
镉胁迫对大青杨不同倍体的生长及生理生化的影响
2020-07-14 01:00宋子文刘焕臻马晓雨孙国语易嘉欣张春华
植物研究 2020年5期
宋子文 刘焕臻 马晓雨 孙国语 易嘉欣 张春华
尤远祥2 王德秋3 李开隆1*
(1.东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040; 2.黑龙江省富锦县林业局,富锦 156100; 3.吉林省临江林业局,临江 134600)
在重金属污染的环境下,植物为减少其对自身生存造成的危害,通过增加细胞的渗透势、功能蛋白的数量和增加抗氧化酶数量来维持细胞的正常代谢,从而提高植物的抗性[1]。近年来,对重金属胁迫条件下的林木生长、叶绿素、生理生化特征的研究越来越深入[2]。叶绿素是一类与光合作用(photosynthesis)有关的最重要的色素。叶绿素含量的变化会影响植物光合作用,可能直接影响光合产物的形成和积累,影响植物生长发育和抵抗逆境的能力[3]。植物在高浓度重金属情况下生长,细胞膜受到破坏,质膜通透性增强,细胞内的物质外渗[4~5],致使植物代谢功能紊乱甚至丧失代谢功能,植物通过增加脯氨酸等渗透调节物质来维持细胞的代谢提高自身的抗性[6]。多倍体细胞核内染色体组加倍后,会带来生长、生理变化,植物高度,抗逆性增强等特点[7]。多倍体的某些代谢产物比二倍体多,在生长条件一致下,9年生三倍体毛白杨的材积要比二倍体高2~3倍[8];徐剑秋等[9]研究了干旱胁迫和盐胁迫下,大青杨三倍体、四倍体叶绿素含量高于二倍体;周珊珊等[10]研究表明在低温和高温胁迫下,大花蕙兰类原球茎三倍体、四倍体的MDA含量均显著低于二倍体。三倍体或四倍体的日本黑松与二倍体相比含有较高的SOD可以明显防止脂质过氧化物的形成和氧化损伤[11]。
大青杨(PopulusussuriensisKom.)是东北林区特有的乡土速生树种,也是恢复森林生态环境、营造生态林及短周期工业用材林最理想的树种之一。目前对大青杨的研究主要集中在干旱胁迫[12]、低温胁迫[13]、耐盐分析[14]、转基因表达谱分析[15]等方面。对大青杨多倍体在抗旱、抗盐等方面有过研究[9],而对于其在重金属胁迫中的情况鲜有研究。本研究利用不同浓度的CdCl2处理大青杨多倍体扦插苗,对其在重金属镉胁迫下的生长、细胞膜透性、光合作用、抗氧化物酶、细胞的渗透调节物质等进行分析,以期探究大青杨多倍体对重金属胁迫的生长及生理响应机制,为大青杨多倍体的选育和推广种植奠定理论基础。
1 材料与方法
1.1 试验材料
大青杨三倍体和四倍体株系是东北林业大学林木遗传育种国家重点实验室创制的,2019年5月中旬,在东北林业大学实验基地塑料大棚,将1年生的大青杨不同倍体的穗条扦插育苗。每个营养杯中扦插一株,营养杯中的扦插基质配比为草炭土和沙子4∶1。扦插苗在相对湿度45%~65%、温度(日/夜)26℃~32℃/20℃~25℃条件下培养,74 d后(8月4日)选择长势一致的幼苗进行后续试验。
1.2 试验方法
CdCl2浓度梯度为50、100、200、400 mg·L-1,每个处理设20次重复。CdCl2水溶液每3 d浇灌1次,以二倍体、三倍体、四倍体0 mg·L-1CdCl2为对照。在CdCl2胁迫处理4、7、10、13、16 d后,取样测量生长、含水量、叶绿素丙二醛(MDA)超氧化物歧化酶(SOD)、过氧化氢酶(CAT)。胁迫处理5、10、15 d后取样测定叶片的叶绿素含量,每次取样时间均为上午8:30~9:30,每个单株选取第3~5片功能叶,并设置3次生物学重复。取样时避免机械损伤,将取下的叶片迅速放入液氮中速冻,保证叶片内的物质没有呼吸代谢,然后放在超低温冰箱中待测。
丙二醛(MDA)测定采用硫代巴比妥氨酸法、超氧化物歧化酶(SOD)活性采用NBT光化还原法测定、过氧化氢酶(CAT)活性采用紫外吸收法测定[16]、叶绿素含量测定采用丙酮和酒精混合法[6],生长指标(苗高、地径)的测定和叶片含水量的测定参照王学奎[17]的方法进行。
1.3 统计分析
采用Excel进行数据的录入,用SPSS 22.0统计软件进行单因素方差分析和多重比较(Duncan法),用GraphPad Prism8软件进行作图。
2 结果与分析
2.1 不同浓度重金属镉对多倍体生长影响
由表1可知,大青杨3种倍体随着胁迫浓度增加,树高和地径相对百分率下降趋势,二倍体、三倍体、四倍体与0 mg·L-1相比,树高和地径在400 mg·L-1显著下降(P<0.05),树高分别下降41.52%、19.42%、25.43%,地径下降34.13%、34.19%、32.17%。随着胁迫浓度的增加3种倍体的相对百分率均升高,相对百分率越高影响越大,所以影响顺序:二倍体>四倍体>三倍体,从生长指标看,大青杨不同倍体扦插苗的苗高和地径的生长对Cd的反应敏感,三倍体受胁迫影响较小,所以三倍体抗Cd胁迫能力最强。
表1 不同浓度重金属镉胁迫第13天时大青杨不同倍体生长比较
Table 1 Comparison of different ploidy growth ofP.ussuriensisunder different concentrations of heavy metal cadmium stress on the 13th day

系号Series处理Deal with(mg·L-1)株高Plant height(cm)相对百分率Relative percentage(%)地径Ground diameter(mm)相对百分率Relative percentage(%)二倍体Diploid0.0144.14±12.12a0.08.32±0.12a0.0二倍体Diploid50138.37±4.58a4.07.51±0.45b9.7二倍体Diploid100116.61±5.13b19.17.17±0.49bc13.8二倍体Diploid200100.33±5.77c30.46.61±0.27c20.6二倍体Diploid40084.3±4.16d41.55.48±0.61d34.1三倍体Triploid0167.13±3.61a0.09.71±0.25a0.0三倍体Triploid50166.34±5.29a0.58.51±0.18b12.4三倍体Triploid100157.69±5.02b5.67.46±0.29c23.1三倍体Triploid200147.67±3.21c11.67.01±0.21c26.9三倍体Triploid400134.67±7.09d19.46.39±0.35d34.2四倍体Tetraploid086.67±3.12a0.06.87±0.11a0.0四倍体Tetraploid5079.33±7.02b8.46.42±0.31b6.1四倍体Tetraploid10077.33±1.53b10.85.95±0.19c13.4四倍体Tetraploid20073.33±2.08c15.45.62±0.16c18.2四倍体Tetraploid40064.63±2.01d25.44.66±0.31d32.2
注:相对百分率=(0 mg·L-1生长值-处理生长值)/0 mg·L-1生长值×100% 不同字母表示差异显著性 下同
Note:Relative percentage=(0 mg·L-1growth value-treatment growth value)/0 mg·L-1growth value×100% Different letters in the table indicate significant difference among different drought stress conditions The same as below
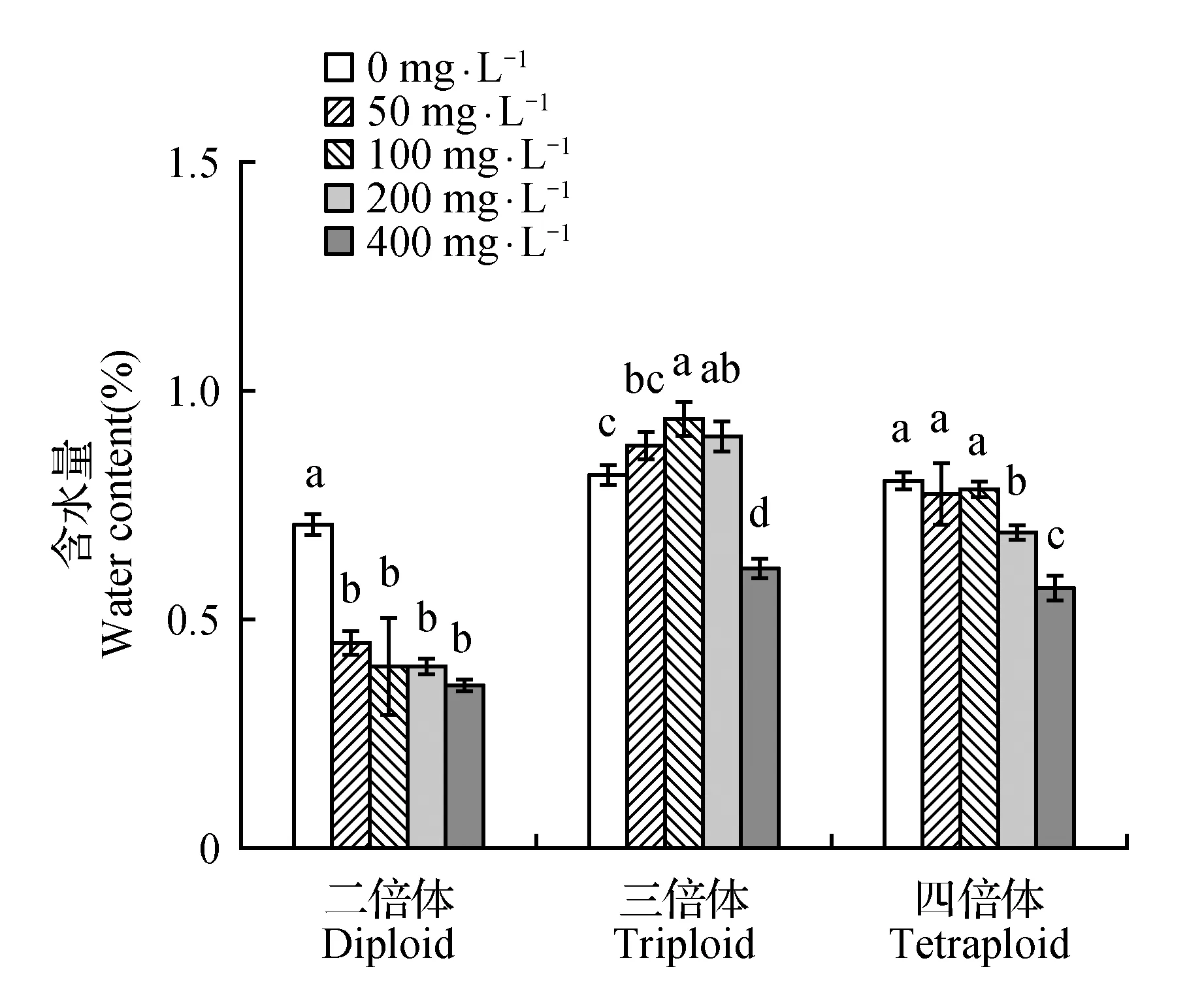
图1 不同浓度重金属镉胁迫第13天对大青杨不同倍体含水量影响 图中不同字母表示差异显著性 下同Fig.1 Effects of different concentrations of heavy metal cadmium on the 13 d on different ploidy water content of P.ussuriensis Different letters in the figure indicate significant difference among different drought stress conditionsThe same as below
2.2 不同浓度重金属镉胁迫对大青杨多倍体叶片含水量的影响
如图1所示,Cd处理对大青杨3种倍体的水分代谢有显著影响。随着镉浓度的增加,二倍体和四倍体的叶片含水量均呈下降趋势,二倍体在0 mg·L-1时最高,200和400 mg·L-1时显著下降,分别下降28.91%、34.1%(P<0.05),其他的略微下降,三倍体呈先上升再下降的趋势,三倍体在100 mg·L-1时显著增长,比0 mg·L-1时增长13.12%(P<0.05)。四倍体在0 mg·L-1时最高,400 mg·L-1时显著下降,下降了29.12%。
2.3 不同浓度重金属镉胁迫对大青杨多倍体叶绿素含量的影响
如图2所示,随着浓度的增加,二倍体、三倍体、四倍体均呈先上升后下降的趋势,3种倍体的表现程度有所不同。二倍体随浓度增加,100 mg·L-1时显著上升(P<0.05),比0 mg·L-1时增长了30.37%,三倍体在随着浓度增加,叶绿素含量均比对照高,100 mg·L-1时叶绿素含量显著增加(P<0.05),比0 mg·L-1时增加43.86%,四倍体在100 mg·L-1时叶绿素含量显著增加(P<0.05),比0 mg·L-1时增加了41.92%。

图2 不同浓度重金属镉胁迫第13天对大青杨不同倍体叶绿素含量影响Fig.2 Effects of heavy metal cadmium stress on different day 13 d on chlorophyll content in different ploidy of P.ussuriensis

图3 不同浓度重金属镉第13天对大青杨不同倍体MDA含量影响Fig.3 Effects of different concentrations of heavy metal cadmium on the 13 d on MDA content in different ploidy of P.ussuriensis

图4 不同浓度重金属镉第13天对大青杨不同倍体CAT活性影响Fig.4 Effect of heavy metal cadmium at different concentrations on day 13 d on the content of CAT activities in different ploidy of P.ussuriensis

图5 不同浓度重金属镉第13天大青杨不同倍体SOD活性影响Fig.5 Effects of different concentrations of heavy metal cadmium on the 13 d on the SOD activities of different ploidy of P.ussuriensis
2.4 不同浓度重金属镉胁迫对大青杨多倍体叶片细胞膜透性的影响
如图3所示,随着胁迫浓度的增加,二倍体增加不显著(P>0.05);3种不同倍体的MDA的含量都是先增加后下降,随着浓度的增加,三倍体在200 mg·L-1时显著增加(P<0.05),增加了40.87%,四倍体随着浓度的增加,在200 mg·L-1时显著增加(P<0.05),增加了36.96%。说明三倍体的膜损伤最小,抗氧化能力最强,有较好的抗重金属镉的能力。
2.5 不同浓度重金属镉胁迫对大青杨多倍体叶片活性氧代谢的影响
如图4所示,不同浓度重金属镉胁迫下,二倍体随着浓度的增加酶活性有显著的下降;在400 mg·L-1时显著(P<0.05),比0 mg·L-1时下降了56.89%,三倍体先上升后下降,在200 mg·L-1时显著增加(P<0.05),比0 mg·L-1时增加了41.44%,400 mg·L-1的时候显著下降,下降了28.97%;四倍体先上升再下降,酶活性略微增加(P>0.05),400 mg·L-1的时候抑制酶的活性显著下降(P<0.05),下降了36.19%,综合来看三倍体有相对较好的抗性。
如图5所示,随着浓度的增加,二倍体先增加然后下降、增加再下降,在50、200、400 mg·L-1时显著下降(P<0.05),下降37.13%、39.27%、41.25%,100 mg·L-1时略微下降(P>0.05);三倍体先上升再下降,三倍体随着浓度的增加,在100 mg·L-1时显著增加,增长28.6%(P<0.05);随着浓度的增加,其他的略微增加;四倍体先下降然后上升再下降。在50 mg·L-1的时候下降显著(P<0.05),下降13.28%。说明三倍体和四倍体比二倍体的抗氧化系统更强。
3 讨论
重金属镉胁迫影响了大青杨多倍体苗高和地径的正常生长,随着镉浓度的增加,大青杨不同倍体的株高和地径都显著下降。王志香等人[18]在研究Cu、Zn、Pd、Cd 4种重金属胁迫中发现,樟树、大头茶、红鳞蒲桃的株高和地径随着重金属浓度的增加呈现下降趋势,与本研究结果相同,说明高浓度重金属能抑制植物株高和地径的生长。本研究还发现,随着胁迫浓度的增加细胞膜的渗透性增加,对植株的伤害越来越大。二倍体和四倍体的含水量持续下降,而在三倍体中呈现先增加后下降的趋势,重金属对植物生长的抑制作用,可能是因为土壤中过量的重金属与土壤组分发生一系列物理和化学反应,导致土壤性质发生变化。余东等[19]对枇杷的研究发现,高浓度镉对枇杷根、茎、叶生物量和含水率均有显著的抑制作用,这也与本研究结果相似。重金属对植物的胁迫后,会影响细胞内叶绿体的合成,使叶绿素含量下降,不利于植物的光合作用[20~21]。低浓度的重金属会促进3种倍体叶绿素的合成,然而随重金属浓度的升高,超出植物的耐性与抗逆性,会造成植物叶绿素的降解,抑制其体内叶绿素的合成[6]。卢扬章[22]的研究说明格木在镉胁迫下,随着镉浓度的增加叶绿素的含量先增加后下降,与本研究结果相同。在正常生长条件下,植物体内活性氧维持着合成与分解之间的动态平衡。在环境发生改变时,植物体内活性氧代谢平衡遭到破坏,活性氧分子累积,造成植物体生理生化功能紊乱。为克服不良环境的影响,植物调整体内抗氧化酶活性来清除活性氧物质,维持正常的生理状态[23]。过氧化氢酶是植物中普遍存在的一种重要的氧化还原反应酶,酶活性的高低与抗逆性有着重要的关系,重金属镉胁迫下,植物细胞能利用抗氧化酶来调节氧化还原平衡,抗氧化系统越活跃则植物的抗性越强[24]。酶活性增加能增强植物在逆境下的抵抗力增强,所以酶活性可作为衡量植物抗性能力的指标之一[25]。万文生等人研究了土沉香幼苗在Cd胁迫下,随着Cd2+胁迫浓度的增加,其叶片中SOD出现不同程度的下降[25]。韦颖等人[26]研究了罗汉果在重金属镉胁迫下,随着Cd浓度的升高,SOD活性逐渐升高,而后又降低。高浓度Cd胁迫加重了叶片氧化程度并对罗汉果生长产生一定的抑制作用,而SOD活性的提高与维持是罗汉果耐Cd胁迫的物质基础之一。在本研究中,大青杨三倍体和四倍体的SOD活性均随Cd胁迫浓度的增加先增加后下降,而二倍体随着Cd浓度的增加而下降,与韦颖等人[26]的研究结果基本相同,说明镉胁迫下在不同树木中,抗氧化性有着类似的特性,显示出抗氧化酶系统对重金属胁迫产生了积极的响应。活性氧物质的增加破坏了细胞膜的透性,致使细胞内的代谢紊乱,而植物体内的抗氧化酶的增加能有效消除活性氧物质。过氧化氢酶(CAT)是抗氧化酶,催化过氧化氢产生氧气和水,其活性与植物的抗逆性有一定的关系。CAT酶反映了植物生理生化代谢及生长发育的情况。CAT含量越高,植物的抗性越好。本研究中,总的来说二倍体中CAT含量随着镉胁迫浓度的增加而下降,而三倍体和四倍体均呈现先升高后下降的趋势。这表明低浓度CdCl2刺激杂草植物提高POD活性以清除活性氧自由基,但氧化胁迫加剧时,过多的活性氧物质不能及时消解,导致细胞质膜受损,抗氧化物酶系统紊乱。丙二醛(MDA)是由于生物体器官衰老或在逆境下受到伤害,其作为过氧化产物能与蛋白质、核酸、氨基酸等活性物质交联,形成不溶性的化合物沉积,从而干扰细胞内正常的生命活动与透性。韦颖等人[27]通过盆栽试验,测定不同浓度Cd处理的罗汉果幼苗,MDA含量在相对低浓度Cd处理下MDA含量少于对照,随着Cd浓度的升高指标有显著升高,高浓度CdCl2胁迫加重了叶片氧化程度,并对罗汉果生长有一定的抑制作用。但本研究中3种倍体MDA含量随着镉浓度的增加均呈现先上升后下降的趋势。说明细胞MDA含量能反映出细胞的受伤害程度,重金属胁迫作用下,大青杨二倍体、三倍体、四倍体生长指标增长趋势相同,均是先增加后下降。低镉浓度下促进叶绿素的形成,在100 mg·L-1时达到最高点。在高浓度下,三倍体的抗性远高于其他两种倍体,导致植物叶绿素含量降低,Cd2+重金属离子对高等植物叶绿体的光合电子传递有抑制作用,直接干扰了叶绿素的生物合成,从而减少了植物对水分和养分的吸收,影响了植物正常的生长发育。细胞膜透性方面三倍体四倍含量均高于二倍体,处理后导致细胞质膜透性增加可能是由于与膜蛋白的结合或与磷酸酞胺乙醇胺和单分子层的磷脂丝氨酸反应,从而造成膜脂结构改变,致使MDA含量下降。活性氧代谢方面,随着镉浓度的增加,大青杨三倍体和四倍体的SOD值先增加后下降,二倍体处于下降趋势。活性氧物质的增加越多,对细胞膜的透性破坏越严重,所以植物体内产生更多的抗氧化酶来维持细胞的代谢。
总体来讲,在重金属镉的5种浓度梯度胁迫下,大青杨多倍体幼苗在生长和生理生化方面有着显著差异,综合所有指标来看,在低浓度CdCl2的时候大青杨3种倍体有较好的耐受性,而在高浓度CdCl2胁迫下大青杨三倍体在含水量、叶绿素含量、CAT活性、MDA含量方面比四倍体分别高5.47%、25.47%、8.59%、28.47%,比二倍体分别高23.47%、44.63%、17.23%、31.48%,表现出较好的抗性。
猜你喜欢
淡水渔业(2022年5期)2022-10-01
西北农业学报(2022年5期)2022-06-07
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
东北林业大学学报(2021年12期)2021-02-10
热带农业科技(2020年3期)2020-12-15
食品与生活(2019年12期)2019-05-15
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
江苏农业科学(2014年9期)2014-11-15
