
生物和非生物因子对秋季海州湾长蛇鲻栖息地适宜性的影响
2020-07-21 05:34朱承之张云雷赛可韦雯钰谢姝妤夏铭璟任一平薛莹
海洋学报 2020年6期
朱承之,张云雷,3,赛可,韦雯钰,谢姝妤,夏铭璟,任一平,2,3,薛莹,3*
( 1. 中国海洋大学 水产学院,山东 青岛 266003;2. 青岛海洋科学与技术试点国家实验室 海洋渔业科学与食物产出过程功能实验室,山东 青岛 266237;3. 中国海洋大学 海州湾渔业生态系统教育部野外科学观测研究站,山东 青岛 266003)
1 引言
长蛇鲻(Saurida elongata)隶属于辐鳍鱼纲(Actinopterygii)、仙女鱼目(Aulopiformes)、狗母鱼科(Synodontidae)、蛇鲻属(Saurida),广泛分布于西北太平洋沿岸的中国、日本以及韩国海域,在中国沿海由北到南均有分布,是我国重要的近海底层暖温性经济鱼类[1-3]。长蛇鲻作为我国的主要经济鱼类之一,年产量较大,从渤海到南海海域均有捕获,通常栖息于底质为泥或泥砂的海区。其性凶猛,游泳迅速,以小型鱼类和幼鱼为食,但通常移动范围不大,一般不作远距离洄游。黄、渤海鱼群的繁殖期是每年的5-7月。长蛇鲻常年可捕获,以冬、春两季为捕捞旺季[3]。国内外对长蛇鲻的研究主要集中于形态学和遗传学[1]、肌肉分析[2]、食性[4]等方面,而对于长蛇鲻栖息地适宜性的研究尚未见报道。
栖息地适宜性指数(Habitat Suitability Index,HSI)于20世纪80年代提出之后,在渔业资源开发、管理、评估及保护等领域中得到广泛应用,已成为渔业科学研究的重要手段之一[5]。目前,HSI模型已被广泛用于物种管理、环境影响评价、丰度分布和生态恢复研究[6-8]。提升回归树(Boosted Regression Tree,BRT)是一种基于决策树的集成学习方法[9],已在渔业上有一定的应用,如单位捕捞努力渔获量(Catch Per Unit Effort,CPUE)标准化[10]、鱼类空间分布、丰度和多样性预测[11-12]、鱼类栖息地研究[13-14]、兼捕预测[15]等。与传统多元回归方法相比,提升回归树能自动拟合自变量的交互作用,且不易出现过度拟合,因此泛化误差较低,对于新数据的预测精度较高[11]。
研究鱼类的栖息分布,除了环境因子外,也需要考虑饵料因子的影响。鱼类通过摄食活动获得维持自身生存、生长和繁殖的能量,进而会影响鱼类的生活习性、洄游分布乃至种群的数量变动。在以往的研究中,HSI模型的构建多采用非生物因子,而较少考虑饵料生物的影响,可能会影响栖息地适宜性评估的效果。
本研究根据2011年和2013-2018年海州湾及其邻近海域的底拖网调查数据,通过BRT模型确定环境因子和饵料因子的贡献率,分别采用算术平均值法(Arithmetic Mean Method, AMM)和几何平均值法(Geometry Mean Method, GMM)构建海州湾长蛇鲻的HSI模型,并绘制HSI分布,分析探究长蛇鲻适宜栖息地的空间分布特征,旨在为海州湾长蛇鲻资源的保护和合理利用提供科学依据。
2 材料与方法
2.1 数据来源
样品来自于2011年和2013-2018年秋季(9-10月)在海州湾及其邻近海域进行的底拖网调查。采用分层随机取样的方法设计调查站位[16],即根据水深、纬度等因素的差异将调查海域分为5个区域(图1),每个航次在各区域中随机选取一定数量的站位。其中2011年设置24个站位,2013-2015年对站位设置进行了优化,设置18个站位[17]。调查船为220 kW单拖渔船,拖速2~3 kn,拖曳时间约1 h,调查网具网口高度约6 m,网囊网目约17 mm。在每一调查站位使用CTD同步测定水深、水温、盐度等环境数据。样品的采集、处理和分析按照《海洋调查规范》进行[18]。在进行数据分析前对调查数据进行拖速2 kn,拖曳时间1 h的标准化处理,采用渔获密度(单位:ind./km2)作为相对资源密度指数。

图1 海州湾调查区域Fig.1 Sampling areas in the Haizhou Bay
2.2 适宜性指数计算
根据前期研究的结果,海州湾长蛇鲻的食性类型以游泳动物为主,鱼类是其最主要的饵料类群,还摄食少量的虾类和头足类等[19]。根据海州湾长蛇鲻的食物组成[20],本研究选取枪乌贼(Loligo spp.)、戴氏赤虾(Metapenaeopsis dalei)和六丝钝尾鰕虎鱼(Amblychaeturichthys hexanema)这3种主要饵料生物作为饵料因子进行研究。
首先开发基于长蛇鲻渔获密度(ind./km2)[21]的适宜性指数(Suitability Index, SI),然后采用平滑函数拟合环境变量(底层水温、底层盐度、水深和饵料)与SI的关系[22],SI的取值范围为 0~1。SI值越接近 1,表示适宜性指数越高,SI越接近0,表示适宜性指数越低。SI值处于0.7~1时所对应的环境因子被认为是最适栖息环境范围[23],SI值的计算公式为

2.3 栖息地适宜性指数模型
HSI是由多个SI综合计算所得。根据BRT模型的结果对不同因子赋予权重,采用的2种HSI计算方法,即AMM[24]和GMM[25]。计算公式分别如下:
式中,HSI为栖息地适宜性指数,SIi为环境变量i的SI值,ωi为环境变量i通过BRT模型被赋予的权重,n为环境因子的个数。
为评估环境因子与饵料因子组合模型的预测性能,将环境因子和饵料因子进行多种组合分别构建HSI模型,通过交叉验证对比各模型的预测性能。
2.4 模型的验证
采用交叉验证的方法评估HSI模型的预测性能。随机选取80%的数据进行建模,用剩余20%的数据进行交叉检验,并评估该模型的预测性能,重复进行100次[26]。对基于AMM和GMM算法的HSI模型,使用赤池信息准则(Akaike Information Criterion,AIC)评估预测性能,选取适合的最优模型。
2.5 绘制 HSI分布图
FVCOM模型(Finite-Volume Community Ocean Model)是由马萨诸塞州立大学达特茅斯分校和伍兹霍尔海洋研究所联合开发的沿海海洋环流模型,具有无约束、非结构化网格、有限体积、自由表面等优点[27]。本研究结合海州湾FVCOM模型中的环境数据,预测长蛇鲻HSI的空间分布。应用R3.3.2软件绘制海州湾长蛇鲻的HSI分布。
3 结果
3.1 适应性曲线和因子权重
本研究比较了5种环境因子和饵料因子的组合,分别为A:底层水温、底层盐度、水深、枪乌贼丰度;B:底层水温、底层盐度、水深、戴氏赤虾丰度;C:底层水温、底层盐度、水深、六丝钝尾鰕虎鱼丰度;D:底层水温、底层盐度、水深、3种饵料丰度之和;E:底层水温、底层盐度、水深、3种饵料丰度之和的对数。分别对其进行交叉验证,得出均值置信区间的R2(Determination Coefficient )值和 AIC 值。结果显示(表1),E组合的模型具有较低的AIC值,且R2值最接近1,拟合效果较好,因此本文选择E组合构建HSI模型。
根据适应性曲线分析可知(图2),海州湾长蛇鲻主要分布在底层水温17.5~26.6℃,底层盐度15~32,水深2~40 m的海域。最适底层水温范围为17.5~18℃,最适底层盐度范围为31.3~32,最适水深范围为24~37 m,最适的3种饵料生物丰度之和的对数范围为 13~14(图 2)。
根据BRT模型的分析结果,对长蛇鲻总偏差贡献率最大的是饵料因子,贡献率为96.05%,其次为水深(3.26%)、底层水温(0.62%)和底层盐度(0.07%)(图 3)。

表1 基于AMM算法和GMM算法的海州湾长蛇鲻HSI模型的交叉验证Table1 Cross-validation of HSI model of the Haizhou Bay Saurida elongata based on AMM and GMM algorithm in mean confidence interval
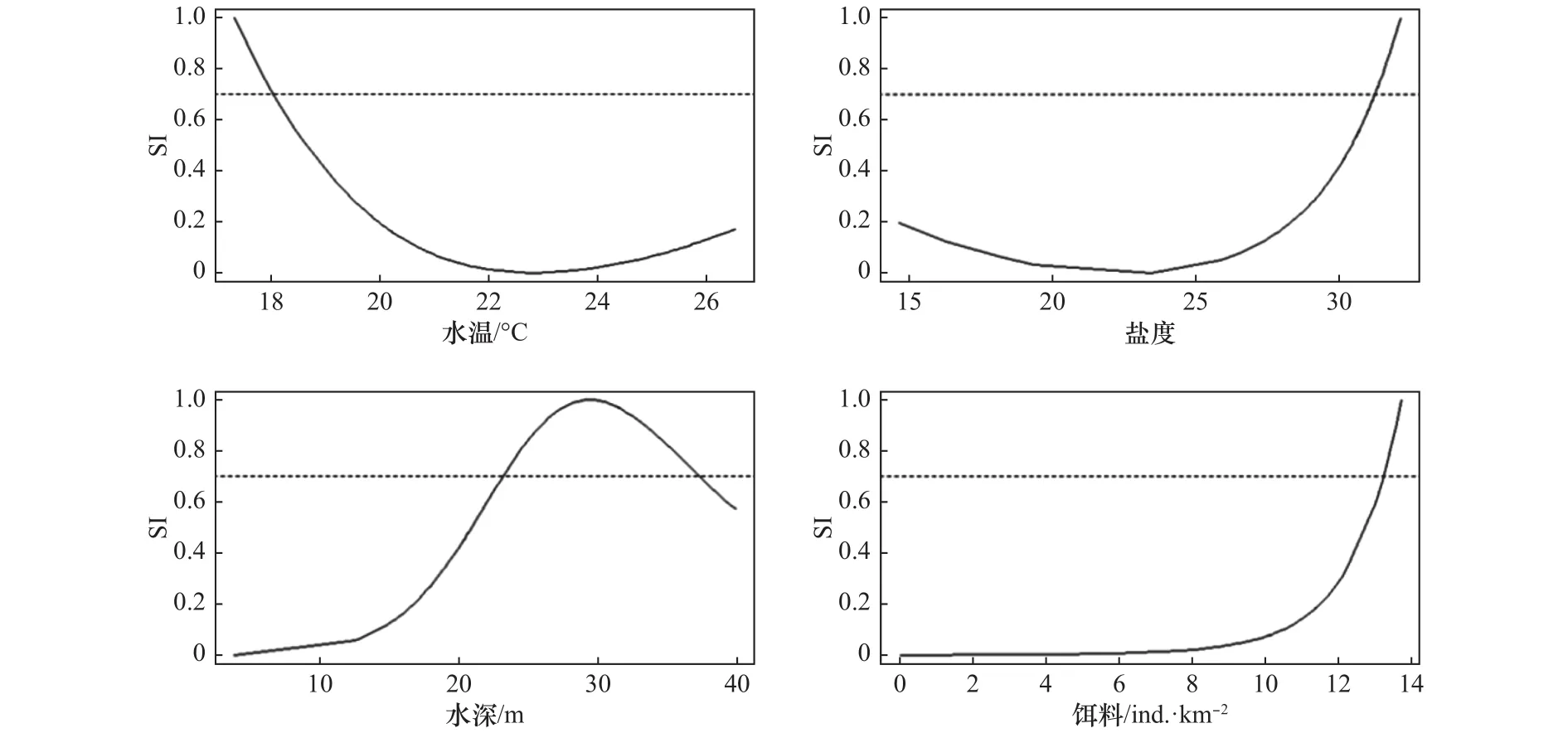
图2 海州湾长蛇鲻对底层水温、底层盐度、水深和饵料因子的适宜性指数曲线Fig.2 Suitability index curves of bottom temperature, bottom salinity, water depth and prey factors for Saurida elongate in the Haizhou Bay
3.2 AMM模型和GMM模型的比较
采用交叉验证的方法评估AMM模型和GMM模型的预测性能,得出均值置信区间的R2值和AIC值。结果显示(表2),运用AMM算法,并赋予权重的HSI模型AIC值较低,且R2值最接近1,拟合效果较好,故选取AMM算法且赋予权重的HSI模型研究长蛇鲻的栖息地适宜性。
3.3 海州湾长蛇鲻的最适栖息地分布
海州湾2011年和2013-2018年秋季长蛇鲻的最适栖息地(HSI≥0.7)主要分布在 34.5°~36°N,119°~121°E 之间,其中 35°~36°N 海域的最适栖息地分布范围大,从近岸至远海,适宜指数有增加的趋势(图4)。2011年,长蛇鲻的最适栖息地主要分布在近岸,集中于 35°~35.5°N,119°~120°E 海域;2013 年,其适宜栖息地分布范围较2011年明显缩小,最适栖息地区域仅占研究海域的很小一部分;2014年,长蛇鲻的最适栖息地分布范围有明显扩大,在 34°~36°N,119.5°~121.5°E 海域都有分布,尤其在 35°~36°N,120.5°~121.5°E 海域,HSI值可达到 1.0。2015年其 HSI值较2014年有所减小,在0.6~0.8之间,但分布范围相似。2016年最适栖息地向北偏移,主要分布在35°~36°N,119.5°~120.5°E 之间。2017 年长蛇鲻最适栖息地范围进一步缩小,只集中于 35.5°~36°N,120°~121°E的海域中。2018年其最适栖息地分布范围较2017 年明显缩小,主要分布在 35°~35.5°N,120°~120.5°E 海域(图 4)。

图3 各个非生物因子和生物因子对海州湾长蛇鲻提升回归树模型总偏差的相对贡献Fig.3 Relative contribution of different abiotic and biotic factors to the total deviation explained by the boosted regression tree models of Saurida elongata in Haizhou Bay

表2 AMM模型和GMM模型预测性能的比较Table2 Comparison of prediction performance of AMM and GMM
4 讨论
4.1 影响海州湾长蛇鲻栖息地适宜性的因素
本研究发现,除了水温、盐度、水深等非生物因子外,生物因子对长蛇鲻栖息地适宜性的影响也十分明显。BRT模型的计算结果显示,对海州湾长蛇鲻适宜栖息地影响最大的是饵料因子,贡献率为96.05%,其次为水深(3.26%)、底层水温(0.62%)和底层盐度(0.07%)。饵料生物通过影响鱼类的生长代谢、繁殖发育等,从而间接地影响鱼类的洄游分布和栖息地适宜性。海州湾长蛇鲻摄食的饵料生物有30余种,鱼类是其最主要的饵料类群,此外还摄食少量的虾类和头足类,其中优势饵料生物有枪乌贼、戴氏赤虾等[20]。本研究表明,饵料生物对长蛇鲻空间分布的影响,明显超过了水深及盐度,是最重要的影响因子。随着海水深度的变化,各种水文要素,特别是温度、盐度、水色、透明度、水体流向、流速等也会随之发生变化,从而会影响鱼类的分布[27]。盐度的变化是支配鱼类行为的另一个重要因素,海水盐度的变化能够直接影响鱼类的渗透压调节,并通过水团、海流等间接影响鱼类的行为和栖息分布[27]。
海州湾长蛇鲻的产卵期主要集中在夏季,而秋季产卵活动结束以后,主要进行分散索饵活动,摄食强度较大,因此秋季饵料生物对其空间分布的影响较为显著。长蛇鲻主要以小型鱼类为食,同时也摄食头足类和虾类,其对头足类的摄食比例随叉长出现变化。当其叉长在140~159 mm之间时,头足类的摄食比例达到最大,叉长达到180~259 mm之间时则主要摄食鱼类[20]。海州湾秋季枪乌贼的资源密度较高[28],故在其饵料生物中占据了优势地位,也成为影响其空间分布的主要饵料生物。
4.2 海州湾长蛇鲻适宜栖息地的分布特征

图4 2011年和2013-2018年秋季海州湾长蛇鲻的HSI分布Fig.4 Distribution of HSI for Saurida elongata during autumn of 2011 and 2013-2018 in the Haizhou Bay
本研究发现,海州湾秋季长蛇鲻主要分布在底层水温17.5~26.6℃,底层盐度15~32,水深2~40 m的海域。最适底层水温范围为17.5~18℃,最适底层盐度范围为31.3~32,最适水深范围为24~37 m,最适的3种饵料生物丰度之和的对数范围为13~14。从HSI分布可以看出,海州湾长蛇鲻最适栖息地的分布随年份有较大变化。长蛇鲻的最适栖息地(HSI≥0.7)主要分布 34.5°~36°N,119°~121°E 之间,其中 35°~36°N海域的最适栖息地范围相对较大。同其他年份相比,2013年秋季海州湾长蛇鲻的适宜分布区范围明显减小。根据2013年的气象资料显示,该年份春、夏季的降水量明显高于其他年份,使得径流量增大,导致该海域的盐度有所下降。同时,根据本研究同步调查数据,2013年10月海州湾底层水温为9.5~15.9℃,3种饵料的丰度较低,因而导致2013年秋季海州湾长蛇鲻最适栖息地范围缩小。从HSI分布来看,长蛇鲻在海州湾北部的最适栖息地范围大于海州湾南部,这是因为海州湾东南部平岛、车牛山岛一带,底质多礁砾,不适宜长蛇鲻分布,而北部海区多为泥砂底质,是长蛇鲻适宜的产卵场和索饵场。
本研究发现,海州湾秋季长蛇鲻栖息地适宜性从近岸至远海有增加趋势。长蛇鲻主要适宜栖息在远岸底层海域,通常栖息于水深20~100 m,底质为砂或泥砂的海区。海州湾位于苏北沿岸流和黄海暖流分支的综合作用区域,湾内丰富的饵料和适宜的环境为长蛇鲻提供了充足的食物保障和良好的育幼场所。远岸海域较高的水温和盐度,为长蛇鲻成体的生长和生殖发育提供了有利条件,因此栖息地适宜性较近岸高。
4.3 海州湾长蛇鲻HSI模型的构建
HSI模型的构建方法是影响栖息地适宜性指数模型预测结果的主要因素,不同的计算方法表现出不同的优势。其中,AMM的计算结果较为稳定[29],GMM对SI的极大值和极小值较敏感[30]。本研究根据交叉验证的结果选取最适合的模型,即AMM,使得预测结果最优化。此外,鱼类对每一种环境因子的响应是不相同的,因此在建立栖息地适宜性指数模型时需要对每一因子赋予权重。在以往的栖息地适宜性研究中,很少考虑饵料等生物因子的影响。本研究采用BRT模型对环境因子赋予权重,并在非生物因子的基础上增加饵料因子,分析不同因子对长蛇鲻栖息地适宜性的影响,能够较好地反映海州湾长蛇鲻的栖息地适宜性特征。
在构建栖息地适宜性指数模型时,除了本次研究所涉及的水温、盐度和水深以及饵料生物等影响因子以外,还有其他因子,如叶绿素、流速、底质类型、溶解氧、捕食者、种间竞争等也会对鱼类的空间分布和栖息地选择产生影响[31-33]。因此,在今后的研究中,需要综合考虑更多的环境因子,以便更好地反映鱼类栖息地分布与环境因子的关系。
猜你喜欢
文萃报·周五版(2022年15期)2022-04-21
读者(2022年8期)2022-04-07
工业设计(2022年1期)2022-02-09
河南水产(2020年1期)2020-06-29
汉语世界(The World of Chinese)(2018年3期)2018-10-22
扬子江诗刊(2016年2期)2016-11-25
中国新技术新产品(2015年3期)2015-07-19
新少年(2015年5期)2015-07-10
河北渔业(2014年3期)2014-07-08
舰船科学技术(2012年3期)2012-07-11
