
Sodium glucose co-transporter 2 inhibition reduces succinate levels in diabetic mice
2020-07-10 07:10LakshiniHeratNatalieWardAaronMagnoElizabethRakoczyMarcioKiuchiMarkusSchlaichVanceMatthews
World Journal of Gastroenterology 2020年23期
Lakshini Y Herat, Natalie C Ward, Aaron L Magno, Elizabeth P Rakoczy, Marcio G Kiuchi, Markus P Schlaich,Vance B Matthews
Abstract
Key words: Sodium glucose co-transporter inhibitors; Sodium glucose co-transporter 2;Diabetes; Diabetic retinopathy; Mouse; Gut microbiota; Empagliflozin; Dapagliflozin;Succinate; Akimba
INTRODUCTION
The gut microbiome plays an important role in physiology, the immune system development and digestion[1]. Microbial richness and the metabolic capacity of the microbiome is an indicator of health status. Most of the microbiota reside in the gut.This accounts for > 5 million genes and > 1000 species, which consists of a small number of phyla[1,2]. In addition, the large diversity and complexity of the microbiota can be divided into clusters of living organisms based on their ecosystem[1,3].
In recent years, the area of gut microbiome research has significantly increased in both human and murine studies. These studies highlight that changes in the gut microbiome and gut dysbiosis play a critical role in susceptibility to numerous diseases[4]. Although, it is widely accepted that diabetes and its complications are multifactorial, recent studies emphasise the importance of perturbations in the gut microbiota as a factor in the pathogenesis of diabetes[5-9]and its complications[10,11]. The mechanisms leading to an altered gut microbiota and gut dysbiosis during diabetes is associated with altered glycaemic control, obesity and insulin resistance, increased inflammation, oxidative stress and vascular permeability[12]. Interestingly, more studies are now focusing on the gut microbiome as a potential source of biomarkers of diabetes and its complications[13].
Sodium glucose co-transporter 2 inhibitors (SGLT2) are oral medications for the treatment of type 2 diabetes (T2D). This drug class blocks SGLT2 in the renal proximal tubules, thereby facilitating glucosuria and subsequently reduced plasma glucose levels and improved glycaemic parameters. The SGLT2 inhibitors canagliflozin,empagliflozin and dapagliflozin have been shown to decrease glycosylated haemoglobin levels, fasting glucose levels, body weight and blood pressure[14-17].Furthermore, SGLT2 inhibitors target two of the main problems of diabetes which are cardiovascular disease (CVD) and renal disease[15,17]. SGLT2 inhibitor related studies are currently underway in the field of type 1 diabetes (T1D)[18-20]. Given the limited understanding of SGLT2 inhibition and its beneficial effects on T1D, further preclinical studies are urgently needed. The objective of our investigation was to examine the effects of two independent SGLT2 inhibitors on gut health of a highly relevant mouse model of T1D and its complications.
MATERIALS AND METHODS
Animals
Specific pathogen free 10-wk-old male C57BL6/J, Kimba, Akita and Akimba mice[21-25]were obtained from the Animal Resources Centre (ARC, Perth, Australia). All experimental and animal handing activities were performed at the Harry Perkins Institute for Medical Research animal holding facility (Perth, Western Australia)according to the guidelines of the Institutional Animal Care and Use Committee.Animal ethics approval (AE141/2019) was received from the Harry Perkins Institute for Medical Research Animal Ethics Committee.
All mice were acclimatised for seven days. Mice were housed under a 12 h light/dark cycle, at 21 ± 2°C and were given a standard chow diet (Specialty Feeds,Glen Forrest, WA, Australia) with free access to food and drinking water containing the SGLT2 inhibitor (dapagliflozin or empagliflozin; Ark Pharma Scientific Limited,China; 25 mg/kg) or vehicle dimethylsulfoxide. Mice underwent treatment for a period of 8 wk. Drinking water containing the SGLT2 inhibitor or vehicle was freshly prepared and replaced on a weekly basis. The specific number of animals per treatment group is as follows: (1) C57BL6/J:n= 7 for vehicle;n= 5 for dapagliflozin;n= 4 for empagliflozin; (2) Kimba:n= 5-6 for vehicle;n= 6 for dapagliflozin;n= 7 for empagliflozin; (3) Akita:n= 7 for vehicle;n= 6 for dapagliflozin;n= 3 for empagliflozin; and (4) Akimba:n= 2-3 for vehicle;n= 3 for dapagliflozin;n= 3 for empagliflozin.
Tissue and serum sample collection
At the end of the experiment, mice were deeply anesthetised using isoflurane inhalation. Blood samples were collected using cardiac puncture and placed on ice immediately. Samples were centrifuged, serum was collected and stored at -80 °C for subsequent experiments. Mice were sacrificed, the cecum content and kidneys were collected, snap frozen and stored at -80°C for subsequent experiments. Pancreatic tissue was collected and fixed for 24 h in 10% buffered formalin for histology.
Succinate and short chain fatty acid analysis
The concentration of succinate and the short chain fatty acid (SCFA) butyric acid was measured in serum using gas chromatography-mass spectrometry as previously described[26,27]. Data were calculated in µmol/L units.
Pancreatic tissue histology and digital imaging
Upon fixation and processing in 70% ethanol overnight, pancreatic tissue was paraffin-embedded and sectioned at 5 µm thickness using a Leica semi-automated RM2245 microtome (Leica Biosystems, Sydney, Australia). Sections were stained with Gill’s hematoxylin (Sigma-Aldrich, Sydney, Australia) and alcoholic eosin (Sigma-Aldrich New South Wales, Australia) for the analysis of pancreatic histology. Tissue sections were visualised and imaged using the inverted Nikon Eclipse Ti microscopic system (Nikon, Tokyo, Japan) and a CoolSNAP HQ2 digital camera (Photometrics,Tucson, AZ, United States). Image analysis was conducted using NIS-Elements Advanced Research software (Nikon, Tokyo, Japan).
Insulin enzyme-linked immunosorbent assay
Serum insulin was determined using a rat/mouse Insulin enzyme-linked immunosorbent assay (ELISA) (Millipore, Australia; EZRMI-13K) according to the manufacturer instructions.
ELISA for leptin
Mouse serum was diluted 1 in 20 and leptin concentrations were determined by Mouse/rat leptin ELISA kit following manufacturer instructions (Quantikine ELISA;Mouse/rat leptin immunoassay; R&D Systems; Minneapolis, MN, United Sates;MOBOO).
ELISA for norepinephrine
Frozen mouse kidney tissue was homogenized on ice in phosphate buffered saline containing cOmplete™, ethylenediaminetetraacetic acid-free protease inhibitor cocktail (Merck, Victoria, Australia) at 10 µL per 1 mg of tissue. Debri free supernatant was analysed for norepinephrine content according to manufacturer instructions(Mouse Norepinephrine NA ELISA Kit; Cusabio, China; CSB-E07870m).
Statistical analysis
The number of mice required was determined with the assistance of Mrs Sally Burrows (Biostatistician, Royal Perth Hospital). Data was analysed using a two-tailed Student'sttest. Quantitative data is presented as mean ± SE. Data was deemed significant whenP< 0.05. Graphs were produced with GraphPad Prism 8 (GraphPad Software Inc., CA, United States).
RESULTS
Type 1 diabetes promotes hypoinsulinemia and pancreatitis in Akita and Akimba mice
In our studies, we used 4 strains of mice. We used C57BL/6J mice as our wild-type strain. The Kimba (VEGF+/+) mouse model possesses transient overexpression of human vascular endothelial growth factor (hVEGF) in photoreceptors and hence,displays changes associated with diabetic retinopathy (DR) but lacks the hyperglycaemic background. The Akita (Ins2Akita) mouse is a T1D model that possesses a dominant mutation in the Mody4 locus in theinsulin 2gene. Male heterozygous mice possess hyperglycaemia and features of DR. The insulin protein is incorrectly folded in Akita mice and this results in β-cell toxicity. By crossing Akita and Kimba mice, the Akimba (Ins2AkitaVEGF+/-) mouse model is generated where the interplay between high blood glucose levels and VEGF-induced pathology can be investigated in diabetic complications such as DR. Insulin destruction and a concomitant reduction in insulin production is a hallmark of T1D. As predicted, the Akita allele in the Akita and Akimba mice conferred hypoinsulinemia compared to C57BL/6J and Kimba mice (Figure 1). In agreement with the higher insulin levels,islets were frequently observed in pancreata from C57BL/6J (Figure 2A) and Kimba mice (Figure 2B). However, islets were observed less frequently in Akita (Figure 2C)and Akimba mice (Figure 2D). In addition, apoptotic bodies were observed in the islets of Akita mice (Figure 2C). The acinar cells of Akita (Figure 2C) and Akimba(Figure 2D) mice appeared swollen which is indicative of acute injury or pancreatitis.These cells possessed high levels of eosin staining.
SGLT2 inhibition influences the main products and intermediate metabolites of gut metabolism in Akita and Akimba mice
Succinate is an intermediate metabolite of gut metabolism and has been shown to be a pathogenic factor in DR[28-30]. We hypothesised that SGLT2 inhibition may reduce the levels of this pathogenic factor in Akimba mice which develop DR. We did indeed demonstrate that dapagliflozin significantly reduced succinate levels (Figure 3A) and empagliflozin suppressed succinate levels (Figure 3B).
The main products of gut metabolism are SCFAs. We examined levels of the beneficial SCFA butyric acid and observed that levels were significantly increased in Akita mice treated with dapagliflozin and mildly elevated with empagliflozin therapy(Figure 3C).
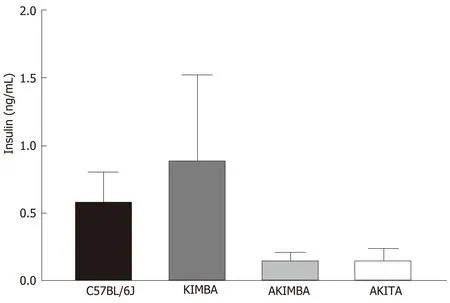
Figure 1 lnsulin dysregulation prevails in mice with type 1 diabetes. Serum insulin levels are reduced in Akimba and Akita mice. n = 3-8 mice/group. All data represented as mean ± SE.
SGLT2 inhibition with dapagliflozin and empagliflozin reduces activation of the sympathetic nervous system in type 1 diabetic mice
Our previously published findings have uncovered that one of the mechanisms by which SGLT2 inhibition imparts metabolic benefits is by reduced activation of the sympathetic nervous system (SNS)[31,32]. Levels of the major neurotransmitter of the sympathetic system, norepinephrine, were measured in the kidneys of our T1D mice.Consistent with our previously published work, SNS activation as evidenced by norepinephrine content, was significantly reduced with both dapagliflozin and empagliflozin therapy in Akita mice (Figure 4A). Norepinephrine content was also significantly reduced after empagliflozin treatment of Akimba mice (Figure 4B).
SGLT2 inhibition promotes hypoleptinemia in non-diabetic C57BL/6J and Kimba mice
As leptin is a metabolically beneficial adipokine, we examined the impact of SGLT2 inhibition on serum leptin levels. Previous studies have shown that SGLT2 inhibition may reduce serum leptin levels[33]. Interestingly, C57BL/6J mice also displayed significantly reduced serum leptin after empagliflozin treatment and reduced leptin after dapagliflozin therapy (Figure 5A). Kimba mice also possessed significantly reduced serum leptin after dapagliflozin treatment (Figure 5B) and suppressed leptin levels after empagliflozin therapy (Figure 5C). The inhibition of SGLT2 failed to influence leptin levels in Akita and Akimba mice.
DISCUSSION
Inhibition of SGLT2 in the kidney is a therapeutic approach to improve glucose homeostasis in diabetic patients. The clinical trial known as the EMPA-REG OUTCOME study has shown that the SGLT2 inhibitor, empagliflozin, increased glycaemic control and significantly reduced the progression of renal disease[15]. In addition, empagliflozin promoted a substantial 38% relative risk reduction in death from cardiovascular causes[17]. To date, the number of studies assessing the impact of SGLT2 inhibition on T1D and its complications including DR in preclinical and human cohorts is limited[34]. In our current study, we aimed to assess whether SCFAs and intermediate metabolites of gut metabolism may be influenced by SGLT2 inhibition in a mouse model of T1D and its complications. Furthermore, we attempted to uncover mechanisms by which SGLT2 inhibitors may impact the progression of disease in our mouse models.

Figure 2 Acute pancreatic injury in Akita and Akimba mice. A: Representative pancreatic images of C57BL/6J mice; B: Representative pancreatic images of Kimba mice; C: Representative pancreatic images of Akita mice; D: Representative pancreatic images of Akimba mice. Black arrows indicate islets; Yellow arrows and dotted outline indicates swollen acinar cells and orange arrows indicate apoptotic bodies in islets; Scale bar = 100 µm.
There is great interest in the role the gut microbiome plays in the metabolic syndrome and T2D[27]. Many chronic diseases including diabetes demonstrate altered bacterial composition[35], which may influence glucose metabolism and development of diabetic complications such as retinopathy[10,36]. SCFAs are a product of the gut microbiota. These products include acetate and butyric acid and contribute to immune and inflammatory responses, as well as control of lipid and glucose homeostasis. Studies from our group and others indicate that numerous drug classes alter the gut microbiome[26,37]. Additionally, a reduction in SCFA production is characteristic of metabolic dysfunction. In our previous studies, we analysed caecal SCFAs from dapagliflozin and vehicle treated hypertensive mice and demonstrated that SGLT2 inhibition results in an increase in the levels of acetate and butyric acid in hypertensive Schlager mice[31]. In a chronic kidney disease mouse model, Mishimaet al[38]also demonstrated that the SGLT2 inhibitor canangliflozin significantly increased the cecal SCFA acetate, propionate and butyrate. Our previous data highlighted a potentially advantageous impact of SGLT2 inhibition on the microbiome.Additionally, as our results were produced only after 2 wk of therapy, future studies should allow a more thorough examination of the status of the microbiome to determine whether a longer duration of therapy confers further benefits on the microbiome. In our current study, our T1D and DR mice were treated with SGLT2 inhibitors for 8 wk. We found that SGLT2 inhibition promoted beneficial changes in the metabolite succinate (Figure 3A and 3B) and the SCFA butyrate (Figure 3C).Recent findings have uncovered that dapagliflozin treatment of T2D Db/Db mice promotes increases in the beneficial bacterial speciesAkkermansia muciniphilaand reduces the Firmicutes : Bacteroidetes (F : B) ratio which is associated with a lean phenotype[39]. Our future studies will plan to assess the levels ofAkkermansia muciniphilaand the F:B ratio in our T1D and DR mouse models before and after SGLT2 inhibition.
Succinate is an intermediate metabolite of gut metabolism that has been correlated with a worsened DR phenotype. It signals through the succinate specific receptor known as GPR91 and it has been shown that succinate may promote the angiogenic factor VEGF in retinal ganglion cells and this may promote the pathogenesis of DR[40].Matsumoto and colleagues[29]demonstrated that vitreous levels of succinate were elevated in human subjects with active proliferative DR. In our novel Akimba mouse model which reproducibly presents with DR[21], SGLT2 inhibition with both dapagliflozin (Figure 3A) and empagliflozin (Figure 3B) lowered serum succinate levels. It would be interesting to measure vitreous succinate and VEGF levels in our mice in future studies. In addition, it would be mechanistically insightful to assess GPR91 expression in the eyes of all mouse strains before and after SGLT2 inhibition as GPR91 and SGLT2 (unpublished data) are both expressed in retinal ganglion cells.
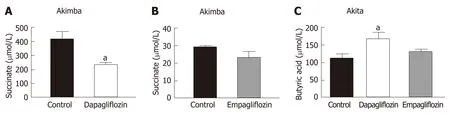
Figure 3 The diabetic retinopathy pathogenic metabolite, succinate, is downregulated with sodium glucose co-transporter 2 inhibition in Akimba mice. A:Serum succinate levels in Akimba mice treated with dapapagliflozin; B: Serum succinate levels in Akimba mice treated with empagliflozin; C: Serum levels of the beneficial short chain fatty acid, butyric acid, are elevated in diabetic Akita mice with sodium glucose co-transporter 2 inhibition. n = 3-7 mice/group for all groups except for control Akimba (Figure 3B) where n = 2; aP < 0.03; Data represented as mean ± SE.
Hyperactivation of the SNS is characteristic of many metabolic diseases such as obesity and T2D[41,42]. Our team previously studied the impact of SGLT2 inhibition on SNS activation[31,32]. We were the first to demonstrate in human proximal tubular cells and mouse models of obesity and neurogenic hypertension that: (1) Activation of the SNSviaincreased secretion of norepinephrine up-regulates SGLT2 protein levels and promotes SGLT2 translocation to the cell membrane; (2) Treatment of mice with a high fat diet and the SGLT2 inhibitor dapagliflozin promotes glucosuria, decreased weight gain and facilitated better glucose control; and (3) SGLT2 inhibition reduced SNS innervation in the kidney and the heart. Our data suggests that SGLT2 expression is upregulated by the major neurotransmitter of the SNS and that SGLT2 inhibition promotes sympatho-inhibition. Excitingly, we also now show that SGLT2 inhibition is once again sympathoinhibitory in our T1D and DR mouse models (Figure 4).
It appears that dapagliflozin was more bioactive than empagliflozin when it comes to the effects on succinate levels (Figure 3A and B). It has been shown in mouse models that dapagliflozin is superior to empagliflozin as it has an increased half-life,longer duration of action, higher distribution and long retention period in the kidney[43]. Therefore, this could be the likely reasoning for the dapagliflozin being more bioactive in relation to succinate levels.
Interestingly, empagliflozin was able to reduce norepinephrine levels to a greater degree compared to dapagliflozin in diabetic Akita mice (Figure 4A). This is interesting in the context of the EMPA-REG study[17]. In this study, factors driven by an increased SNS such as mortality from CVD were reduced to a markedly greater degree compared to the dapagliflozin DECLARE-TIMI 58 clinical trial[44]. Hence,regulation of the SNS may be one factor where empagliflozin may be specifically more effective than dapagliflozin.
The activation status of the SNS in T1D is currently quite controversial. It has been shown that there was a reduction in islet selective sympathetic nerves in human T1D pancreata isolated during autopsy compared to non-diabetic controls[45]. On the other hand, it has been demonstrated that children with T1D have a higher incidence of postural orthostatic tachycardia syndrome which is associated with sympathetic overactivity[46,47]. Our study conclusively shows that SGLT2 inhibition reduced levels of norepinephrine in our T1D and DR mouse models.
We assessed serum levels of the adipokine leptin in our mice treated with SGLT2 inhibitors or not. It was clear that leptin levels were reduced with SGLT2 inhibition with dapagliflozin and empagliflozin, particularly in C57BL/6J and Kimba mice(Figure 5A-C). This appears to be a common phenomenon that has been observed in response to a range of SGLT2 inhibitors which include ipragliflozin and canagliflozin[33,48]. We can conclude that the reductions in leptin were not due to reductions in body weight after SGLT2 inhibition in C57BL/6J and Kimba mice (data not shown). Reductions in leptin may also be metabolically beneficial as leptin is known to activate the SNS and may impair pancreatic β-cell function[49,50]. In future studies, we will also assess the effect of SGLT2 inhibition on other adipokines such as TNF-α and IL-6.
We are the first study to compare and contrast the effects of SGLT2 inhibition on the main products and intermediate metabolites of gut metabolism in Akita and Akimba mice. Excitingly, our results highlighted that after SGLT2 therapy, the pathogenic biomarker succinate is reduced in our Akimba DR mouse model. Our next task is to ascertain whether our finding may be reproduced in a human DR cohort treated with the SGLT2 inhibitor empagliflozin.
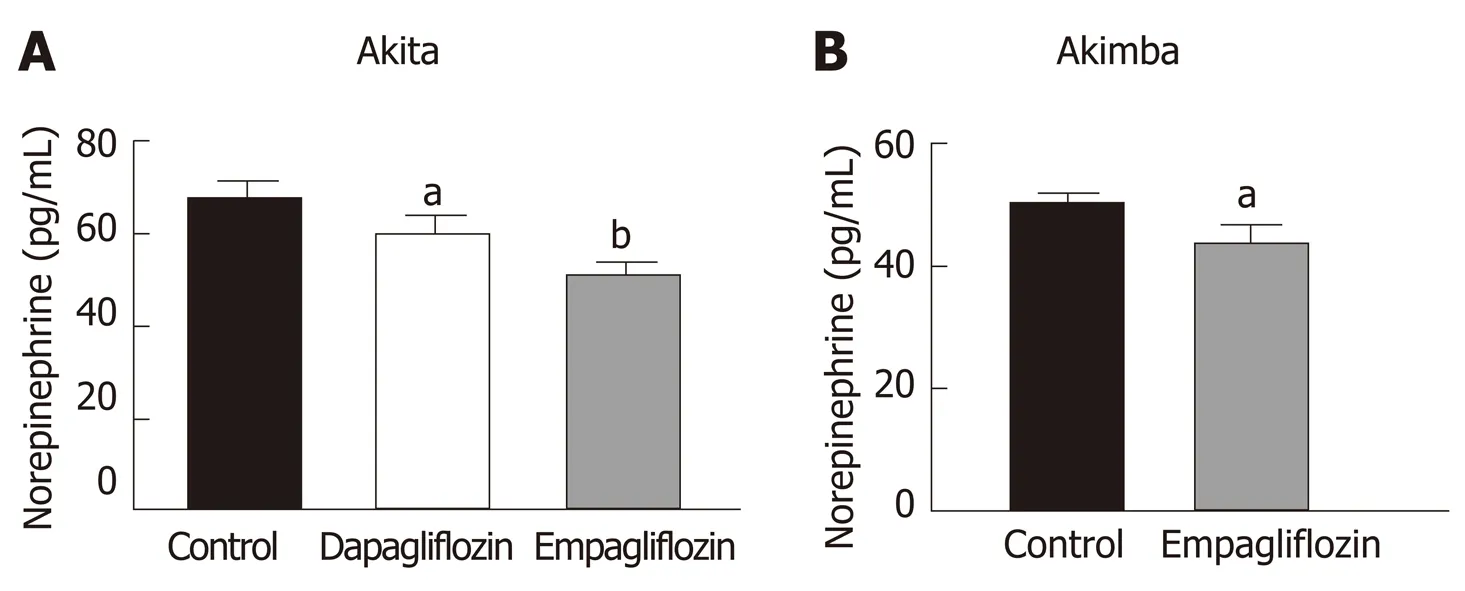
Figure 4 lnhibition of sodium glucose cotransporter 2 promotes sympathoinhibition in the kidney of diabetic Akita and Akimba mice. A: Renalnorepinephrine levels in Akita; B: Renal norepinephrine levels in Akimba mice. n = 3-7 mice/group; aP < 0.05; bP < 0.005; All data represented as mean ± SE.

Figure 5 lnhibition of sodium glucose cotransporter 2 reduces serum leptin levels. A: Serum leptin levels in C57BL/6J. B: Serum leptin levels in Kimba mice treated with dapagliflozin. C: Serum leptin levels in Kimba mice treated with empagliflozin. n = 4-7 mice/group; aP < 0.05; All data represented as mean ± SE.
ARTICLE HIGHLIGHTS

In comparison to C57BL/6J and Kimba mice, both Akita and Akimba mice showed reduced levels of insulin production due to the presence of the Akita allele. In line with this, Akita mice also showed the presence of apoptotic bodies within the pancreatic islets and the acinar cells of both the Akita and Akimba mice displayed swelling which is suggestive of acute injury or pancreatitis. In Akimba mice, SGLT2 inhibition with dapagliflozin for 8 wk significantly reduced succinate levels when compared to vehicle treated mice. Furthermore, succinate levels in Akimba mice treated with the SGLT2 inhibitor empagliflozin showed a similar trend. In diabetic Akita mice, the beneficial SCFA butyric acid was significantly increased after dapagliflozin treatment when compared to vehicle. There was a significant reduction in the kidney norepinephrine content in both dapagliflozin and empagliflozin treated Akita mice.Furthermore, the diabetic Akimba mice also showed a significant reduction in kidney norepinephrine content when treated with empagliflozin. Lastly, both non-diabetic C57BL/6J and Kimba mice showed significantly reduced serum leptin levels after dapagliflozin therapy.
Research conclusions
Our novel study compares and contrasts the effects of SGLT2 inhibition on the main products and intermediate metabolites of gut metabolism particularly in Akita and Akimba mice. We conducted studies using two independent SGLT2 inhibitors and showed that both inhibitors reduced the pathogenic biomarker succinate in our novel T1D Akimba mouse model of retinopathy. However, in relation to succinate levels in Akimba mice, dapagliflozin was more bioactive than empagliflozin, potentially due to factors such as increased half-life, longer duration of action, higher distribution and long retention period in the kidney. Furthermore, we demonstrate for the first time that SGLT2 inhibition is sympathoinhibitory in a T1D mouse model.
Research perspectives
In line with our findings, it would be mechanistically insightful in the future to assess the expression of the succinate specific receptor GPR91 in ocular tissue before and after SGLT2 inhibition as SGLT2 is expressed in the eye. Furthermore, it is important to determine whether our findings can be reproduced in patients with T1D and its complications who are treated with SGLT2 inhibitors.
ACKNOWLEDGEMENTS
The authors acknowledge Wei Ern Ong (School of Biomedical Sciences, University of Western Australia, Perth, Western Australia).
 World Journal of Gastroenterology2020年23期
World Journal of Gastroenterology2020年23期
- World Journal of Gastroenterology的其它文章
- Liver-directed therapies for liver metastases from neuroendocrine neoplasms: Can laser ablation play any role?
- Potential of the ellagic acid-derived gut microbiota metabolite - Urolithin A in gastrointestinal protection
- Endosonographic diagnosis of advanced neoplasia in intraductal papillary mucinous neoplasms
- Medications in type-2 diabetics and their association with liver fibrosis
- Pancreatic necrosis and severity are independent risk factors for pancreatic endocrine insufficiency after acute pancreatitis: A long-term follow-up study
- Impact of a national basic skills in colonoscopy course on trainee performance: An interrupted time series analysis