
NBA1/MERIT40在3种沙棘中的适应性分化研究
——兼论二代测序技术在杂种鉴定中的应用
2020-07-14 01:00丁雪洋李雪丽刘本立
植物研究 2020年5期
孙 坤 丁雪洋 张 辉 李雪丽 汪 颖 王 娟 刘本立
(西北师范大学生命科学学院,兰州 730070)
沙棘属(HippophaeL.)广泛分布于欧亚大陆温带地区,全球共有7种11亚种,其中我国具7种7亚种,分为有皮组和无皮组[1~2]。无皮组代表中国沙棘(Hippophaerhamnoidesssp.sinensis)为较原始的种群,一般分布在海拔3 000米以下的地区[3]。肋果沙棘(H.neurocarpa)为青藏高原隆起过程中形成的适应更寒冷生态环境的有皮组一员,其生长于海拔3 400~4 300 m的地区[4~5]。棱果沙棘(H.goniocarpa)总是分布在肋果沙棘与中国沙棘同域分布的地区。形态研究及分子证据表明棱果沙棘为中国沙棘与肋果沙棘同倍化杂交形成,且是双向杂交[6~7]。海拔导致的不同生态环境,使得生存于此的中国沙棘与肋果沙棘在表型上已有较大差异,如肋果沙棘已经进化出适应高海拔生境、较中国沙棘植株更矮小,叶片更小更厚等特征[8~9]。棱果沙棘表型大多呈中国沙棘与肋果沙棘的居间性质。但目前对三者的生态适应性研究大多数仅停留在表型层面,鲜少有人在基因层面研究不同海拔下三者适应性分化的分子机制。
BRCA1(breast cancer suppressor gene 1)是位于17号染色体上与乳腺癌和卵巢癌有关的抑癌基因,主要通过修复受损DNA及控制细胞周期检查点等来维持基因组的稳定。BRCA1突变会提高患遗传性乳腺癌与卵巢癌的几率[10~11]。BRCA1的C末端包含两个保守的BRCT结构域,该结构域结合Abraxas/CCDC98(coiled-coil domain-containing protein)[12~13]、RAP80(receptor associated protein80/ubiquitin interaction motif containing 1)[14]、BRCC36(BRCA1-BRCA2-containing complex subunit 3)、BRCC45(BRCA1-BRCA2-containing complex subunit 45)/BRE(brain and reproductive organs)[15~16]、NBA1(new component of the BRCA1-A complex)/MERIT40(mediator of RAP80 interactions and targeting 40 kDa)[17~18]等亚基组成BRCA1-A复合体(breast cancer type 1 susceptibility pro-tein);该复合体位于细胞核中,主要功能是募集BRCA1至双链断裂处、控制G2/M检查点、去泛素化Lys-63泛素链信号、防止过度切除DSB(double-stranded DNA breaks)末端[19],这些功能由每个亚基之间相互协作完成。Abraxas亚基通过其磷酸化的SXXF基序直接与BRCT域结合,也招募具UIM(ubiquitin-interacting motif)域的RAP80亚基结合至BRCT域[20]。BRCC36为锌离子金属蛋白质酶体MPN+/JAMM家族的一个成员,能特异性地剪切K-63链接的多聚泛素链[21~22],其与细胞有丝分裂G2/M的检查点有关[21];也在减数分裂时期的姐妹染色单体互作中扮演着重要的角色[23]。NBA1为该复合体中重要的接头蛋白;与BRCC45亚基组成一个分子支架来维持该复合体的稳定[24~25];NBA亚基也是抵抗电离辐射所必需的,其与RAP80亚基相互作用募集BRCA1/BARD1 E3连接酶至DNA损伤位点,敲低NBA1导致细胞对化学试剂导致的DNA损伤、电离辐射更加敏感[23,26];NBA1具有介导蛋白酶体加工蛋白质的VWA结构域[27];与其同源的19S调节亚基Rpn10也具有VWA结构域,为募集修复因子至DNA损伤点所必需[28]。BRCC36、NBA1、BRE等亚基也参与形成位于细胞核及细胞质的BRISC复合体(BRCC36 isopeptidase complex)[20~21];该复合体主要在细胞核外行使功能,与有丝分裂纺锤体的组装、泛素化信号及病毒的蛋白降解有关[21,29];BRISC复合体在细胞质中具有多个底物,如细胞质中的代谢酶SHMT2(serine hydroxy methyltransferase 2)与其结合,导致二者的酶活性受到抑制。NBA1亚基在BRISC复合体与BRCA1-A复合体中的功能稍有不同,前者主要与泛素结合有关,后者不仅参与泛素化结合,也与RAP80亚基的整合有关[29]。目前对BRCA1-A&BRISC复合体的研究,主要集中在哺乳动物上。植物中对BRCA1-A&BRISC复合体研究较少。Block等人在拟南芥中发现了该复合体亚基BRCC36的同源蛋白质AtBRCC36A(At1g80210)及AtBRCC36B(At3g06820),二者与DNA的交叉连接修复有关[25]。
极端环境下生存的植物一定有其独特的适应机制[30]。我们推测肋果沙棘和中国沙棘中与低温、高辐射等相关的基因已经产生了一系列适应性分化,本研究筛选出中国沙棘、肋果沙棘与棱果沙棘RNA-seq数据中的功能基因NBA1,从该基因在三者体内的表达量与功能两方面来研究其在三者中的适应性分化。此外,随着测序技术不断发展,基于转录组或基因组序列鉴定杂交物种已得到广泛应用[31],但目前还未见文献评价第二代测序是否适用于鉴定杂交物种。本研究以NBA1基因为例,探讨了二代测序结果及在杂交物种鉴定中的应用。
1 材料与方法
1.1 材料
1.1.1 实验材料
采摘中国沙棘、肋果沙棘及棱果沙棘的嫩叶,随后放入液氮中迅速冷却。每种沙棘取3个个体重复。采样情况具体见表1。
1.1.2 数据来源
NCBI数据库(https://www.ncbi.nlm.nih.gov/);Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html);Piceaabies基因组官网(http://congenie.org/),TAIR数据库(https://www.arabidopsis.org/index.jsp)(见表2)。

表1 实验材料及采集地
注:标本均保存于西北师范大学生命科学学院标本馆
Note:The specimens stored in College of Life,Northwest Normal University
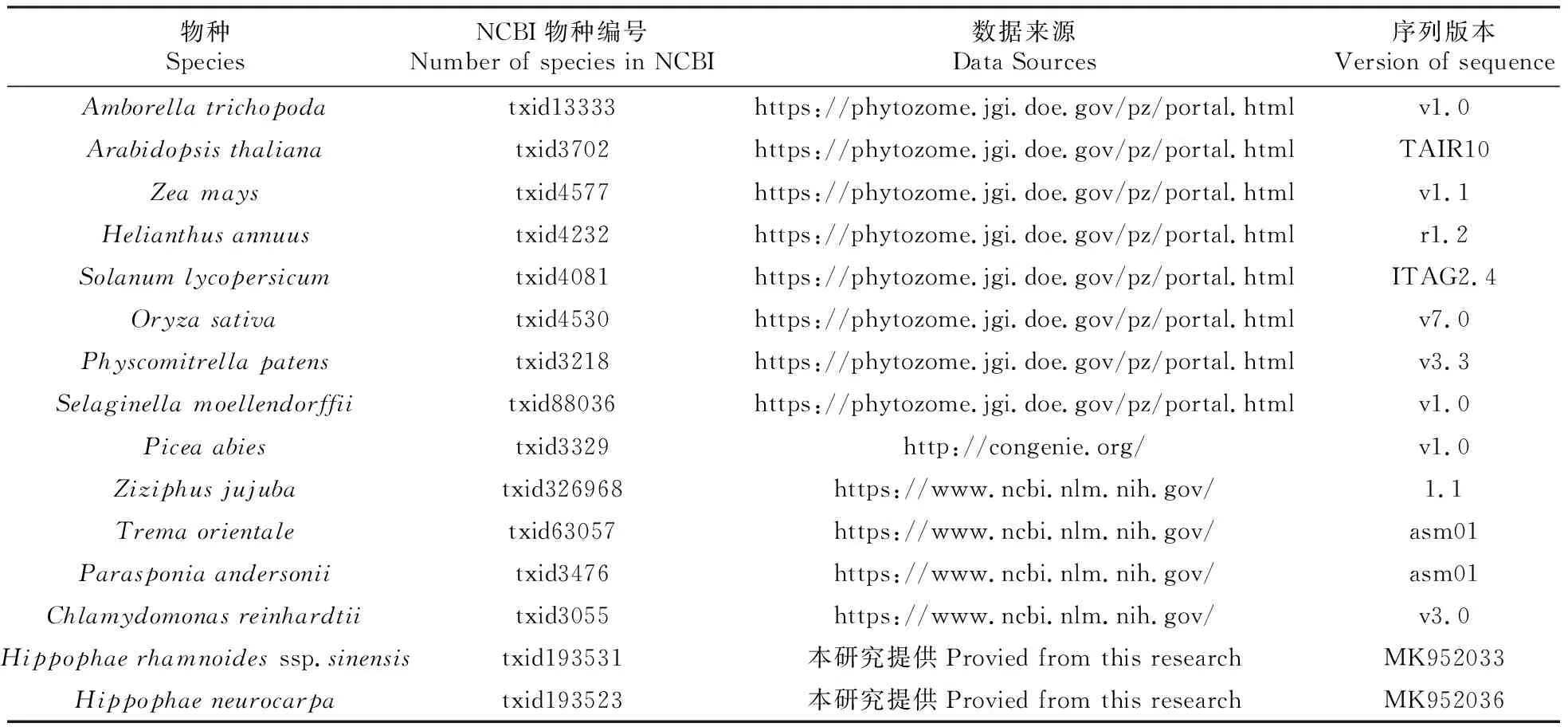
表2 15个物种蛋白质序列数据来源及版本
注:除中国沙棘和肋果沙棘外的其他13种植物选自植物系统进化树上关键节点
Note:The other 13 species exceptH.rhamnoidesssp.sinensisandHippophaeneurocarpaare selected from the key nodes of the phylogenetic tree of plants
表3 克隆NBA1基因所选用的个体及样本
Table 3 Experimental individuals and samples using cloningNBA1 gene
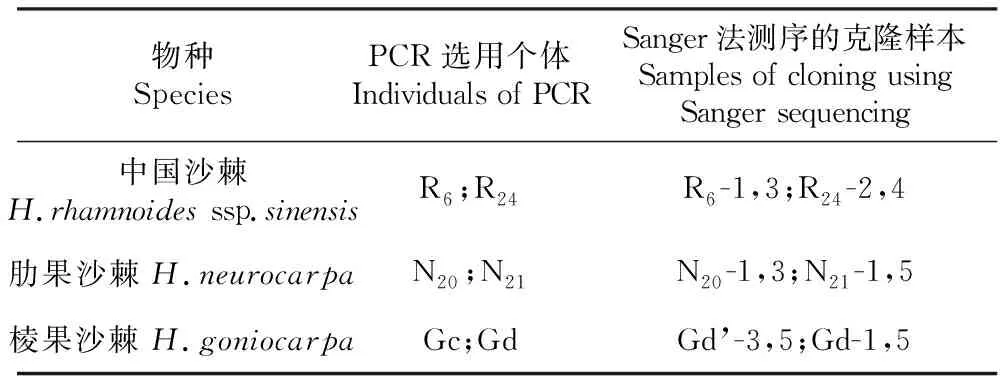
物种SpeciesPCR选用个体Individuals of PCRSanger法测序的克隆样本Samples of cloning using Sanger sequencing中国沙棘H.rhamnoides ssp.sinensisR6;R24R6-1,3;R24-2,4肋果沙棘H.neurocarpaN20;N21N20-1,3;N21-1,5棱果沙棘H.goniocarpaGc;GdGd’-3,5;Gd-1,5
注:表中的大写字母表示选用物种;R.中国沙棘;N.肋果沙棘;G.棱果沙棘;下标数字和小写字母表示选用的个体编号;“Sanger法测序的克隆样本”一栏中数字编号表示选用的克隆样本编号;Gd.所加引物为中国沙棘NBA1基因的引物;Gd’.所加引物为肋果沙棘NBA1基因的引物
Note:The capital letters in the table indicate the selected species; R.H.rhamnoidesssp.sinensis; N.H.neurocarpa; G.H.goniocarpa; in which the subscript numbers and lowercase letters indicate selected individuals’ number; The number in the “Samples of cloning using Sanger sequencing” column represent number of selected samples of cloning; Gd indicates that the primer isNBA1 gene ofH.rhamnoidesssp.sinensis; Gd’ indicates that the primer isNBA1 gene ofH.neurocarpa
1.2 方法
1.2.1 克隆NBA1基因
采用Qiagen试剂盒分别提取3种沙棘的总RNA,随后采用illumina平台的第二代测序技术对3种沙棘进行转录组测序;转录组数据存于西北师范大学生物信息学平台。每种沙棘随机选取2个个体利用Takara反转录试剂盒进行反转录后设计引物,用于目的基因NBA1扩增(见表3)。其中,中国沙棘和肋果沙棘的NBA1基因的引物序列分别为HrF:5′-AGAGAGAGAGAGGAGAGAGAG-3′,HrR:5′-ATCTAAAAATATATTCAAAATGTCCAG-3′和HnF:5′-GAGAGAGAGAGAGAAGAAGAGA-3′,HnR:5′-TAGAATCTGATAAGTCAATTAAACCA-3′,而杂交种棱果沙棘则分别利用中国沙棘和肋果沙棘的引物扩增目的基因。
PCR产物经Takara纯化试剂盒纯化后,使用PMDTM19-T载体进行克隆,选取条带清晰的克隆(见表3)进行Sanger法测序。
1.2.2 生物信息学分析
筛选正选择基因:基于OrthoMCL找出的直系同源基因对,以PAML中codeml程序下的“选择模型”计算dN/dS值(即w)。w>1即为受到正选择的基因[32~33]。
查找同源序列:利用Chromas软件分析Sanger法测序结果并导出[34];再利用NCBI网站BLASTX程序进行对比[35]。过滤条件为e值<10-6。
蛋白质结构分析:选择同源性最高且e值最小的BLAST结果,用DNASTAR软件中的Editseq及SeqBuilder程序截取目的基因的CDS区并翻译为氨基酸序列[36]。利用ProtParam工具(http://www.expasy.org/tools/protparam.html)分析蛋白质的分子量、等电点、亲水性等基本理化性质[37~38]。利用https://ch.embnet.org/software/TMPRED_form.html网站预测蛋白质的定位。蛋白质二级结构的预测使用Predictprotein(https://predictprotein.org/)[39~40]。将氨基酸序列提交至Zhanglab网站(https://zhanglab.ccmb.med.umich.edu/I-TASSER/)进行蛋白质三级结构预测[41]。用PyMOL软件分析蛋白质三维结构[42]。
直系同源基因的筛选:在Linux系统下进行本地BLAST来抽提13个物种中的同源蛋白质[43]。过滤条件:e<10-6。
多序列比对及进化树分析:利用ClustalX软件进行多重序列比对[44]。利用Prottest软件基于BIC算法计算14个物种中16条氨基酸序列最佳替代模型[45]。随后在MEGA中利用最大似然法选择最佳模型进行建树[46]。
2 结果与分析
2.1 Sanger测序验证杂交相关的3种沙棘中第二代测序结果的准确性
分析OrthoMCL软件的计算结果发现,NBA1基因在亲本种中国沙棘与肋果沙棘中为纯合的单拷贝基因,在杂种棱果沙棘的个体中以单个杂合位点的共显性方式存在。在中国沙棘R6号与R24号个体及肋果沙棘N20与N21号个体中均克隆出一条NBA1基因,而在二者的杂种棱果沙棘个体中同时克隆出两条NBA1基因。如图1所示,以Gd号棱果沙棘为例,Sanger测序显示该个体中同时存在中国沙棘与肋果沙棘的NBA1基因;二代测序的Gb及Gc号棱果个体与中国沙棘聚为一支,Ga及Gd号棱果个体与肋果沙棘聚为一支。这表明NBA1基因在亲本种中国沙棘与肋果沙棘中为单拷贝的纯合基因型,在杂种棱果沙棘中以共显性的杂合基因型存在。随后利用Clustal软件对Sanger法测序序列与二代测序序列进行多重序列比对,以验证亲本种中二代测序序列的准确性。结果表明,对中国沙棘而言,Sanger法测序与二代测序结果无差异。肋果沙棘N21个体利用Sanger法测出在CDS区的465位点碱基为T,二代测序为G;N16及N20个体Sanger法测序结果与二代测序结果均为G。在二代测序序列计算dN/dS值为1.112,dN/dS>1,即该基因在3个物种中受到达尔文正选择[33];基于Sanger测序序列(GeneBank号为MK952033,MK952034,MK952035,MK952036,MK952037,MK952038,MK952039)计算dN/dS值为2.560,dN/dS>1,表明该基因在3个物种中确实受到了达尔文正选择。
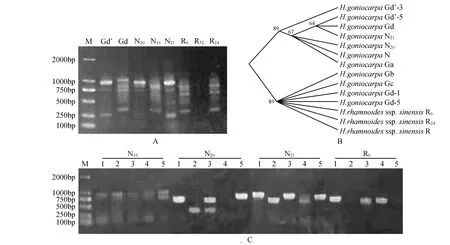
图1 NBA1基因克隆结果及Sanger测序与二代测序序列进化树 A.NBA1基因PCR结果;B.Sanger法测序与二代测序序列进化树(带下划线的为Sanger法测序序列);C.NBA1基因菌液PCR部分结果Fig.1 Cloning of the NBA1 gene and phylogenetic tree using Sanger sequencing and next generation sequencing A.PCR results of NBA1 gene; B.Phylogenetic tree using Sanger sequencing and next sequencing techonoly(Underlined is sequences of Sanger sequencing); C.Bacteria liquid PCR results of NBA1 gene
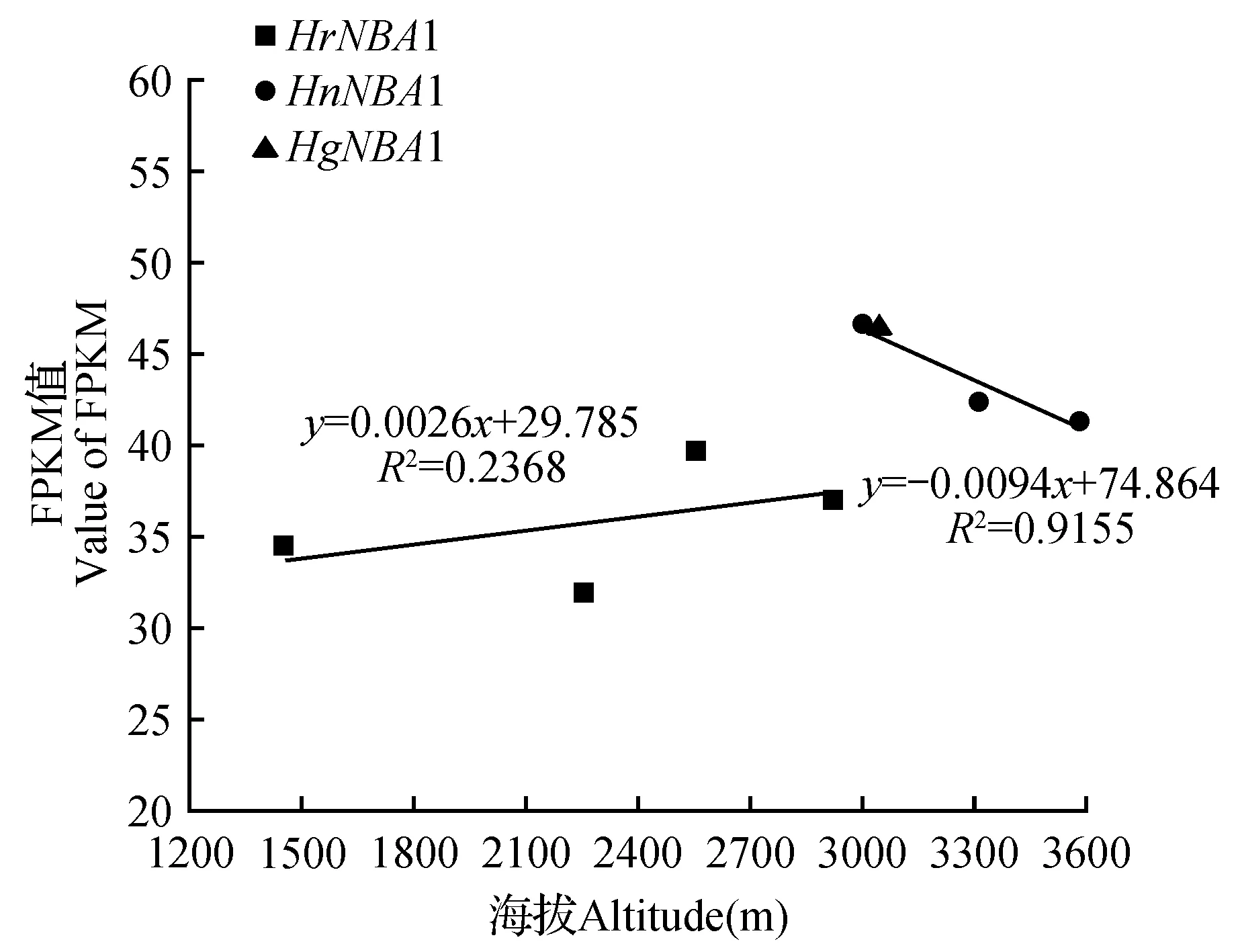
图2 NBA1基因在3种沙棘不同海拔下的FPKM值Fig.2 FPKM values of NBA1 gene in three species of Hippophae L. at different altitudes
2.2 NBA1基因在3种沙棘中的表达量分析
基因的表达水平可用转录本的丰度来体现。转录本丰度越高,即代表基因表达水平越高。在转录组数据分析中,可通过转录本的reads/fragments数来计算基因的表达水平[47]。本研究统计并分析了该基因在中国沙棘、肋果沙棘、棱果沙棘中的reads数量,并将测序深度与测序长度标准化,以FPKM(Fragments per Kilobase Million)值来表示基因的表达量水平。NBA1基因在中国沙棘(HrNBA1)、肋果沙棘(HnNBA1)、棱果沙棘中(HgNBA1)的FPKM均值分别为35.772、43.542、46.805。HrNBA1的FPKM均值稍低于HnNBA1与HgNBA1。以海拔为横坐标,FPKM值为纵坐标对HrNBA1与HnNBA1作线性回归分析,结果显示,HrNBA1的FPKM值随着海拔的升高而增加,R2值为0.237。HnNBA1的FPKM值随海拔的升高而减小,R2值为0.916。
2.3 NBA1基因在亲本种中国沙棘与肋果沙棘中的序列差异分析
利用NCBI网站的BLASTX程序查找NBA1基因的同源蛋白质序列,结果发现该序列与Tremaorientale的BRCA1-A&BRISC复合体序列相似性最高,E值为5e-145,序列覆盖度为72%。利用DNASTAR软件中的Seqbuilder与Editseq程序获得HrNBA1及HnNBA1的CDS区。全长均为771 bp,共编码256个氨基酸。二者在618,652及706的碱基位点的突变导致其编码的氨基酸序列在218位点及236位点有差异。如图3所示,将HrNBA1及HnNBA1与人类中NBA1氨基酸序列比对得知,HrNBA1及HnNBA1的VWA结构域位于135~218区域。HrNBA1在位于VWA结构域的218位点处为甲硫氨酸(M),HnNBA1为亮氨酸(L);236位点处HrNBA1为丝氨酸(S),HnNBA1为脯氨酸(P)。这两处氨基酸位点的变化可能导致HrNBA1及HnNBA1的高级结构产生差异。
2.4 NBA1基因在亲本种中国沙棘与肋果沙棘中的蛋白质三级预测结构及差异分析
ProtParam工具分析得出,HrNBA1与HnNBA1的分子式分别为:C1257H1998N352O389S14,C1260H2002N352O388S13;分子量分别为:28 714.66,28 706.66;等电点均为5.910;且为亲水性蛋白。分析其在细胞中的定位得知,该蛋白质位于细胞核内,且核定位序列为209~240区域。Zhanglab网站I-TASSER程序预测HrNBA1与HnNBA1蛋白的三级结构如图4所示,二者的蛋白质三级结构与小鼠中BRCA1-A复合体亚基NBA1的蛋白质三级结构高度相似[48]。也与TFIIH转录因子的Tfb4/p44亚基的蛋白质三级结构相似[49]。该蛋白质的结合位点由26,59,63,74,141,165,167,197,198,200,211等十一个位点的氨基酸组成;HrNBA1与HnNBA1在218位点处氨基酸的差异导致二者218位点后的蛋白质结构发生变化;236位点处HnNBA1的脯氨酸导致此处蛋白质的无规则卷曲发生折叠,HrNBA1较之更为舒展。
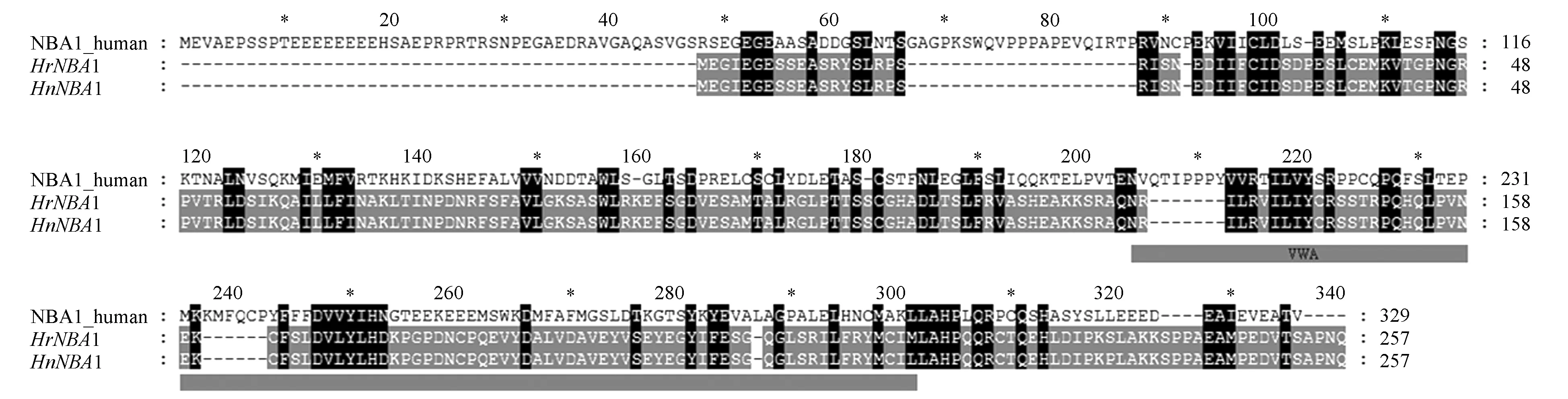
图3 HrNBA1及HnNBA1与人类的NBA1氨基酸序列Fig.3 Amino acid sequence of NBA1 in Human and HrNBA1 and HnNBA1

图4 NBA1的蛋白质三级预测结构 A,C.HrNBA1的三级预测结构(cartoon模式);B,D.HnNBA1的三级预测结构(mesh模式,红色标记为该蛋白的配体结合位点)Fig.4 Predicted results of the tertiary structure of protein of NBA1 A,C.The tertiary prediction structure of HrNBA1(in which A,B is the cartoon mode); B,D.The tertiary prediction structure of HnNBA1(C,D is the mesh mode,red is labeled as the ligand binding site of the protein)
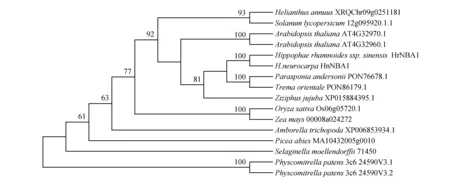
图5 14个物种中NBA1蛋白的系统进化树 该系统发育树中支持率小于50的均未显示Fig.5 Phylogenetic tree for NBA1 protein in 14 species The support rate less than 50 is not shown in this phylogenetic tree
2.5 NBA1基因在14个陆生植物中的进化关系分析
BRCA1-A在动物中为重要的抑癌基因[27~28],其亚基NBA1在复合体稳定中扮演着重要的角色[17]。NBA1基因在哺乳动物中有大量研究[19],植物中鲜少有人研究。本地BLAST结果显示,Chlamydomonasreinhardtii中无该基因的同源蛋白质。将其他12物种中共得到14条同源蛋白质序列,与本试验中的HrNBA1与HnNBA1做进化树分析。利用Prottest软件基于BIC模型计算得出,“JTT+I”为最佳氨基酸模型。利用最大似然法选择“JTT+I”模型建树。结果表明,该复合体基因在14个陆生植物中广泛存在,且序列较保守。如图4所示,该基因在这14个陆生植物中的进化关系符合APG version14的植物系统发育树关系[50]。
3 讨论
通过的RNA-seq数据计算直系同源基因对发现,NBA1基因在中国沙棘与肋果沙棘中为单拷贝基因,在棱果沙棘中以共显性的方式存在。
3.1 NBA1基因在3种沙棘中的适应性分化
青藏高原的沙棘属植物分布于不同的海拔,肋果沙棘较中国沙棘而言生长于更高的海拔,因此其长期暴露在更为寒冷、辐射更高的环境中。而生长于极端环境的植物已经进化出一套独特的适应机制[30],故推测中国沙棘与肋果沙棘已进化出一系列基因以应对高海拔引起的低温高辐射等极端环境。本研究通过比较中国沙棘NBA1基因与肋果沙棘NBA1基因的碱基序列发现,有3个碱基(618,652和706位点)发生突变,其中两处(652与706位点)为非同义突变,这两处碱基突变引起二者在218位点及236位点处氨基酸发生变化。Shao G等学者研究人类中的NBA1发现,该蛋白质由329个氨基酸组成,其中第95至第298位的氨基酸片段构成VWA结构域[24],该结构域可能与泛素化有关[29]。本研究中HrNBA1及HnNBA1由256个氨基酸组成,其中VWA结构域为第135位至第218位。HrNBA1及HnNBA1的218差异位点正好位于VWA结构域。将HrNBA1及HnNBA1氨基酸序列与13个位于植物系统进化树关键节点的物种的NBA1同源蛋白序列进行比对,其中在Chlamydomonasreinhardtii中未发现与NBA1的同源蛋白序列,在其他12个陆生植物中均找到了NBA1的同源蛋白。对包括中国沙棘与肋果沙棘在内的14种植物中NBA1的氨基酸序列进行比对发现,该蛋白质序列在这些物种中非常保守,这与Shao G等对人类等哺乳动物中NBA1蛋白序列的保守性研究结果一致[24]。进一步分析14种植物的NBA1氨基酸序列得知,位于VWA结构域的218位点在HrNBA1中为甲硫氨酸,在其他13个物种中均为保守的亮氨酸。这表明相比于中国沙棘而言,肋果沙棘的NBA1具有更加保守的VWA结构域,也具更加完备的NBA1蛋白功能,推测生长于更高海拔的肋果沙棘通过更加保守的蛋白质序列来保持NBA1基因的功能。BRISC复合体主要在细胞核外行使功能[21],BRCA1-A复合体位于细胞核中[29],该蛋白的亚细胞定位结果显示,HrNBA1与HnNBA1均位于细胞核内。但由于细胞核内也存在少量BRISC复合体,因此也不排除本研究中的NBA1蛋白是位于细胞核中少量BRISC复合体的NBA1亚基。目前尚不清楚核内的BRISC复合体行使何种功能,因此后续基于NBA1蛋白为BRCA1-A复合体亚基来分析本研究中的NBA1蛋白,BRCA1-A复合体中的NBA1亚基可与ABRAXAS-BRCC36二聚体及BRE结合,构成该复合体的蛋白支架[29],随后RAP80亚基结合至该支架之上形成完整的BRCA1-A复合体,装配完整的复合体结合至DNA双链断裂位点,从而使BRCA1远离DNA损伤位点,通过抑制切除来达到修复断裂的DNA双链的目的[23~24,29]。本研究中计算机蛋白质三维结构建模结果进一步显示,HrNBA1与HnNBA1的三级结构有较大差异,且二者的配体结合位点在空间排布上发生显著变化。推测218位点的差异使中国沙棘的NBA1与肋果沙棘的NBA1在蛋白质三级结构上发生了较大变化,导致了二者结合位点发生变化,从而使HrNBA1与HnNBA1结合其他亚基出现差异,进一步导致二者在修复高辐射引起的DNA损伤功能方面有差异。除NBA1基因序列的差异之外,本研究发现该基因在3种沙棘中的表达量也有所不同,其在3种沙棘中均大量表达。Shao G等学者敲低Hela细胞NBA1基因表达量后发现细胞对电离辐射更加敏感[18,24],Bolton等研究发现上皮卵巢癌细胞中NBA1基因的过表达,保护了BRCA1功能失调的细胞中修复双链DNA断裂的活性,从而使细胞对DNA损伤更加耐受[48]。推测中国沙棘通过升高NBA1基因的表达量来应对随着海拔升高而增强的高辐射引起的DNA损伤,而肋果沙棘则通过保持该基因序列的保守性来保证其在DNA损伤修复方面的功能,这些差异可能使得中国沙棘与肋果沙棘对不同生境的适应产生差异。
3.2 第二代测序技术在鉴定杂种方面的适用性
目前鉴定杂交物种通常采用形态鉴定、细胞学标记、分子标记、基因序列的系统分析等方法。上述方法各有优缺点,如由于形态特征的可塑性,常常模糊亲代与子代的界限。细胞学标记需要明确染色体倍性与亲本范围。分析基因序列是目前鉴定杂交物种最常用的手段;单拷贝核基因为理想的分析序列[31]。本研究的Sanger测序结果表明,NBA1基因的二代测序序列准确。分析二代测序序列与Sanger测序序列进化树得知,中国沙棘与肋果沙棘明显聚为两枝;在二代测序的棱果沙棘个体中,Gb,Gc个体与中国沙棘聚为一枝,Ga,Gd个体与肋果沙棘聚为一枝。以棱果沙棘Gd个体为例,进行Sanger法测序结果表明,Gd个体同时存在HrNBA1与HnNBA1。这是由于棱果沙棘为中国沙棘与肋果沙棘的双向同倍化杂交物种[2,7],故双亲纯合基因型中的等位基因以共显性的方式存在于棱果沙棘的杂合基因型中,这与二代测序不适用于杂交物种个体测序的传统认识是一致的。二代测序技术不适用于在个体水平鉴定杂交物种,这是因为二代测序原理与组装方式的统计学特点,会随机丢失测序个体体内的部分共显性基因[51],但若选取居群内多个杂种个体进行测序,则可在一定程度上随机反应杂种的亲本来源。
综上所述,本文研究了中国沙棘、肋果沙棘与棱果沙棘中受环境选择的功能基因NBA1在三者中的适应性分化及与三者不同海拔适应的关系。并以NBA1基因为例,探讨了二代测序的准确性以及其并不适用于在个体水平上鉴定杂种。以期为选择二代测序鉴定杂交物种提供借鉴;也为进一步揭示不同海拔下沙棘属植物的适应性进化提供基因数据基础。
猜你喜欢
今日农业(2022年3期)2022-11-16
农业工程学报(2022年13期)2022-10-09
今日农业(2022年13期)2022-09-15
麦类作物学报(2022年3期)2022-05-19
大气科学学报(2022年2期)2022-05-12
临床与实验病理学杂志(2022年3期)2022-04-06
聊城大学学报(自然科学版)(2022年3期)2022-02-14
作物学报(2022年3期)2022-01-22
食品安全导刊(2021年21期)2021-08-30
西北农业学报(2021年4期)2021-05-19
