
噬菌体Chy5的生物学和基因组学特征
2020-07-17 06:31江莉莎邬亭亭郭述良
中国人兽共患病学报 2020年6期
江莉莎,刘 平,邬亭亭,郭述良
结核病的传播在全球范围内愈演愈烈,特别是耐药结核菌的出现更加剧了疫情流行。噬菌体在结核病的诊断和治疗上已得到了多方面的应用。在结核病诊断方面,荧光报告噬菌体(LRP)已逐步应用于结核菌及其耐药性检测[1-3]。但国内研究均以现有的荧光报告噬菌体为基础做扩展探究,尚无自行构建荧光报告噬菌体的报道。现有的荧光报告噬菌体多以噬菌体TM4、D29、L5、Che12为基础。其中,利用噬菌体TM4、D29构建的荧光报告噬菌体会杀死宿主菌,而死菌无法发出荧光,导致其敏感度较低;L5、Che12因为其溶源性噬菌体的特性,其荧光报告噬菌体敏感度较高,但仍需104/mL结核菌才能检测到荧光[1,4]。另外上述噬菌体均已注册专利,对后续研究及专利申请不利。因此,不断筛选新的能感染结核分枝杆菌的溶源性噬菌体,并完成基因测序,仍是噬菌体研究者十分重要的基础工作,以期构建出敏感度更高的荧光报告噬菌体。
1 材料与方法
1.1菌株 耻垢分枝杆菌(MycobacteriumSmegmatis, MS)mc2155(CMCC 93202)由中国药品生物制品检定所王国治教授馈赠。结核分枝杆菌标准株H37Rv(CMCC93004)由重庆市肺科医院馈赠。分枝杆菌临床株36株由重医一院患者痰标本中分离得到,利用罗氏药敏培养基(珠海贝索公司)完成菌种和药敏鉴定。
1.2噬菌体的分离 取5 g土壤样本,用10 mL 噬菌体缓冲液浸泡30 min,使噬菌体充分进入缓冲液中,4 500 g 离心10 min,小心吸取上清液,0.22 μm滤器过滤除菌。取1 mL上清液与1 mL耻垢分枝杆菌混合,再加入8 mL 7H9液体培养基(BD,美国,含10% ADC),混合均匀,在恒温振荡器中(37 ℃,160 r/min)震荡培养过夜,离心收集上清液,并用0.22 μm过滤器过滤除菌。将100 μL的过滤液和1 mL的耻垢分枝杆菌菌液充分混匀后室温静置15 min,再加入3 mL 7H9固体培养基(BD,美国),混匀后平铺于7H10固体培养基(BD,美国)平板上。倒置培养24 h。收集到典型的噬菌斑后再纯化5代。根据噬菌体分离地点及样本序列号,将噬菌体命名为Chy5。
1.3噬菌体电镜观察 取20 μL电纯化的噬菌体液,滴于铜网上,静置15 min,用滤纸从侧面吸去多余的液体,再加20 μL 2%磷钨酸于铜网上,染色10 min,待其自然干燥,并在透射电镜(Hitachi-7500, 日本)下观察噬菌体形态。
1.4噬菌体宿主谱 利用单斑法[5]测定宿主谱。将稀释的各宿主菌(各结核临床株及标准株H37Rv)分别制成均匀的菌苔。再在各菌苔上滴加5 μL噬菌体样液,待其晾干后倒置于 37 ℃恒温箱中培养,每天观察结果至6周后。(阳性:有噬菌斑形成,阴性:无噬菌斑形成;阳性对照组为耻垢分枝杆菌。)
1.5基因组测序及生物信息学分析 用λ噬菌体DNA 提取试剂盒(北京艾根比公司),按操作说明提取噬菌体Chy5的 DNA。噬菌体DNA的全基因组测序采用鸟枪法由北京六合华大基因科技股份有限公司完成。得到的序列用Phred/PhraD/Consed软件包组装、拼接成若干个重叠群,各重叠群间的缺口通过设计PCR引物,扩增连接,得到完整的全基因组序列。用DNAStar软件包分析基因组组成成分;用Glimmer3.0、GeneMark软件对基因组序列作基因预测,所得结果互相补充。为完成噬菌体的基因组比较,将噬菌体基因与公共数据库基因作blast m8比对,e-value <= 1e-5,选取每个蛋白的最好比对结果,将两两比对均是最好的结果保留,然后用perl语言,生成svg的共线性图。
1.6噬菌体Chy5溶源菌的分离 参照Vanaja Kumar的实验方法[1],用无菌接种环蘸取Chy5噬菌斑中几乎清晰透明的区域,在一块新的7H10固体培养基平板上接种划线,倒置于37 ℃恒温箱中培养,直至某块平板上有菌落长出,即认定为Chy5溶源菌。
1.7紫外线诱导试验 参照Vanaja Kumar的实验方法[1],从7H10固体培养基上挑取Chy5溶源菌菌落,置于7H9液体培养基中重悬。取500 μL混悬菌液,在30 W的紫外灯(间距50 cm)下照射30 min,再和500 μL耻垢分枝杆菌菌液及2 mL 7H9固体培养基混合,平铺于7H10下层平板上冷却凝固,倒置于37 ℃恒温箱中培养24 h。以耻垢分枝杆菌为对照组重复上述实验。
1.8噬菌体超免反应 参照Vanaja Kumar的实验方法[1],将Chy5溶源菌与7H9固体培养基充分混匀,铺于7H10下层平板上冷却凝固,制成菌苔。分别取10 μL 10倍梯度稀释的Chy5、D29、Leo噬菌体液点涂于菌苔上,并标记斑点位置,倒置在37 ℃恒温箱中培养24 h。以耻垢分枝杆菌为对照组重复上述实验。
2 结 果
2.1噬菌体分离及噬菌斑形态 我们以耻垢分枝杆菌为宿主菌,在重庆医科大学附属第一医院结核病区分离得到噬菌体Chy5。噬菌体Chy5在最初24 h形成透明、圆形的噬菌斑,但继续培养至48 h,噬菌斑逐渐变浑浊。
2.2电镜观察 噬菌体Chy5由头部和尾部组成。头部为对称的多面体结构,直径(61.5±1.4) nm,尾部可弯曲、不可收缩,长度(114.1±2.1) nm。迄今发现的分枝杆菌噬菌体均为有尾噬菌体;其中大部分具有可弯曲、不可收缩的长尾,称为长尾噬菌体;小部分仅有不可收缩的短尾,称为肌尾噬菌体[6]。因此,噬菌体Chy5属于长尾噬菌体。
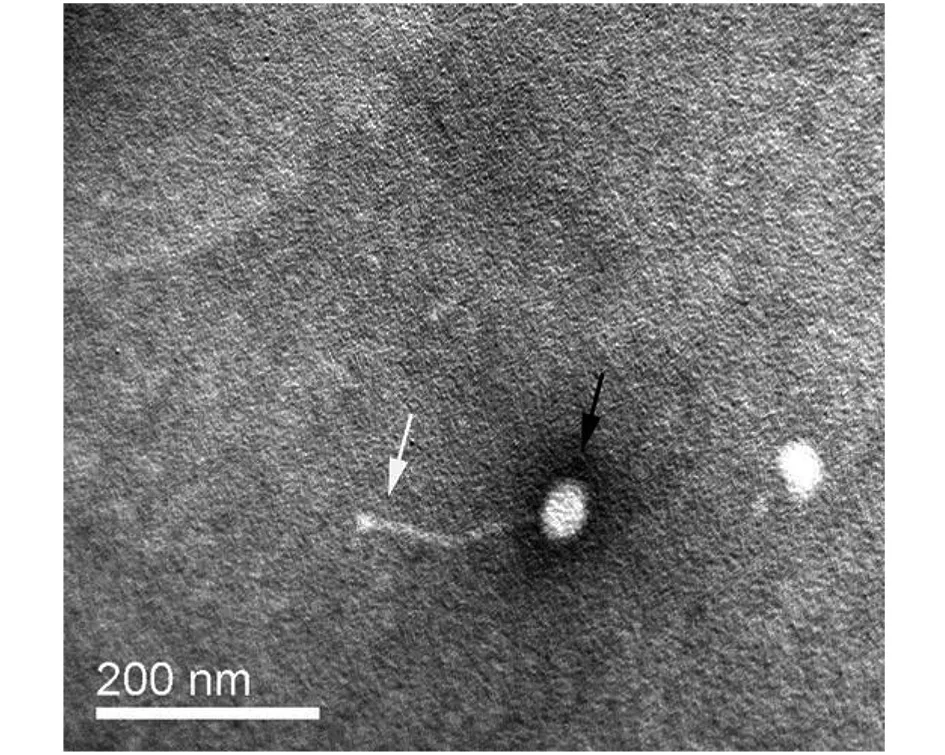
注:白色箭头指示噬菌体尾部,黑色箭头指示噬菌体头部。图1 噬菌体Chy5电镜图(×60 000)Fig.1 Electronmicrogragh of phage Chy5
2.3噬菌体宿主谱 噬菌体Chy5可感染结核菌标准株H37Rv,对临床结核敏感菌感染率可达100%。除广泛耐药菌外,对耐多药及多耐药结核菌的感染率在85%以上。同时,噬菌体Chy5还能感染部分牛结核分枝杆菌及非结核分枝杆菌,见表1。
2.4 噬菌体基因组测序及生物信息学分析
2.4.1基因组组成成分分析 Chy5基因组全长51 214 bp,G+C含量63.60%,有3种起始密码子,分别是ATG (n=52), GTG (n=27) 和TTG (n=10)。
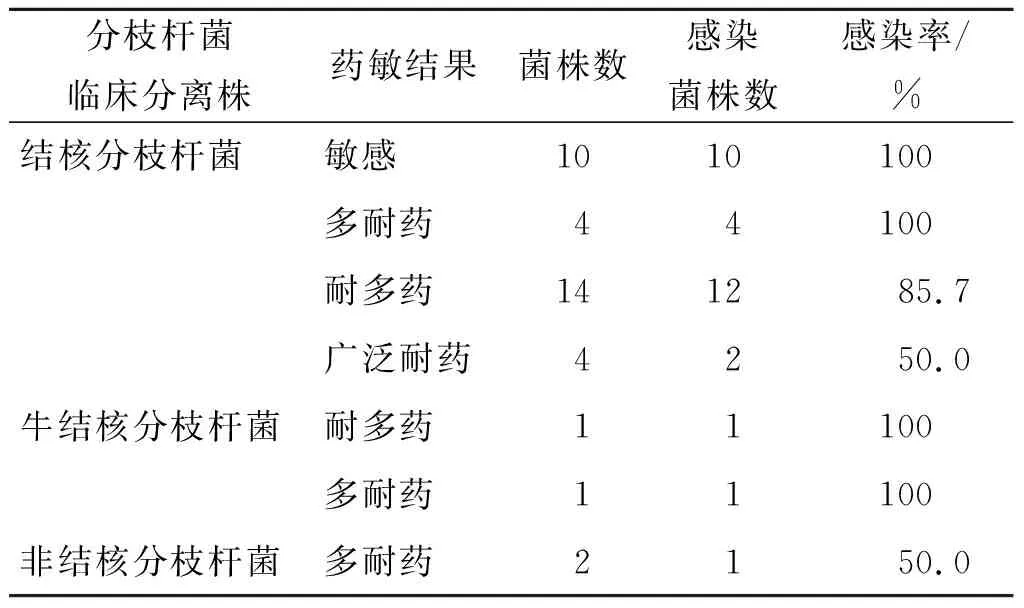
表1 噬菌体Chy5宿主谱Tab.1 Host range of phage Chy5
2.4.2基因组功能预测分析 经软件预测,Chy5有88个推定基因,大部分与噬菌体D29所对应的基因相似,其中有30个基因有推测功能,如表2。按基因功能,主要分为3大类,裂解元件、整合元件以及病毒结构和聚集基因。
噬菌体Chy5的裂解元件由裂解酶A、裂解酶B和Holin蛋白组成,其中ORF 6与D29的裂解酶A相似度99.8%,ORF 7与D29的Holin蛋白相似度100%,ORF 8与D29的裂解酶B相似度99.21%。
噬菌体需将其基因组整合到宿主菌的染色体中,才能产生溶源现象[7]。据溶源性噬菌体L5的研究表明,attP位点和整合酶在此过程是必不可少的[7-8]。除此之外,要维持溶源现象的稳定,基因组中必须存在有活性的阻遏蛋白[9]。Chy5 ORF 31与D29的整合酶相似度82.58%,推定其编码整合酶。attP位点通常位于整合酶的下游,且与宿主菌tRNA中的某一段(attB位点)重合[10]。因此,利用NCBI的BLASTN功能,对ORF 31(即整合酶)下游区域和结核分枝杆菌H37Rv基因组作比较,发现一段核酸序列(26973-27013bp)与H37RV基因组中tRNA-Gly基因中某片段(2765541-2765611bp)完全重合,即为attP位点,如图2。另外,ORF 70推定为编码阻遏蛋白的基因,但Chy5 ORF 70与噬菌体Pukovnik的阻遏蛋白相似度仅70.82%,是否具有活性,尚待进一步实验证实。
病毒结构和聚集基因位于基因组的左臂,共13个。其中ORF 23编码卷尺蛋白,与噬菌体尾部长度相关[11]。

表2 噬菌体Chy5基因注释Tab.2 Gene annotation of phage Chy5

402974631569-60768.19ATGDNA polymerase413157731912-11112.11ATG423232232528-687.88ATG433191232322-13615.3ATGHTH_Hin_like443253133238-23526.53ATGRP thymidylate synthase453330633884-19220.33TTG463389635788-63070.04ATGribonucleotidereductase473597736174-657.37GTG483617136356-617.06ATG493704437196-506.06TTG503634037047-23526.41TTG513720037967-25528.55GTGphosphoesterase523796038415-15117.38GTG533841238684-9010.22ATGThioredoxin543868439316-21023.68TTGRP primase/helicase553953039949-13915.93GTGRecombination endonuclease VII563997840121-474.97ATG574011840954-27829.81ATGAbhydrolase/ Esterase_lipase584095141343-13014.58TTG594132841483-515.84GTG604148041716-788.86ATG614187942265-12813.99ATG624229742947-21623.52TTGP-loop_NTPase /RecA-like_NTPases634302743173-485.49GTG644317043769-19922.88GTG654376643906-465.31GTG664390344136-778.7ATG674417744419-809.46ATG684446745276-26930.86GTGRecB-like nuclease694527345692-13915.71ATG704574846242-16419.08ATGrepressor714648946647-525.85GTG724664446916-909.84ATG734714047274-444.93GTGnovel NTPase744691347143-768.76GTG754727447552-9210.19ATG764755248325-25728.94ATG774832848465-455.48ATG784847648679-677.2ATG表2(续)ORFStartStopStrandSize(aa)MW(kDa)Start codonPredicted Function794868948817-425.13GTG804881449080-889.59ATG

814911449476-12013.7ATGParB-like nuclease824950349883-12614.21ATG835000850121-374.13GTG844988050011-435.07GTG855010050294-646.89GTG865030950575-8810.09ATG875057850742-546.21GTG885075051202-15016.94ATG
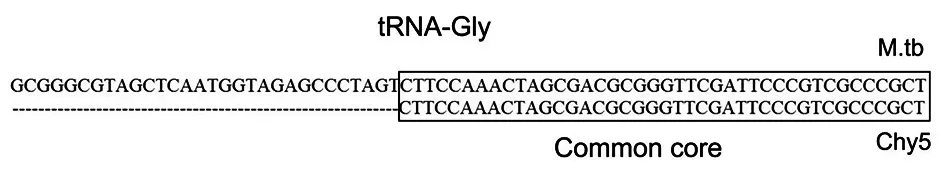
图2 噬菌体Chy5基因组的attP位点Fig.2 attP site of phage Chy5 genome
2.4.3共线性分析 与Chy5基因组最相似的噬菌体是D29 和Pukovnik,尤其是D29,如图3。各基因高度同源,共线性清晰,提示Chy5为A簇噬菌体。
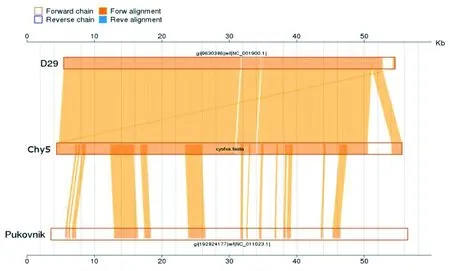
图3 噬菌体Chy5基因共线性分析Fig.3 Colinearity analysis of the genes of phage Chy5
2.5紫外线诱导试验 将分离得到的Chy5溶源菌在紫外线下照射30 min后,再将其与耻垢分枝杆菌共培养过夜,平板上可见噬菌斑形成。而以野生型耻垢分枝杆菌为对照组重复上述实验,则没有噬菌斑。上述结果表明Chy5为溶源性噬菌体,可形成溶源菌。
2.6超免反应 与对照组相比,Chy5几乎无法在Chy5溶源菌菌苔上形成噬菌斑;D29在高滴度时可在Chy5溶源菌菌苔上形成噬菌斑,但在低滴度时亦无法形成清晰的噬菌斑。而Leo在任何滴度下均可在Chy5溶源菌及耻垢分枝杆菌菌苔上呈现清晰的噬菌斑(图4)。

注:噬菌体滴度从左到右依次为107、106、105、104 PFU/mL图4 噬菌体Chy5超免反应Fig.4 Superinfection immunity of phage Chy5
3 讨 论
本实验分离到一株噬菌体,命名为Chy5。其噬菌斑在48 h后变浑浊,提示其可能为溶源性噬菌体。电镜观察噬菌体颗粒形态,见其尾部可弯曲、不可收缩,长度(114.1±2.1) nm,属于长尾噬菌体科。通过宿主谱测定,发现噬菌体Chy5宿主谱广,不仅能感染结核标准株和结核临床敏感株,还能感染绝大多数结核临床耐药株,有望应用于荧光报告噬菌体构建,故进一步完成基因测序工作,了解其遗传信息。
Chy5基因组全长51 214 bp,G+C含量63.60%,同大多数A簇噬菌体相同,但低于其宿主菌耻垢分枝杆菌(67.4%) 和结核分枝杆菌(65.6%)[12]。据共线性分析结果,与Chy5基因组最相似的噬菌体是D29,各基因高度同源,提示其为A簇噬菌体。
经过基因功能预测分析,证实Chy5基因组中含有编码整合酶和阻遏蛋白的基因及attP位点。然而,尽管具有整合酶及attP位点,D29仍然是裂解性噬菌体,这是由于其阻遏蛋白基因上一个3.5 kb长的片段的缺失所致[13]。Donnelly-W等证实,当溶源性噬菌体L5的阻遏蛋白部分或全部缺失后,其亦无法形成溶源菌[9]。因此,具有活性的阻遏蛋白是维持噬菌体溶源性的必备条件。Chy5 ORF 70与噬菌体Pukovnik的阻遏蛋白相似度仅70.82%,尚无法确定其阻遏蛋白活性。但进一步行紫外线诱导试验发现,经紫外线照射,可从分离所得的Chy5溶源菌中诱导出噬菌体,证实Chy5确为溶源性噬菌体。另外,溶源性噬菌体无法裂解自身溶源菌的现象,称为超免反应;这是由于噬菌体与溶源菌中相同的基因组相互免疫导致的[7,14]。而相似的基因组之间也会不同程度的发生这种现象[14]。实验发现,噬菌体Chy5几乎无法在Chy5溶源菌菌苔上形成噬菌斑,低滴度的噬菌体D29亦无法在Chy5溶源菌菌苔上形成清晰的噬菌斑,提示发生超免反应。
上述实验均证实Chy5为溶源性噬菌体,然而Chy5 ORF 70与Pukovnik的阻遏蛋白存在差异,提示两者共存某些重要元件使Chy5阻遏蛋白的活性得以保持。有研究表明,L5阻遏蛋白能保持溶源性稳定与其含有helix-turn-helix (HTH)DNA 结构域相关[9,15]。利用NCBI的CD-search功能,搜索噬菌体Chy5和Pukovnik的阻遏蛋白序列中的保守结构域,发现均存在HTH DNA 结构域,这可能是Chy5 阻遏蛋白能保持活性的原因。
因此,噬菌体Chy5是一株可感染结核菌的溶源性噬菌体,遗传背景清楚,有望应用于结核分枝杆菌鉴定及耐药菌检测。
利益冲突:无
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
科学24小时(2020年4期)2020-05-14
中外医学研究(2017年6期)2017-03-30
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
