在前期研究发现两种不同的扁桃属植物抗旱性很强的基础上,拟对其在不同干旱胁迫下的抗旱生理生化指标进行测定,结果表明,在轻度、中度和重度胁迫条件下,除光合速率、株高和气孔开张度外,其他7种生理指标均表现出迅速增长的趋势,这与前人在小麦、多油辣木和藜麦上的研究结果相似[13-15]。在逆境胁迫加大的情况下,长柄扁桃和蒙古扁桃的净光合速率呈显著下降趋势;株高也呈现出降低的趋势,但不显著。
此外,研究在长柄扁桃和蒙古扁桃上测定了其叶表皮气孔随着干旱胁迫的变化情况,结果表明,随着干旱胁迫的不断加重,叶表面气孔呈现出先稳定不变后迅速关闭的趋势,但在两种材料上亦存在差异,即蒙古扁桃在中度干旱下,气孔没有完全关闭,表明其还能进行光合作用,说明蒙古扁桃对中度胁迫具有一定的抵御能力,这与Guo等[16]研究结果相似。
作为生物体最外层的表面,植物表皮层蜡质是与外界环境密切接触的直接部位,它对植物与外界交流、植物的生长与发育等具有重要的意义。同时蜡质在植物自身适应逆境及其生物非生物胁迫中也扮演着极其重要的角色。植物在长期适应逆境胁迫的过程中,通过自身代谢,会产生生理或形态的变化来避开逆境所造成的伤害。众所周知,减少或阻止非气孔性水分的散失是角质层最初的生理作用。角质层的保水功能是不容忽视的,当把角质层蜡质从角质层膜上去除后,角质层膜的水分渗透率会显著性改变。Mohammadian等[17]有选择性的从植物Leucadendron lanigerum的离体叶片中剥离了上表皮蜡质,从而导致其蒸腾速率显著升高,表明表皮蜡质层是蒸腾失水的天然屏障。特别是在干旱少雨的条件下,植物在其表皮积累更多的蜡质进而减少非气孔性水分的散失来抵御干旱胁迫。本研究中,笔者用氯仿浸提法提取了长柄扁桃和蒙古扁桃的表皮蜡质,并且粗略的进行了测定,结果表明,当长柄扁桃和蒙古扁桃遭受到不同程度的干旱胁迫后,其蜡质总量发生了变化,其变化规律是:总蜡质含量随着干旱胁迫的加深呈现出先稳定不变后显著升高的规律,当中度胁迫后,蜡质总量达到最大值。综合上面所述,蒙古扁桃比长柄扁桃更具有抗干旱胁迫和耐瘠薄的能力,因此在沙漠地带可以优先推广种植。
猜你喜欢
麦类作物学报(2023年6期)2023-06-20 00:56:56
疯狂英语·新读写(2022年4期)2022-11-22 09:10:30
疯狂英语·读写版(2022年4期)2022-04-08 00:22:23
花火·慧阅读(2022年4期)2022-03-18 09:27:10
中成药(2018年6期)2018-07-11 03:01:20
作物学报(2016年11期)2016-11-09 08:43:12
甘肃林业(2016年3期)2016-11-07 08:56:28
国外医药(抗生素分册)(2016年2期)2016-07-12 14:25:01
西北林学院学报(2015年5期)2015-01-03 05:12:24
中国粮油学报(2014年8期)2014-02-06 01:34:14
