
平潭岛东北部近岸海域大型污损生物群落结构特征
2020-07-21 05:34刘坤林和山李众何雪宝黄雅琴林俊辉牟剑锋张舒怡马丽全俊杰王建军孙军
海洋学报 2020年6期
刘坤,林和山,李众,何雪宝,黄雅琴,林俊辉,牟剑锋,张舒怡,马丽,全俊杰,王建军*,孙军
( 1. 天津科技大学 生物工程学院,天津 300457;2. 自然资源部第三海洋研究所 海洋生物与生态实验室,福建 厦门361005;3. 厦门大学 嘉庚学院 信息科学与技术学院,福建 漳州 363105)
1 引言
海洋大型污损生物是指栖息、附着和生长在海中船体和其他人为设施表面的动植物的总称,主要种类包括藻类、水螅虫、盘管虫、牡蛎和藤壶等,海洋生物附着或栖息在海中设施表面并产生不利影响的现象和过程称为生物污损[1-2]。污损生物对海洋经济开发的负面影响早已引起人们的广泛关注,其危害主要包括增加船舶的阻力、堵塞管道、加速金属腐蚀和对声学仪器的影响等方面[2-3];从生态研究角度来看,污损生物是海洋生态系统的重要组成部分,其在物质循环、能量流动和水体净化方面具有重要作用[4]。
我国大型污损生物生态学的研究始于20世纪50年代,主要涉及渤海、黄海、东海和南海等海域的多个海湾和港口[5],其中东海沿岸大型污损生物的研究主要涉及物种多样性分布、附着机理、污损生态效应及物种入侵等方面的内容[2]。近年来,很多研究通过挂板试验系统分析了宁德海域[6]、湄洲湾[7]、洋山港[5]、北关港[8]等东海港湾的污损生物物种组成和数量分布,部分研究涉及了污损生物的生活方式、摄食功能群及其二者之间的关系。
平潭岛,亦称海坛岛,位于福建省平潭市东部,地处台湾海峡交通要道,地理位置优越,是福建省第一大岛,其周边海域属于开敞式海湾,受外海流系影响较大。目前平潭岛附近海域大型污损生物的报道仅见于1963-1964年岛西南部竹屿口海域的挂板试验,分析了该海域污损生物的多样性和优势种组成[2],而对于遮蔽程度更低、受外海海流影响更大的东北部海域则缺乏相关研究。另外,随着经济的快速发展,人类活动的影响日趋严重,了解平潭岛周边海域海洋生物群落结构和多样性状况可更好地为合理开发利用海洋资源奠定基础。
因此,本文拟通过在平潭岛东北部海域的周年挂板试验,对该海域污损生物的群落结构、生活方式、摄食功能群现状和群落的季节更替等特征进行系统研究,分析其群落组成和优势种的时空变化规律,以期为东海沿岸海洋设施污损生物的防除和海洋生态系统的保护提供相关理论基础。
2 材料与方法
2.1 调查区域
于2017年6月至2018年5月在平潭岛东北部近岸海域(25.578 8°N, 119.857 7°E)进行污损生物周年挂板试验,调查位置如图1所示,水深约为20 m。该试验站位东临台湾海峡,西隔海坛海峡,周边岛屿众多,地形复杂;主要的水团活动有浙闽沿岸流、台湾暖流和附近的上升流,加上台湾海峡的狭管效应,使得附近海域海流复杂,结构形态多变[9]。
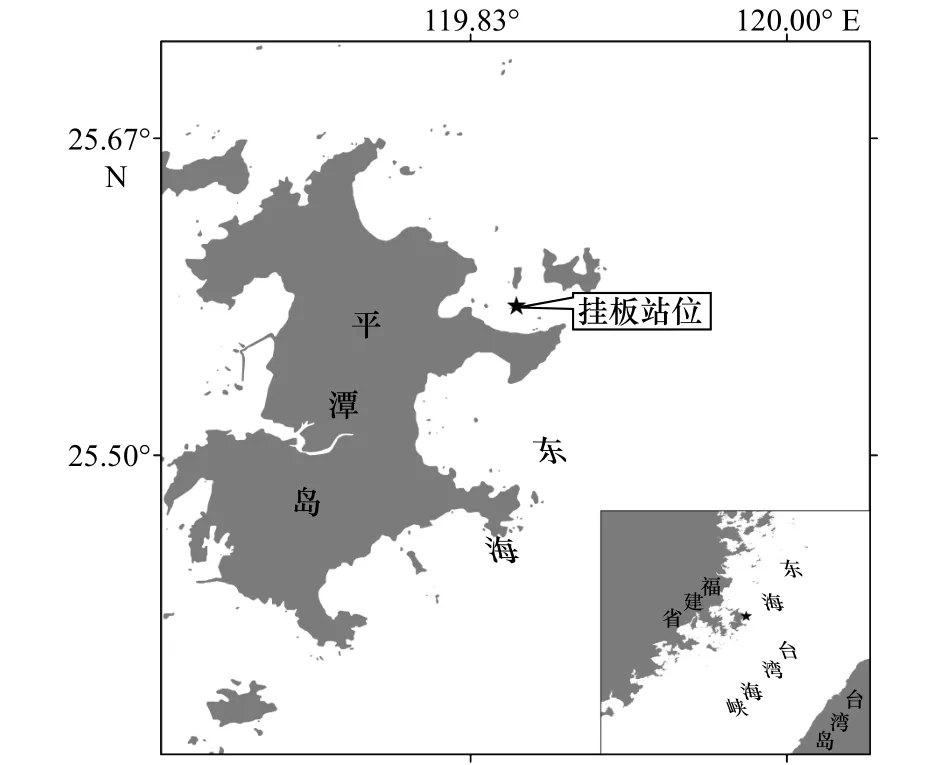
图1 污损生物挂板试验站位示意图Fig.1 The location of macro-fouling investigation
2.2 采样方法
参照Lin等[6]的方法,采用160 mm(长度)×160 mm(宽度)×20 mm(厚度)的混凝土试板,试板分左右两面,设A、B两个平行组挂放。季板、半年板和周年板的收放周期分别按每季、每半年和1周年进行。每组试板均含表层和底层,且试板面与海面垂直;表层试板上沿与水面齐平,底层试板置于水深5 m。全年共回收试板23块,包括季板13块,半年板7块,周年板3块。样品处理和分析及资料整理均按《海洋调查规范》(GB/T 12763.6-91)要求进行,其中厚度值为测量试板的4个角落和中央5个点的平均厚度;覆盖面积率为整个群落覆盖附着基的百分比率;密度值为逐种记录单位面积的个体数;生物量为吸去外表水分后的质量。使用YZY4型温盐传感器进行温度数据的采集,盐度测量只在2018年3月6日(春季)、2017年6月5日(夏季)、2017年9月2日(秋季)和2017年12月1日(冬季)进行。物种名称与WoRMS网站(http://www.marinespecies.org/)进行比对并统一。
2.3 数据分析
采用相对重要性指数(Index of Relative Importance,IRI)进行群落优势种分析[10],IRI≥100 作为优势种的判别标准,公式如下:
式中,W为物种的生物量占总生物量的百分比,N为物种的丰度占总丰度的百分比,F为该物种的出现频率。
除大型藻类等初级生产者之外,其他污损生物根据其摄食习性,划分为5类功能群[11-13]:悬浮物食者、植食者、肉食者、杂食者和碎屑食者。按照生活方式,将污损生物分为固着、半固着和自由活动等3种方式[3]。
利用PRIMER6.0软件包,基于物种-生物量数据进行Bray-Curtis相似性系数的计算,构建相似性矩阵,用组平均法进行分等级聚合聚类分析和非度量多维标度(Nonmetric Multidimensional Scaling,NMDS),分析之前将生物量数据进行四次方根转化。采用胁迫系数(Stress)来衡量NMDS分析结果是否具有参考价值:Stress<0.1,可以正确解释样本间的相似性关系;0.1<Stress<0.2,具有一定的解释意义和参考价值;Stress>0.2,不能解释样本间的相似性关系[14]。
使用单因素方差分析(One-way ANOVA)进行污损生物物种数、密度和生物量等参数的季节和试板差异。图表绘制使用ArcMap 10.3软件。
3 结果与分析
3.1 环境因子
挂板试验站位的月平均水温变幅为11.0~27.8℃,9月份(秋季)温度最高,2月份(冬季)最低(图 2)。盐度在4个季节变幅为32.5~34.2,冬季盐度最低,春季最高,夏、秋两季盐度分别为33.23和33.89。
3.2 物种组成
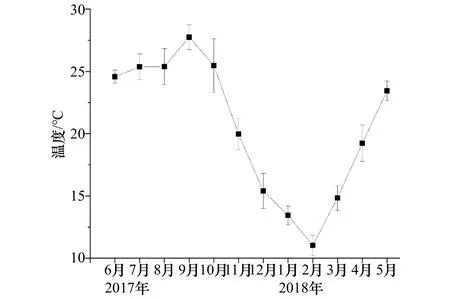
图2 试验站位2017年6月至2018年5月温度变化趋势Fig.2 The temperature in sampling location during June 2017 to May 2018
23块水泥试板共鉴定大型污损生物13门92种,其中节肢动物31种、环节动物23种、软体动物17种,三者构成该海域污损生物的主要优势类群;另外还有大型藻类5种、刺胞动物5种、扁形动物2种、纽形动物2种、星虫动物1种、苔藓动物3种、棘皮动物2种和脊索动物1种(附表A1,附表A2)。
营固着生活的网纹藤壶(Amphibalanus reticulatus)的优势度最大(IRI=4 116),其次是自由生活的廉形叶钩虾(Jassa falcata),二者摄食类型均以悬浮颗粒物为主。其他优势种分别为固着生活的大室别藻苔虫(Biflustra grandicella)、三角藤壶(Balanus trigonus)和中胚花筒螅(Ectopleura crocea),自由生活的齿掌细身钩虾(Maeropsis serratipalma)、加尔板钩虾(Stenothoe gallensis)、细毛背鳞虫(Lepidonotus tenuisetosus)和四角细身钩虾(Quadrimaera quadrimana),以及附着生活的翡翠贻贝(Perna viridis)(表 1)。
3.3 附着量及时空变化
研究海域全年均有生物附着,但附着种类和数量的季节性差异明显,附着盛期为6月至8月(生物量介于7326.0~12 970.0 g/m2之间),12月至次年 2月为附着淡季(生物量介于39.5~1 580.5 g/m2之间)。表层和底层板附着生物的密度(F=6.281,p<0.05)和生物量(F=4.568,p<0.05)均具有显著性差异。
表层板中,春季的附着厚度为最高((17±22)mm),秋季最低((4±2)mm);上半年((17±3)mm)和下半年((15±2)mm)的附着厚度差异不大;周年板的平均附着厚度可达(24±10)mm。秋季的平均覆盖面积为最低(86%±11%),其余季板、半年板和周年板的覆盖面积均达百分百(表2)。
表层板附着密度不同季节间呈极显著性差异(F=18.453,p<0.01),其中夏季最高((26 625±7 319)ind./m2),秋季为最低((2 714±2 423)ind./m2);下半年的附着密度((18 863±2 000)ind./m2)明显高于上半年((5 575±768) ind./m2); 周 年 板 的 附 着 密 度 高 达 (99 625±33 650)ind./m2。在生物附着量(湿质量)方面,夏季最高,可达(11 917.8±1 488.1)g/m2,与其他3个季节有极显著性差异(p<0.01);下半年((11 049.9±1 413.9)g/m2)也明显高于上半年((1 061.0±410.9)g/m2);周年板的附着生物量可达(11 005.6±4 113.9)g/m2。节肢动物、藻类、刺胞动物和苔藓动物为最主要的生物量贡献类群(表 2)。
底层板中,春、夏、秋、冬季的平均附着厚度差异不大(p>0.05),介于12~15 mm 之间;上半年((10±5)mm)的附着厚度略低于下半年((15±1)mm);周年板的平均附着厚度高达(38±17)mm。除了秋季(74%±37%)和冬季(24%±10%)的试板外,春、夏季板和半年及周年试板的覆盖面积均达百分百(表3)。
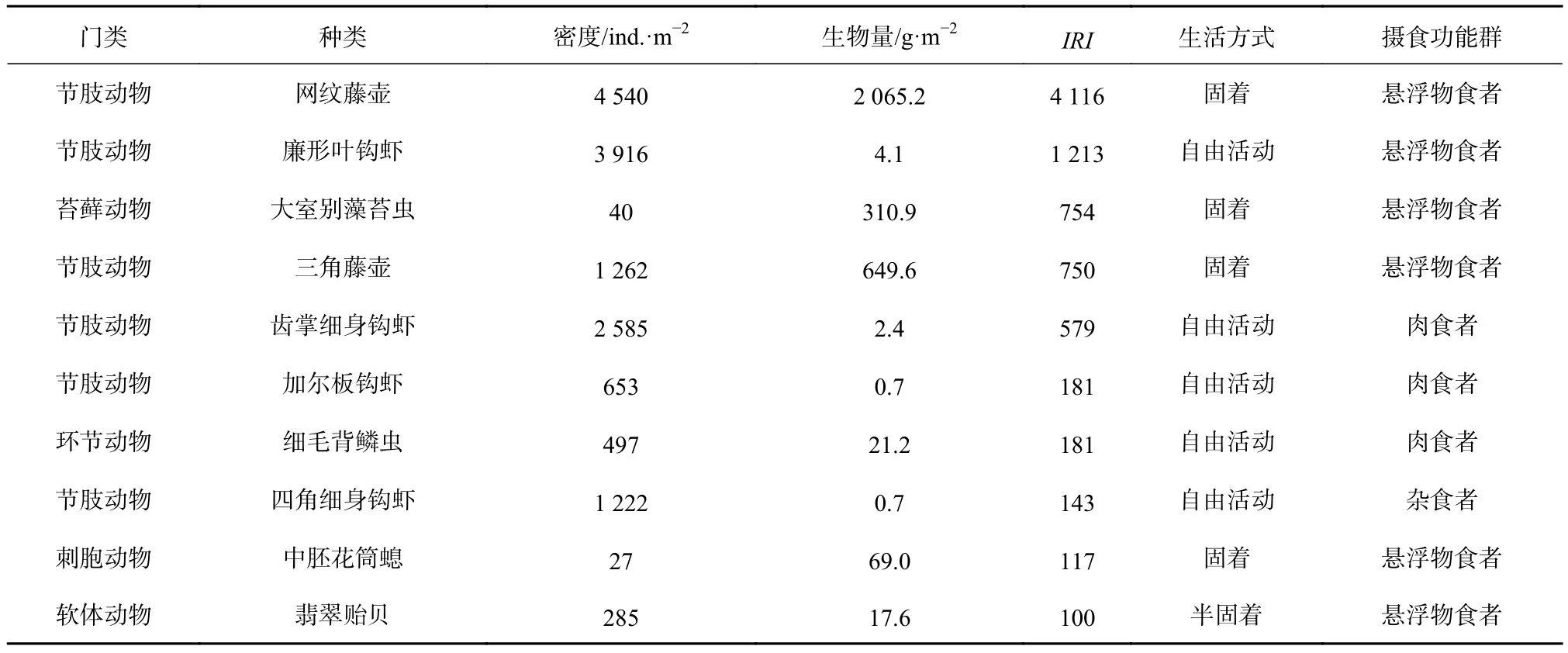
表1 污损生物优势种Table1 The dominant species of the macro-fouling
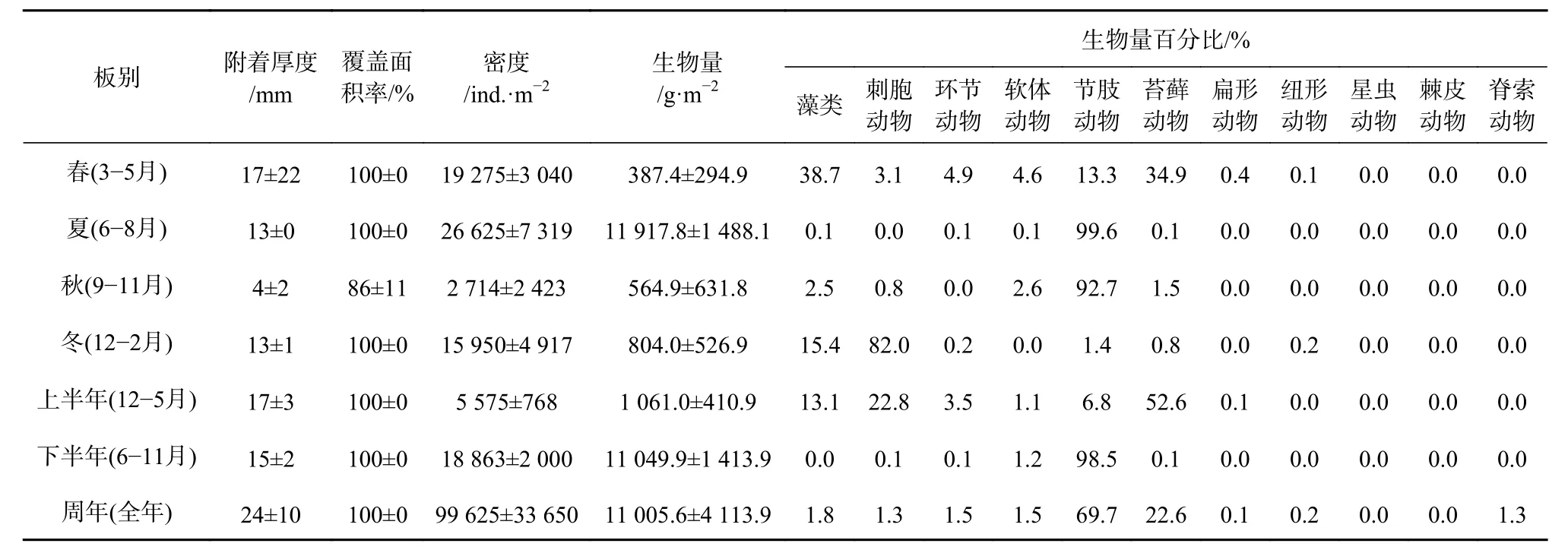
表2 表层板大型污损生物附着状况Table2 The settlement level of the macro-fouling on the top test panels
底层夏季板的附着密度为最高,可达(35 575±16 228)ind./m2,与其他季节有显著性差异(p<0.05);下半年的附着密度((16 275±1 520)ind./m2)高于上半年((7 925±1 785)ind./m2);周年板的附着密度为(10 660±1 966)ind./m2。夏季的附着生物量与其他季节有极显著性差异(p<0.01),可达(9 929.0±3 681.2)g/m2,其中冬季最低,仅为(93.4±36.0) g/m2;下半年的附着生物量((7 584.8±906.2)g/m2)高于上半年((1 099.0±677.4)g/m2);周年板的附着生物量为(4 421.3±505.2)g/m2;节肢动物、刺胞动物和苔藓动物为最主要的生物量贡献类群(表 3)。
3.4 群落的季节更替
根据50%的相似度将污损生物群落划分为5个聚类组(图3),NMDS显示Stress值为0.11,分析结果相对可靠;单因素相似性分析表明,5个聚类组间的物种组成呈显著性差异(Global R=0.989,p=0.001)。聚类组Ⅰ(网纹藤壶群落)出现在表、底层的秋季板上,以固着生活的种类为主,主要出现在温度较高的夏、秋季(图4)。聚类组Ⅱ(网纹藤壶-翡翠贻贝群落)出现在表、底层的夏季板和下半年板上,主要附着季节是温度较高的6-11月,附着强度较大,以固着和半固着生活的种类为主,其中优势种网纹藤壶附着生物量贡献率为39.5%,夏季表层板中附着密度和生物量最大可达29 300 ind./m2和12 900.0 g/m2。聚类组Ⅲ(大室别藻苔虫-细毛背鳞虫群落)出现在表、底层的春季板和上半年板上,主要附着季节是温度较低的12月至次年5月,附着强度较小,以固着和活动性生活的种类为主,而且肉食性生物细毛背鳞虫常栖息于其他固着类生物群落内。聚类组Ⅳ(三角藤壶-大室别藻苔虫群落)出现在表、底层的年板上,附着强度大,固着、半固着和活动性生活种类均大量存在;优势种三角藤壶在年板的表层板中附着密度和生物量最大可达10 600 ind./m2和12 720.0 g/m2。聚类组Ⅴ(中胚花筒螅-大室别藻苔虫群落)出现在表、底层的冬季板上,主要附着季节为温度较低的冬季,附着强度小,以固着和半固着生活的种类为主。
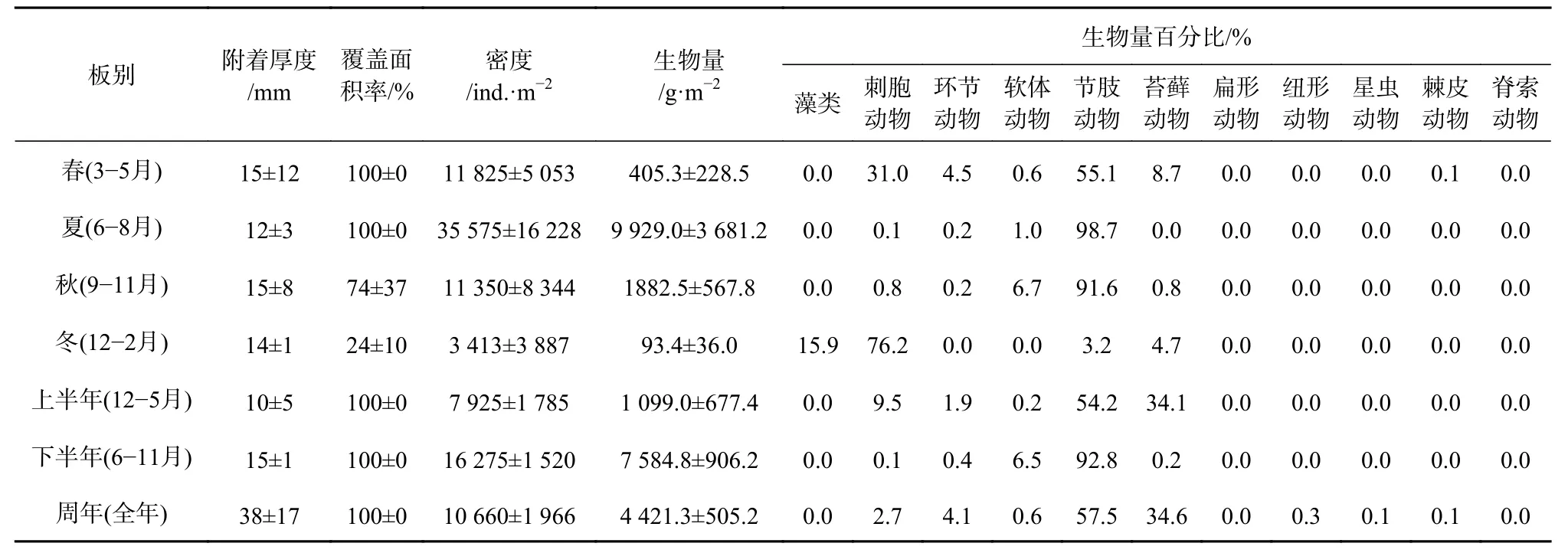
表3 底层板污损生物附着状况Table3 The settlement level of the macro-fouling on the bottom test panels
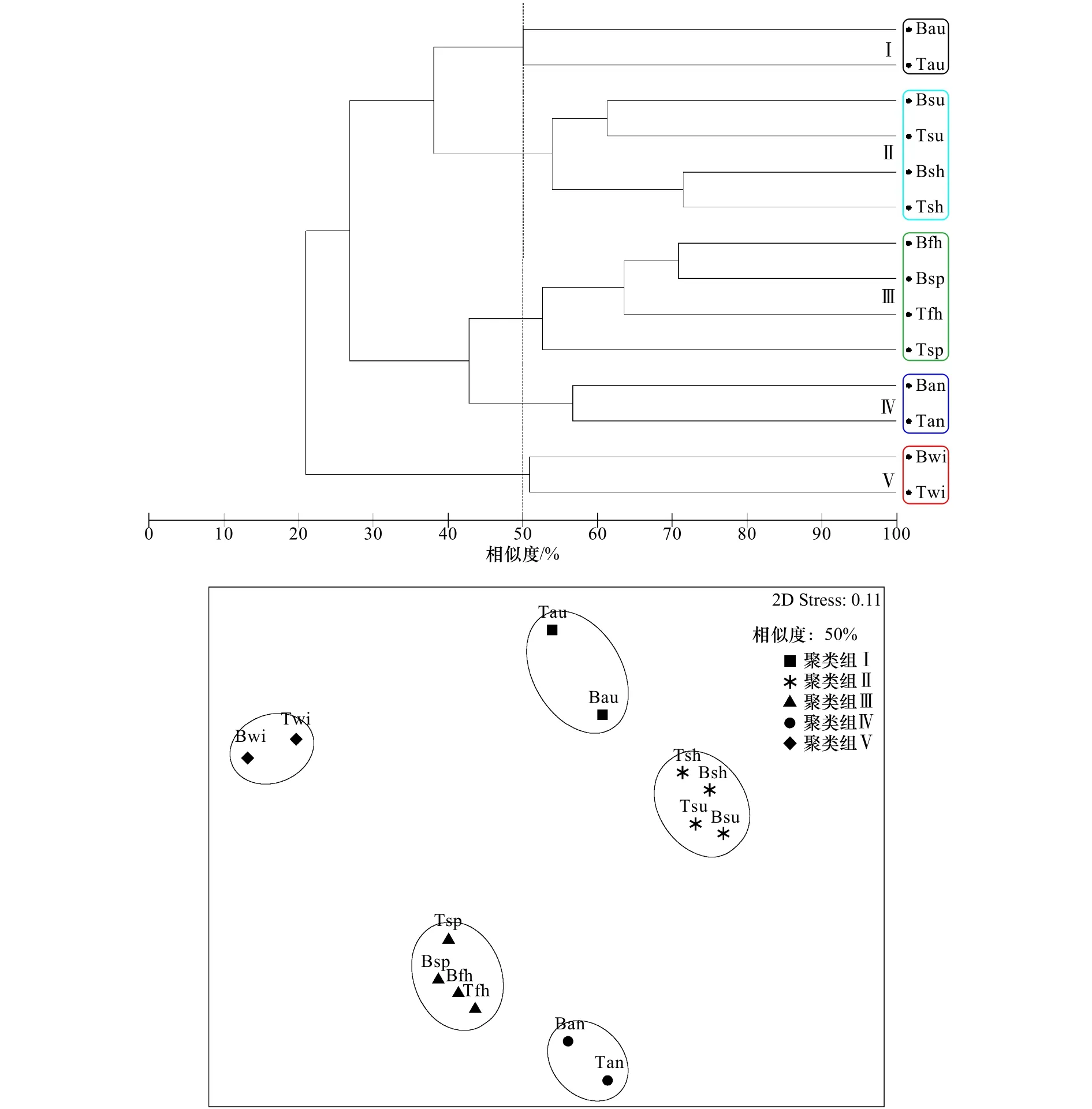
图3 污损生物群落聚类分析和非度量多维标度排序Fig.3 Cluster and nonmetric multidimensional scaling plots based on biomass of the macro-fouling
3.5 功能群组成
研究海域表、底层试板污损生物均以悬浮物食者为主(图5),其中春季表层板悬浮物食者占总附着生物量的54.5%,其他季节试板上悬浮物食者的附着量均占总生物量的80%以上。网纹藤壶、三角藤壶、大室别藻苔虫和中胚花筒螅为悬浮物食者的优势代表种,其中大室别藻苔虫和中胚花筒螅的附着高峰期均在12月至次年5月,网纹藤壶则为6-11月,三角藤壶仅在年板上大量附着(图6)。初级生产者主要出现在春季表层板上,占总附着生物量的38.7%。

图4 污损生物主要优势种的附着季节Fig.4 The settlement period and peak time of main fouling organisms
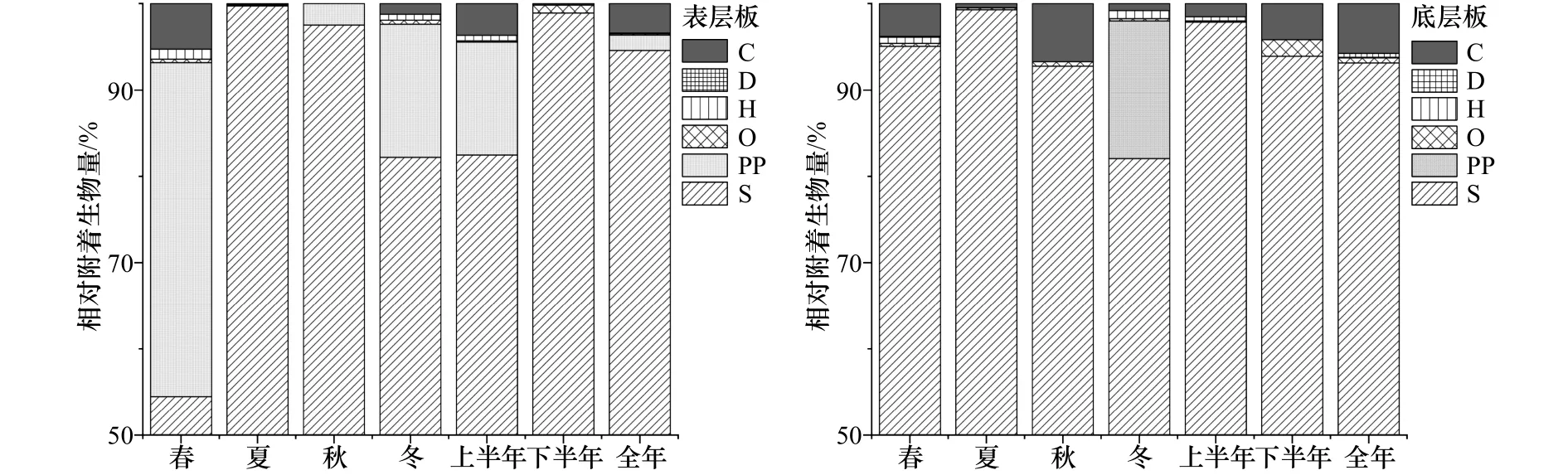
图5 大型污损生物群落中各摄食功能群所占百分比(相对附着生物量)Fig.5 The relative biomass of the macro-fouling functional feeding groups
污损生物群落按其活动情况和生活方式,可分成固着生活、半固着生活和活动性生活3种类型。在4个季节中表底层板污损生物均以固着型生物为主(图7),其附着生物量均占总生物量的80%以上。固着生物的食物以悬浮颗粒物为主,优势种主要为网纹藤壶、三角藤壶、大室别藻苔虫和中胚花筒螅,其主要分布时期见图4和图6。
4 讨论
平潭岛东北部近岸海域大型污损生物群落种类多且生物量大,以内湾型群落为主,兼具部分外海或高盐群落特点[2],其中节肢动物和苔藓动物为最主要的优势类群。研究海域污损生物种类组成以世界性广分布暖水种为主,其中固着型生物网纹藤壶是该海域最主要的优势种,附着季节主要是6-11月,盛期为7-8月,广泛分布于东海和南海潮间带至潮下带区域[15-16];大室别藻苔虫的分布遍及中国沿海,黄、渤海比南海多[2];钟巨藤壶广泛分布于我国长江口外至西沙海域,作为分布于开阔海域的一种巨型藤壶,在内湾很少分布。
黄宗国和蔡如星[2]在平潭岛西部海域挂板研究结果显示,共鉴定污损生物71种,平均生物量为10 785.2 g/m2,优势种主要为网纹藤壶、总合草苔虫(Bugula neritina)和管钩虾(Corophiidae und.)等,但外海性种类如钟巨藤壶(Megabalanus tintinnabulum )和红巨藤壶(Megabalanus rosa)等没有附着。与本研究结果相比,污损生物物种数较低,生物量较高,主要优势种则均为网纹藤壶;另外一个明显的差异是,本研究中出现了钟巨藤壶等外海性种类。主要原因可能是因为本研究挂板位置位于平潭岛东北部海域,离岸相对较远,盐度高且较稳定,一般维持在30以上,环境条件有利于部分外海性高盐种类的生存。
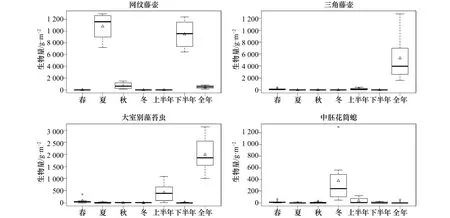
图6 优势种附着量状况Fig.6 The settlement level of dominant species

图7 不同类型污损生物的相对附着生物量Fig.7 The relative biomass of different lifestyle groups of macro-fouling
黄宗国和蔡如星[2]曾根据夏季底层板的附着量,把中国沿海22个港口的污损生物附着程度分为5个等级,研究海域夏季底层板的附着生物量达(9 929.0±3 681.2) g/m2,属于第二级,与泉州湾[1](11 295 g/m2)、湄洲湾[7](11 493.21 g/m2)、宁德海域[6](12 353.1 g/m2)、北关港[8](14 313.8 g/m2)和东山湾[17](11 799.0 g/m2)等东海近岸港口污损生物附着状况相似,远远高于受长江冲淡水影响较大的洋山港[5](426.6 g/m2),也明显大于黄、渤海沿岸的烟台内港[18]、大连港和青岛港[19]等海域,温度和盐度应是造成沿海港口污损生物附着量存在较大差异的主要原因。
生物种类温度属性的差异是中国沿海各海域之间污损生物差别在“质”方面的表现[2]。东海和南海污损生物种类与黄、渤海,特别是北黄海有很大不同,其中网纹藤壶和三角藤壶广泛分布于东海和南海的近岸海域,长江口是其在我国近岸海域的地理分布北界,而且越往南其附着期越长,在南海近岸甚至全年均可附着[19]。寒半藤壶(Semibalanus balanoides)、缺刻藤壶(Balanus crenatus)、紫贻贝(Mytilus edulis)和大型褐藻(Laminaria saccharina)等冷水种,呈环寒带或温带分布,其主要分布区域是黄海和渤海沿岸[2-3,20]。本研究调查海域地处亚热带,物种种类以亚热带暖水种为主,污损性双壳类中未见紫贻贝、长牡蛎(Crassostrea gigas)等温水种类出现,取而代之以翡翠贻贝和福建牡蛎(Magallana angulata)等为主。
盐度是影响污损生物分布的另一关键因素[21],不同盐度区污损生物的种类组成有很大差别。淡水种纤细蓆藻(Phormidium tenue)是河港区域的绝对优势种;淡水棒螅(Cordylophora lacustris)则是河口区域污损生物的主要优势种[22]。对于藤壶、泥藤壶(Fistulobalanus kondakovi)适应河口低盐水域,而网纹藤壶、三角藤壶和钟巨藤壶等则在东海和南海的高盐海域出现[2,20]。如果水体盐度降至18甚至更低,则会妨碍网纹藤壶幼虫的发育甚至导致其死亡[16]。在本研究中,网纹藤壶、三角藤壶和钟巨藤壶附着量较大且没有泥藤壶出现的现象,应与所处海域的盐度在32以上密切相关。
除此之外,影响污损生物生长的重要因素还应有水体透明度、光照和水深等方面。由于研究海域栖息的附着型藻类多为喜强光藻类(如浒苔和石莼等),水体透明度下降会减弱水下光照和降低初级生产力[23],从而影响藻类的附着[24],因此,本研究中大型藻类一般在表层且光照充足的试板上附着量较大。
因此,大型污损生物群落的形成和发展并非受单一环境因子的影响,而是在温度、盐度、水体透明度、水动力条件、地表径流和养殖等人类活动干扰等因素综合作用下,最终通过群落结构的演替达到相对稳定的状态。

表 A1 大型污损生物种类组成Table A1 The species composition of macro-fouling

表 A2 大型污损生物物种名录Table A2 The list of macro-fouling organisms

续表 A2

续表 A2

续表 A2
猜你喜欢
计算机仿真(2022年9期)2022-10-25
材料科学与工程学报(2022年4期)2022-08-25
食品研究与开发(2022年16期)2022-08-24
理化检验(物理分册)(2022年3期)2022-03-31
数学小灵通·3-4年级(2021年6期)2021-07-16
铀矿冶(2021年2期)2021-05-24
石油化工腐蚀与防护(2021年2期)2021-04-27
今日农业(2020年14期)2020-08-14
船海工程(2016年6期)2017-01-03
中国舰船研究(2016年3期)2016-09-02
